
Comparison of Canopy–Vegetation Parameters from Interior Parts to Edge of Multi-Story Grove Forest Patch and Meadow Field Within Rural Landscape for Soil Temperature and Moisture



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methodology
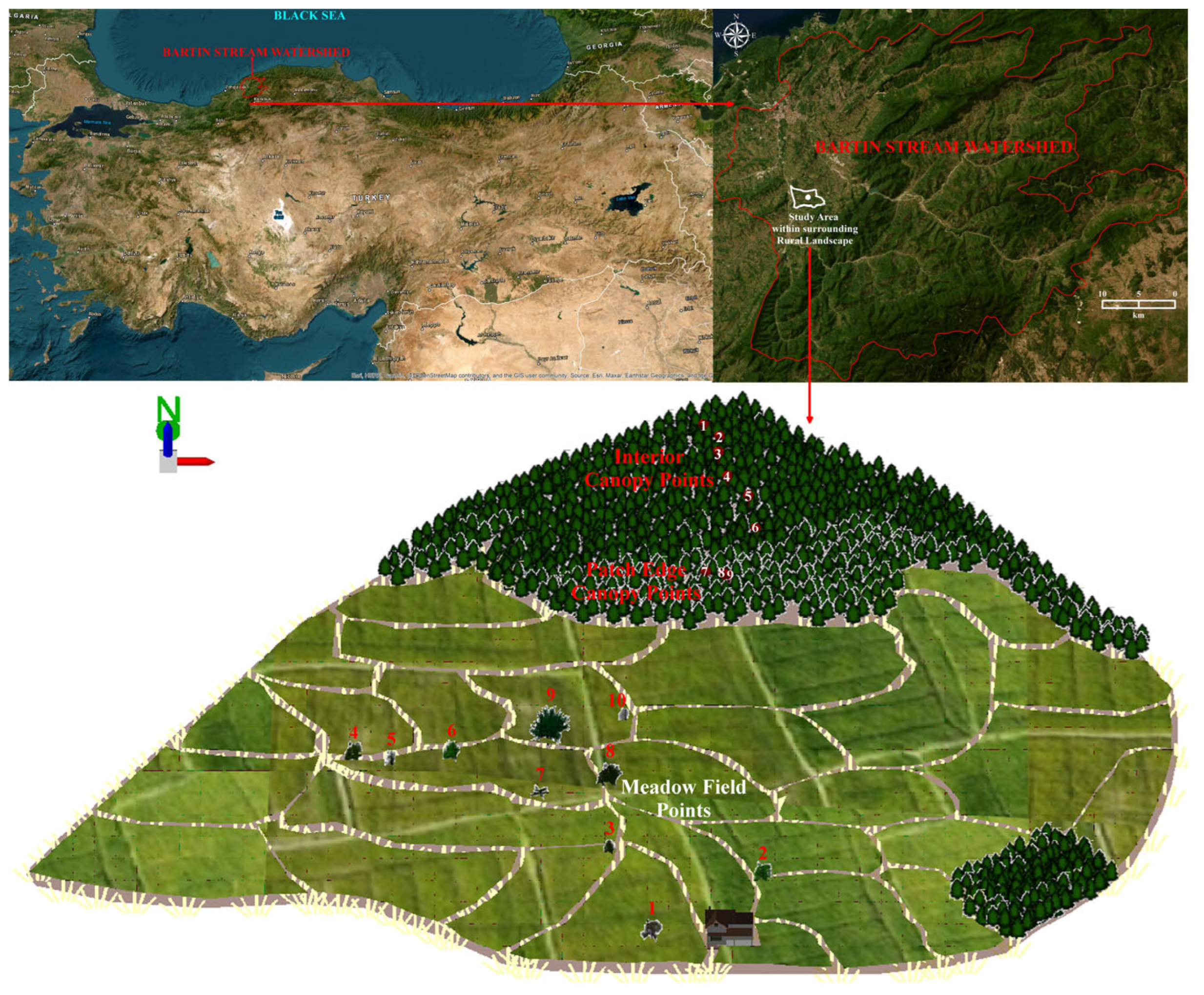
2.1. Location, Topography, Parent Material, Soil, and Climate
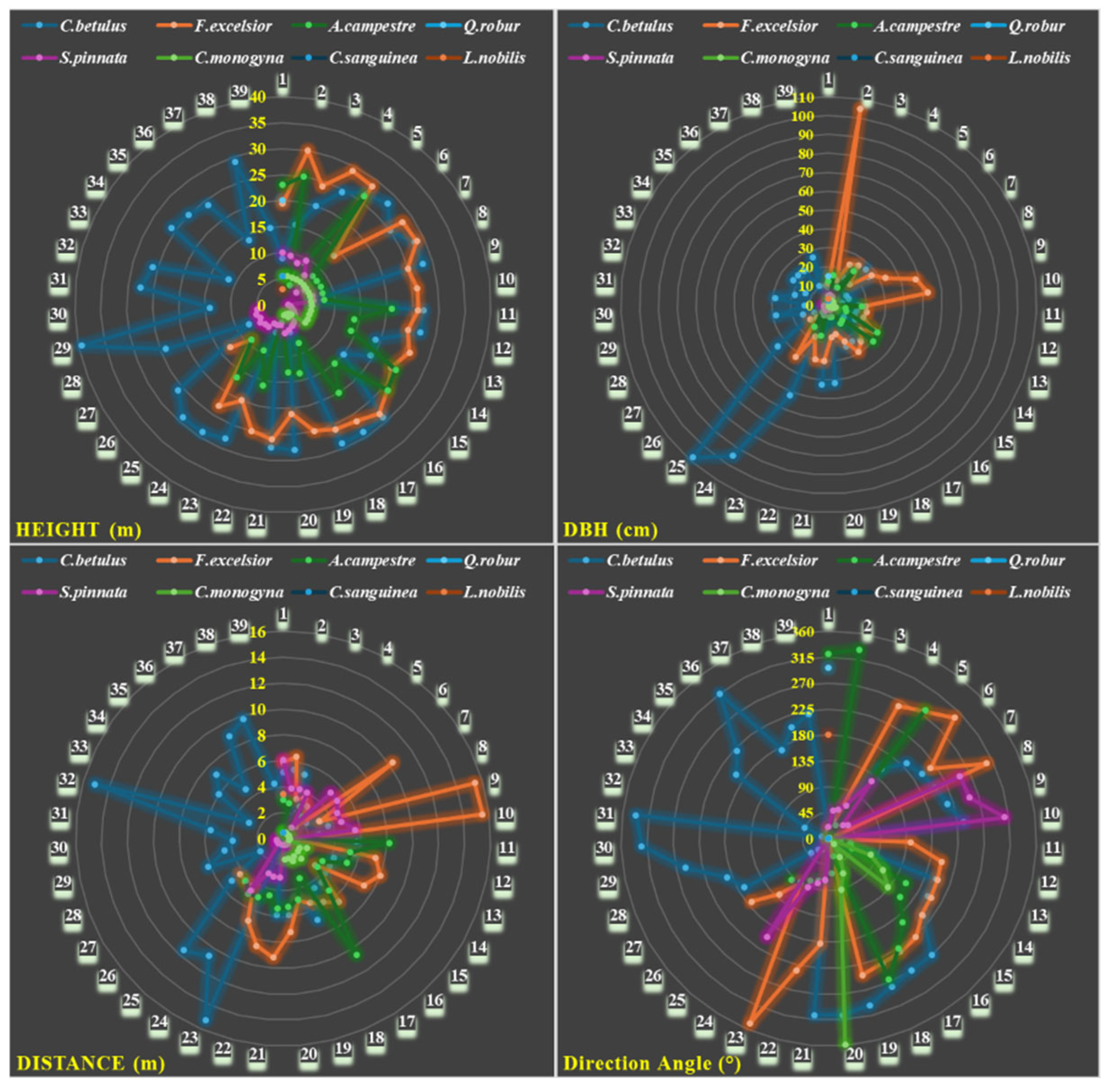
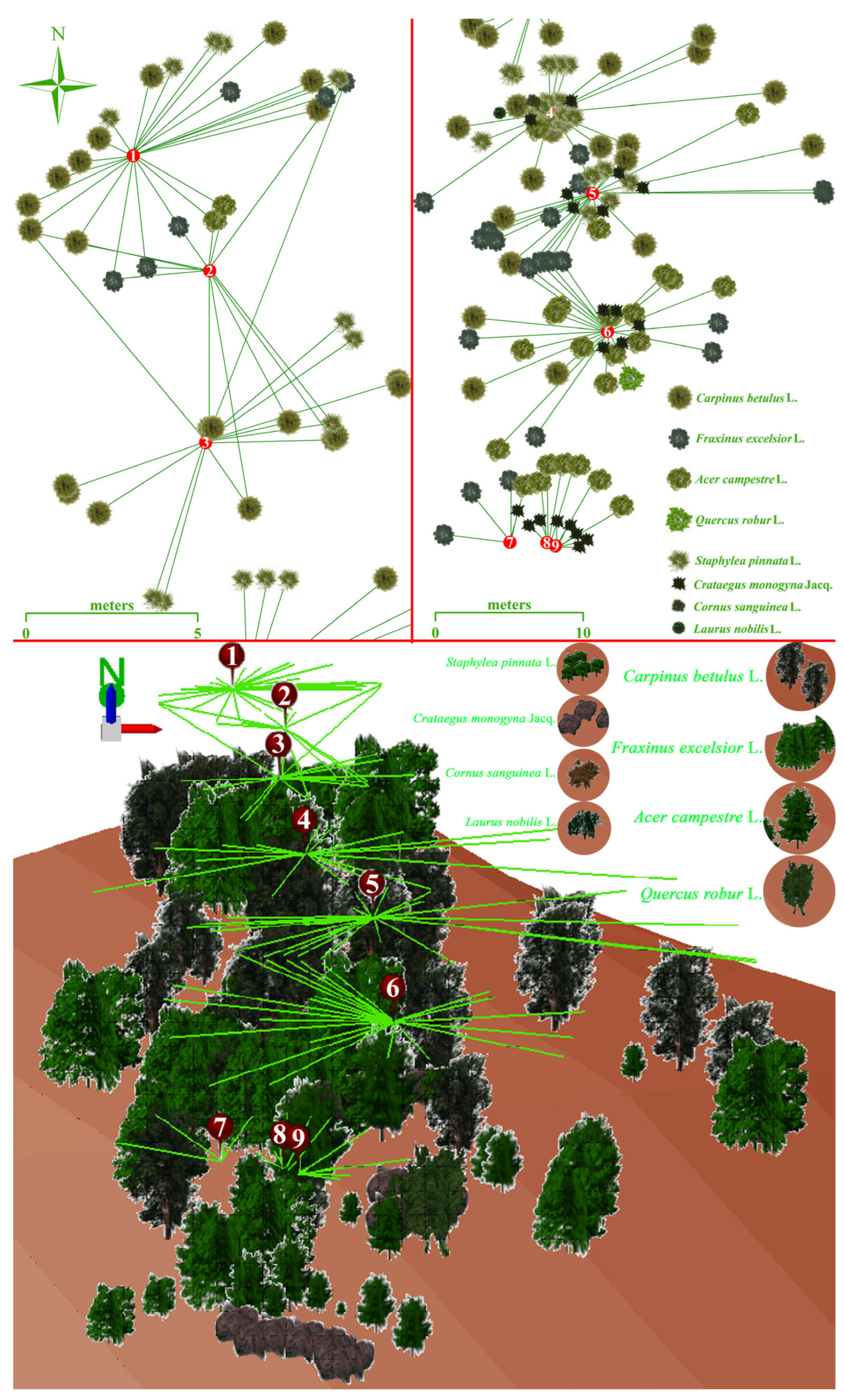
2.2. Forest Patch and Meadow Field
2.3. LAI, LT, CO, GF, Physiology, and Soil Temperature and Moisture Analyses
3. Results
3.1. Correlation of Referred and Measured Meteorological Data
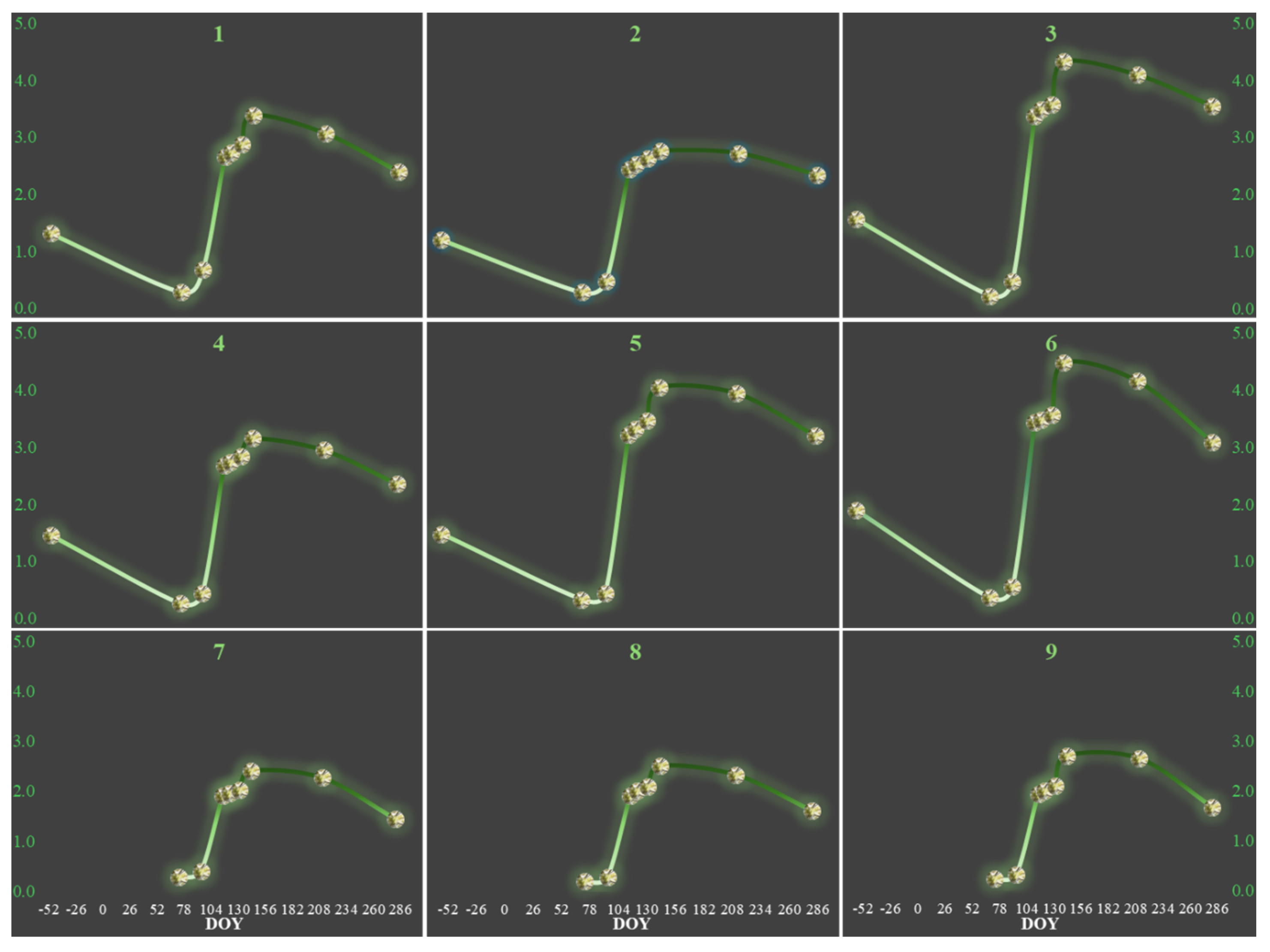
3.2. Leaf Senescence Periods (2023 and 2024)
3.3. Leaf Budburst and Onset
3.4. Leaf Unfolding and Stable Full-Leaf Periods
4. Discussion
4.1. Leaf Senescence Period
4.2. Leafless, Budburst, and Leaf Onset Period
4.3. Leaf Unfolding Period and Meadow Field
4.4. Full-Leaf Stable Period
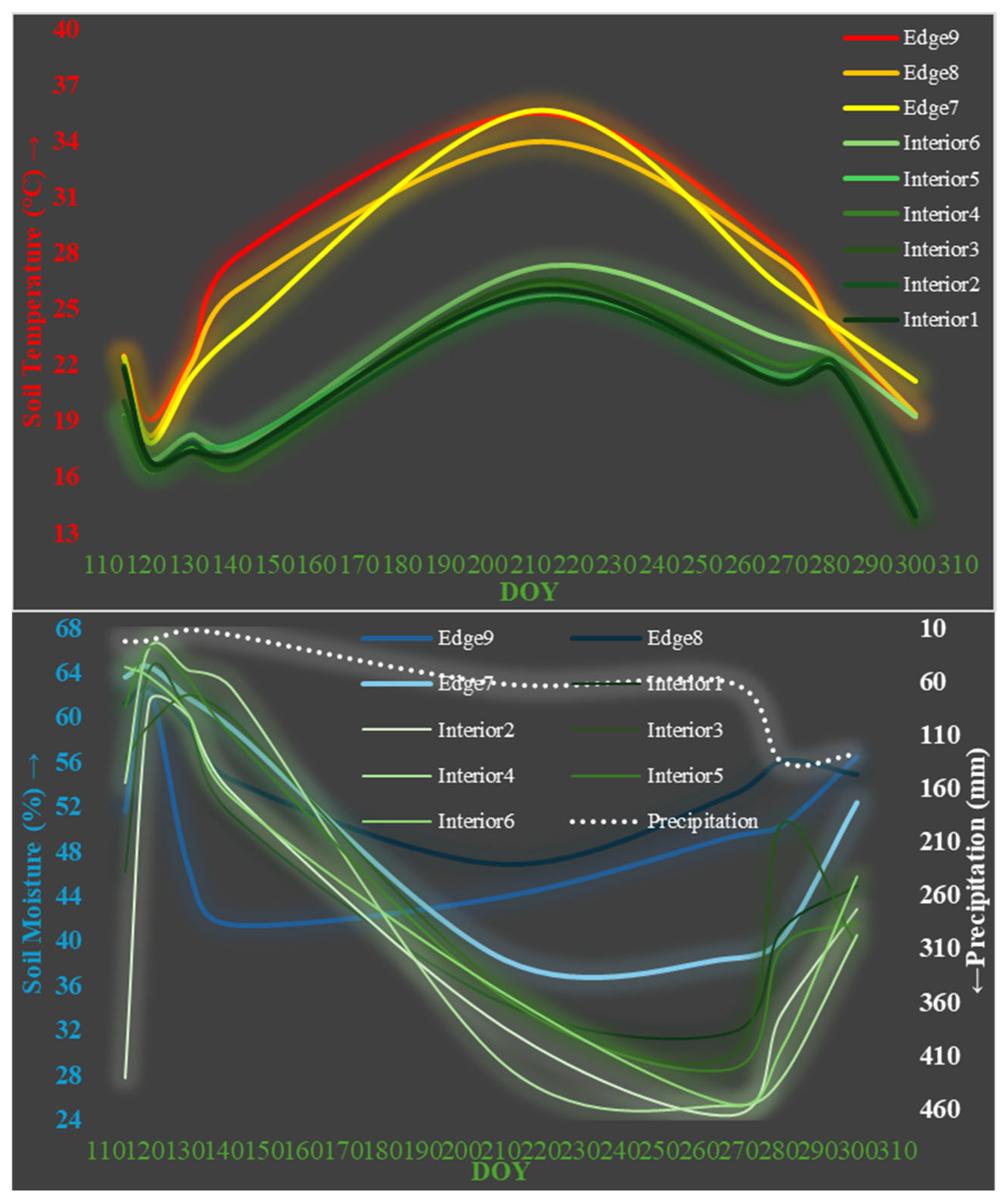
4.5. LAI, LT, GF, CO and Soil Temperature
4.6. LAI, LT, GF, CO and Soil Moisture
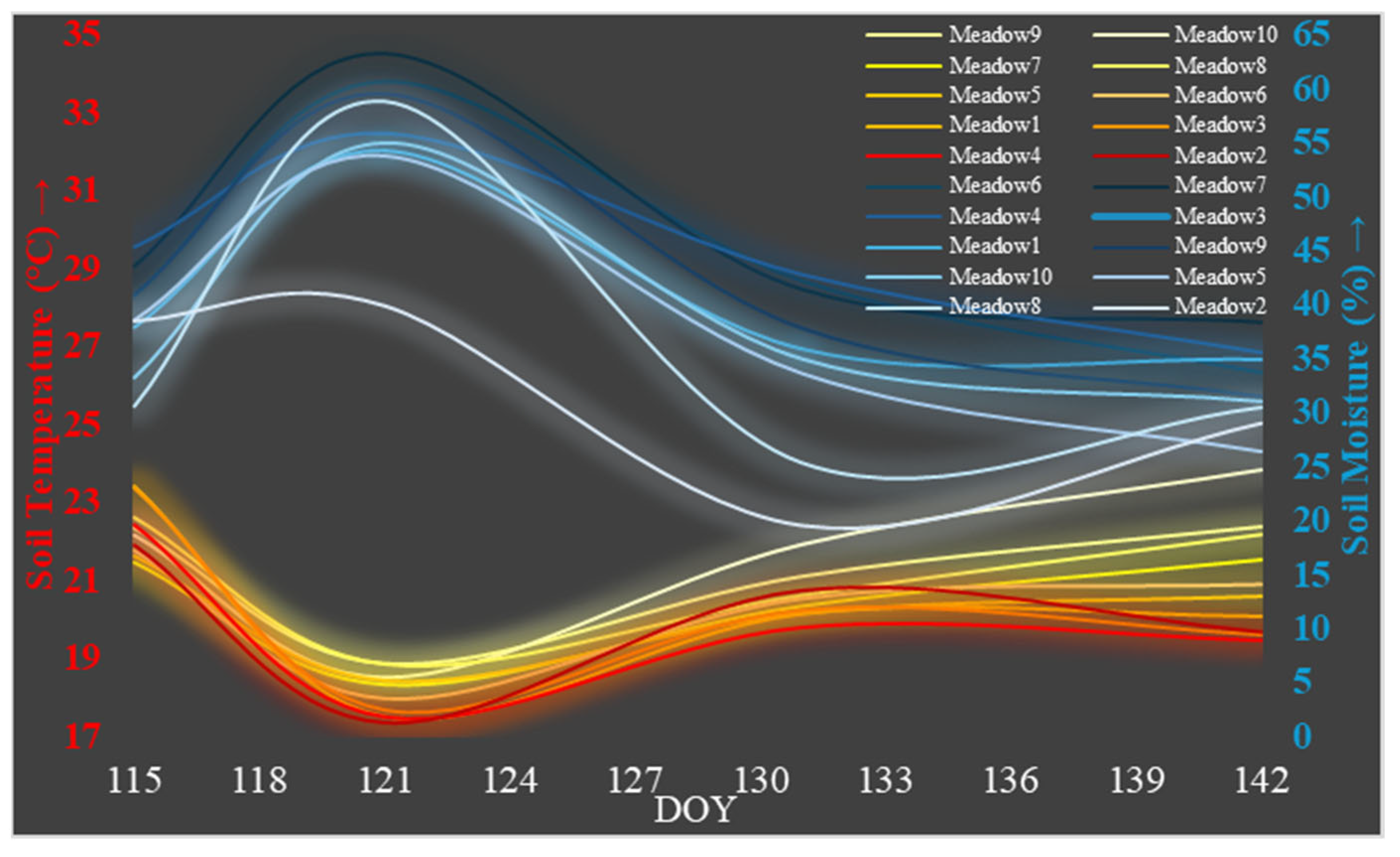
4.7. Meadow Field, Soil Temperature, and Soil Moisture
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buck, J.R.; St. Clair, S.B. Stand composition, proximity to overstory trees and gradients of soil moisture influence patterns of subalpine fir seedling emergence and survival. Plant Soil 2014, 381, 61–70. [Google Scholar] [CrossRef]
- Brockett, B.F.T.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Xu, M.; Zhong, Z.; Sun, Z.; Han, X.; Ren, C.; Yang, G. Soil available phosphorus and moisture drive nutrient resorption patterns in plantations on the Loess Plateau. For. Ecol. Manag. 2020, 461, 117910. [Google Scholar] [CrossRef]
