Driving Mechanisms and Changes in Dominant Forest Tree Taxa in Europe Under Climate Change

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Taxa Occurrence Records
2.2. Climate, Land Use, and Topographical Factors
2.3. Model Construction
3. Results
3.1. Evaluation of Species Distribution Models