- Olesinski, J.; Lavigne, M.B.; Krasowski, M.J. Effects of soil moisture manipulations on fine root dynamics in a mature balsam fir (Abies balsamea L. Mill.) forest. Tree Physiol. 2011, 31, 339–348. [Google Scholar] [CrossRef]
- Petraglia, A.; Cacciatori, C.; Chelli, S.; Fenu, G.; Calderisi, G.; Gargano, D.; Abeli, T.; Orsenigo, S.; Carbognani, M. Litter decomposition: Effects of temperature driven by soil moisture and vegetation type. Plant Soil 2019, 435, 187–200. [Google Scholar] [CrossRef]
- Petrie, M.D.; Hubbard, R.M.; Bradford, J.B.; Kolb, T.E.; Noel, A.; Schlaepfer, D.R.; Bowen, M.A.; Fuller, L.R.; Moser, W.K. Widespread regeneration failure in ponderosa pine forests of the southwestern United States. For. Ecol. Manag. 2023, 545, 121208. [Google Scholar] [CrossRef]
- Bonan, G. Ecological Climatology: Concepts and Applications, 3rd ed.; Cambridge University Press: New York, NY, USA, 2016; 692p. [Google Scholar]
- Chang, M. Forest Hydrology: An Introduction to Water and Forests, 2nd ed.; CRC Press, Taylor and Francis Group: New York, NY, USA, 2006. [Google Scholar]
- Waring, R.H.; Running, S.W. Forest Ecosystems: Analysis at Multiple Scales, 3rd ed.; Elsevier Academic Press: London, UK, 2007. [Google Scholar]
- Perry, D.A.; Oren, R.; Hart, S.C. Forest Ecosystems, 2nd ed.; The Johns Hopkins University Press: Baltimore, MD, USA, 2008; 632p. [Google Scholar]
- Stickley, S.F.; Fraterrigo, J.M. Understory vegetation contributes to microclimatic buffering of near-surface temperatures in temperate deciduous forests. Landsc. Ecol. 2021, 36, 1197–1213. [Google Scholar] [CrossRef]
- Forman, R.T.T.; Godron, M. Landscape Ecology; John Wiley & Sons Inc.: Hoboken, NJ, USA, 1986; 648p. [Google Scholar]
- Turner, M.G.; Gardner, R.H. Landscape Ecology in Theory and Practice; Pattern and Process, 2nd ed.; Springer Science + Business Media LLC: New York, NY, USA, 2015; 482p. [Google Scholar]
- Bae, J.; Ryu, Y. The magnitude and causes of edge effects on soil organic carbon stocks within and across urban to rural forest patches. Landsc. Urban Plan. 2021, 215, 104223. [Google Scholar] [CrossRef]
- Zellweger, F.; Cooms, D.; Lenoir, J.; Depauw, L.; Maes, S.L.; Wulf, M.; Kirby, K.J.; Brunet, J.; Kopecký, M.; Máliš, F.; et al. Seasonal drivers of understorey temperature buffering in temperate deciduous forests across Europe. Glob. Ecol. Biogeogr. 2019, 28, 1774–1786. [Google Scholar] [CrossRef]
- Parker, G.G. Tamm review: Leaf Area Index (LAI) is both a determinant and a consequence of important processes in vegetation canopies. For. Ecol. Manag. 2020, 477, 118496. [Google Scholar] [CrossRef]
- Leblanc, S.G.; Fournier, R.A. Measurement of forest structure with hemispherical photography. In Hemispherical Photography in Forest Science: Theory, Methods, Applications; Fournier, R.A., Hall, R.J., Eds.; Springer Science+Business Media B.V.: Dordrecht, The Netherlands, 2017; pp. 53–83. [Google Scholar] [CrossRef]
- Thimonier, A.; Sedivy, I.; Schleppi, P. Estimating leaf area index in different types of mature forest stands in Switzerland: A comparison of methods. Eur. J. For. Res. 2010, 129, 543–562. [Google Scholar] [CrossRef]
- Brüllhardt, M.; Rotach, P.; Schleppi, P.; Bugmann, H. Vertical light transmission profiles in structured mixed deciduous forest canopies assessed by UAV-based hemispherical photography and photogrammetric vegetation height models. Agric. For. Meteorol. 2020, 281, 107843. [Google Scholar] [CrossRef]
- TGDF (Turkish General Directorate of Forestry). Forest Management Plans (2011) of Bartın-Kozcağız Sub-District of Bartın Forestry Administration Affiliated to the Zonguldak Regional Forest Directorate; TGDF: Ankara, Turkey, 2011. [Google Scholar]
- TMAF (Turkish Ministry of Agriculture and Forestry). Digital Soil Maps of Bartın Stream Watershed; TGDF: Ankara, Turkey, 2005. [Google Scholar]
- TGDMRE (Turkish General Directorate of Mineral Research and Exploration). Digital Geological Maps of Zonguldak E-29 Section; TGDMRE: Ankara, Turkey, 2007. [Google Scholar]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils, 15th ed.; Pearson Education: Upper Saddle River, NJ, USA, 2016; 1104p. [Google Scholar]
- TSMS (Turkish State Meteorological Service). Meteorological Data of Bartın (17020) and Kozcağız (19007) (Turkey) Meteorology Stations; TSMS: Ankara, Turkey, 2024. [Google Scholar]
- Atalay, İ. Climate Atlas of Turkey; İnkılâp Bookstore Press: İstanbul, Turkey, 2011; 136p. (In Turkish) [Google Scholar]
- Liu, Y.; Liu, R.; Pisek, J.; Chen, J.M. Separating overstory and understory leaf area indices for global needleleaf and deciduous broadleaf forests by fusion of MODIS and MISR data. Biogeosciences 2017, 14, 1093–1110. [Google Scholar] [CrossRef]
- Wen, Y.; Zhuang, L.; Wang, H.; Hu, T.; Fan, W. An automated hemispherical scanner for monitoring the Leaf Area Index of forest canopies. Forests 2022, 13, 1355. [Google Scholar] [CrossRef]
- Mudi, S.; Paramanik, S.; Behera, M.D.; Prakash, A.J.; Deep, N.R.; Kale, M.P.; Kumar, S.; Sharma, N.; Pradhan, P.; Chavan, M.; et al. Moderate resolution LAI prediction using Sentinel–2 satellite data and indirect field measurements in Sikkim Himalaya. Environ. Monit. Assess. 2022, 194, 897. [Google Scholar] [CrossRef]
- Schleppi, P.; Conedera, M.; Sedivy, I.; Thimonier, A. Correcting non–linearity and slope effects in the estimation of the leaf area index of forests from hemispherical photographs. Agric. For. Meteorol. 2007, 144, 236–242. [Google Scholar] [CrossRef]
- Nobis, M.; Hunziker, U. Automatic thresholding for hemispherical canopy photographs based on edge detection. Agric. For. Meteorol. 2005, 128, 243–250. [Google Scholar] [CrossRef]
- Chen, J.M.; Cihlar, J. Quantifying the effect of canopy architecture on optical measurements of leaf area index using two gap size analysis methods. IEEE Trans. Geosci. Remote Sens. 1995, 33, 777–787. [Google Scholar] [CrossRef]
- Devore, J.; Farnum, N. Applied Statistics for Engineers and Scientists, 1st ed.; Duxbury Press: Belmont, CA, USA, 1999. [Google Scholar]
- Wang, J.; Liu, D.; Ciais, P.; Peñuelas, J. Decreasing rainfall frequency contributes to earlier leaf onset in northern ecosystems. Nat. Clim. Chang. 2022, 12, 386–392. [Google Scholar] [CrossRef]
- Öztürk, M.; Bolat, İ.; Ergün, A. Influence of air–soil temperature on leaf expansion and LAI of Carpinus betulus trees in a temperate urban forest patch. Agric. For. Meteorol. 2015, 200, 185–191. [Google Scholar] [CrossRef]
- Gressler, E.; Jochner, S.; Capdevielle-Vargas, R.M.; Morellato, L.P.C.; Menzel, A. Vertical variation in autumn leaf phenology of Fagus sylvatica L. in southern Germany. Agric. For. Meteorol. 2015, 201, 176–186. [Google Scholar] [CrossRef]
- Lukasová, V.; Lang, M.; Škvarenina, J. Seasonal changes in NDVI in relation to phenological phases, LAI and PAI of beech forests. Balt. For. 2014, 20, 248–262. [Google Scholar]
- Noda, H.M.; Muraoka, H.; Nasahara, K.N. Phenology of leaf optical properties and their relationship to mesophyll development in cool-temperate deciduous broad-leaf trees. Agric. For. Meteorol. 2021, 297, 108236. [Google Scholar] [CrossRef]
- Hase, N.; Doktor, D.; Rebmann, C.; Dechant, B.; Mollenhauer, H.; Cuntz, M. Identifying the main drivers of the seasonal decline of near-infrared reflectance of a temperate deciduous forest. Agric. For. Meteorol. 2022, 313, 108746. [Google Scholar] [CrossRef]
- Vilhar, U.; Roženbergar, D.; Simončič, P.; Diaci, J. Variation in irradiance, soil features and regeneration patterns in experimental forest canopy gaps. Ann. For. Sci. 2015, 72, 253–266. [Google Scholar] [CrossRef]
- Kim, J.; Ryu, Y.; Jiang, C.; Hwang, Y. Continuous observation of vegetation canopy dynamics using an integrated low-cost, near-surface remote sensing system. Agric. For. Meteorol. 2019, 264, 164–177. [Google Scholar] [CrossRef]
- Prévost, M.; Raymond, P. Effect of gap size, aspect and slope on available light and soil temperature after patch-selection cutting in yellow birch–conifer stands, Quebec, Canada. For. Ecol. Manag. 2012, 274, 210–221. [Google Scholar] [CrossRef]
- Horváth, C.V.; Kovács, B.; Tinya, F.; Locatelli, J.S.; Németh, C.; Crecco, L.; Illés, G.; Csépányi, P.; Ódor, P. A matter of size and shape: Microclimatic changes induced by experimental gap openings in a sessile oak–hornbeam forest. Sci. Total Environ. 2023, 873, 162302. [Google Scholar] [CrossRef]
- Klosterman, S.T.; Hufkens, K.; Gray, J.M.; Melaas, E.; Sonnentag, O.; Lavine, I.; Mitchell, L.; Norman, R.; Friedl, M.A.; Richardson, A.D. Evaluating remote sensing of deciduous forest phenology at multiple spatial scales using PhenoCam imagery. Biogeosciences 2014, 11, 4305–4320. [Google Scholar] [CrossRef]
- Nezval, O.; Krejza, J.; Světlík, J.; Šigut, L.; Horáček, P. Comparison of traditional ground-based observations and digital remote sensing of phenological transitions in a floodplain forest. Agric. For. Meteorol. 2020, 291, 108079. [Google Scholar] [CrossRef]
- Hofmeister, J.; Hošek, J.; Brabec, M.; Střalková, R.; Mýlová, P.; Bouda, M.; Pettit, J.L.; Rydval, M.; Svoboda, M. Microclimate edge effect in small fragments of temperate forests in the context of climate change. For. Ecol. Manag. 2019, 448, 48–56. [Google Scholar] [CrossRef]
- Greiser, C.; Hederová, L.; Vico, G.; Wild, J.; Macek, M.; Kopecký, M. Higher soil moisture increases microclimate temperature buffering in temperate broadleaf forests. Agric. For. Meteorol. 2024, 345, 109828. [Google Scholar] [CrossRef]
- Meeussen, C.; Govaert, S.; Vanneste, T.; Bollmann, K.; Brunet, J.; Calders, K.; Cousins, S.A.O.; De Pauw, K.; Diekmann, M.; Gasperini, C.; et al. Microclimatic edge-to-interior gradients of European deciduous forests. Agric. For. Meteorol. 2021, 311, 108699. [Google Scholar] [CrossRef]
- Abd Latif, Z.; Blackburn, G.A. The effects of gap size on some microclimate variables during late summer and autumn in a temperate broadleaved deciduous forest. Int. J. Biometeorol. 2010, 54, 119–129. [Google Scholar] [CrossRef]
- Song, Y.; Ryu, Y. Seasonal changes in vertical canopy structure in a temperate broadleaved forest in Korea. Ecol. Res. 2015, 30, 821–831. [Google Scholar] [CrossRef]
- von Arx, G.; Pannatier, E.G.; Thimonier, A.; Rebetez, M. Microclimate in forests with varying leaf area index and soil moisture: Potential implications for seedling establishment in a changing climate. J. Ecol. 2013, 101, 1201–1213. [Google Scholar] [CrossRef]
- Meijers, E.; Groenewoud, R.; de Vries, J.; van der Zee, J.; Nabuurs, G.J.; Vos, M.; Sterck, F. Canopy cover at the crown-scale best predicts spatial heterogeneity of soil moisture within a temperate Atlantic forest. Agric. For. Meteorol. 2025, 363, 110431. [Google Scholar] [CrossRef]
- Bolat, İ.; Kara, Ö.; Öztürk, M. Short-term changes of leaf area index, light transmission, and gap in a temperate mixed deciduous forest ecosystem in Bartın, Turkey. Turk. J. Agric. For. 2016, 40, 597–605. [Google Scholar] [CrossRef]
- Schleppi, P.; Thimonier, A.; Walthert, L. Estimating leaf area index of mature temperate forests using regressions on site and vegetation data. For. Ecol. Manag. 2011, 261, 601–610. [Google Scholar] [CrossRef]
- Chen, J.; Shao, C.; Jiang, S.; Qu, L.; Zhao, F.; Dong, G. Effects of changes in precipitation on energy and water balance in a Eurasian meadow steppe. Ecol. Process. 2019, 8, 17. [Google Scholar] [CrossRef]
- Wang, X.; Fan, K.; Yan, Y.; Chen, B.; Yan, R.; Xin, X.; Li, L. Controls of Seasonal and Interannual Variations on Soil Respiration in a Meadow Steppe in Eastern Inner Mongolia. Agronomy 2023, 13, 20. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Öztürk, M.; Bolat, İ.; Şensoy, H.; Çakıroğlu, K. Comparison of Canopy–Vegetation Parameters from Interior Parts to Edge of Multi-Story Grove Forest Patch and Meadow Field Within Rural Landscape for Soil Temperature and Moisture. Forests 2025, 16, 904. https://doi.org/10.3390/f16060904
Öztürk M, Bolat İ, Şensoy H, Çakıroğlu K. Comparison of Canopy–Vegetation Parameters from Interior Parts to Edge of Multi-Story Grove Forest Patch and Meadow Field Within Rural Landscape for Soil Temperature and Moisture. Forests. 2025; 16(6):904. https://doi.org/10.3390/f16060904
Chicago/Turabian StyleÖztürk, Melih, İlyas Bolat, Hüseyin Şensoy, and Kamil Çakıroğlu. 2025. "Comparison of Canopy–Vegetation Parameters from Interior Parts to Edge of Multi-Story Grove Forest Patch and Meadow Field Within Rural Landscape for Soil Temperature and Moisture" Forests 16, no. 6: 904. https://doi.org/10.3390/f16060904
APA StyleÖztürk, M., Bolat, İ., Şensoy, H., & Çakıroğlu, K. (2025). Comparison of Canopy–Vegetation Parameters from Interior Parts to Edge of Multi-Story Grove Forest Patch and Meadow Field Within Rural Landscape for Soil Temperature and Moisture. Forests, 16(6), 904. https://doi.org/10.3390/f16060904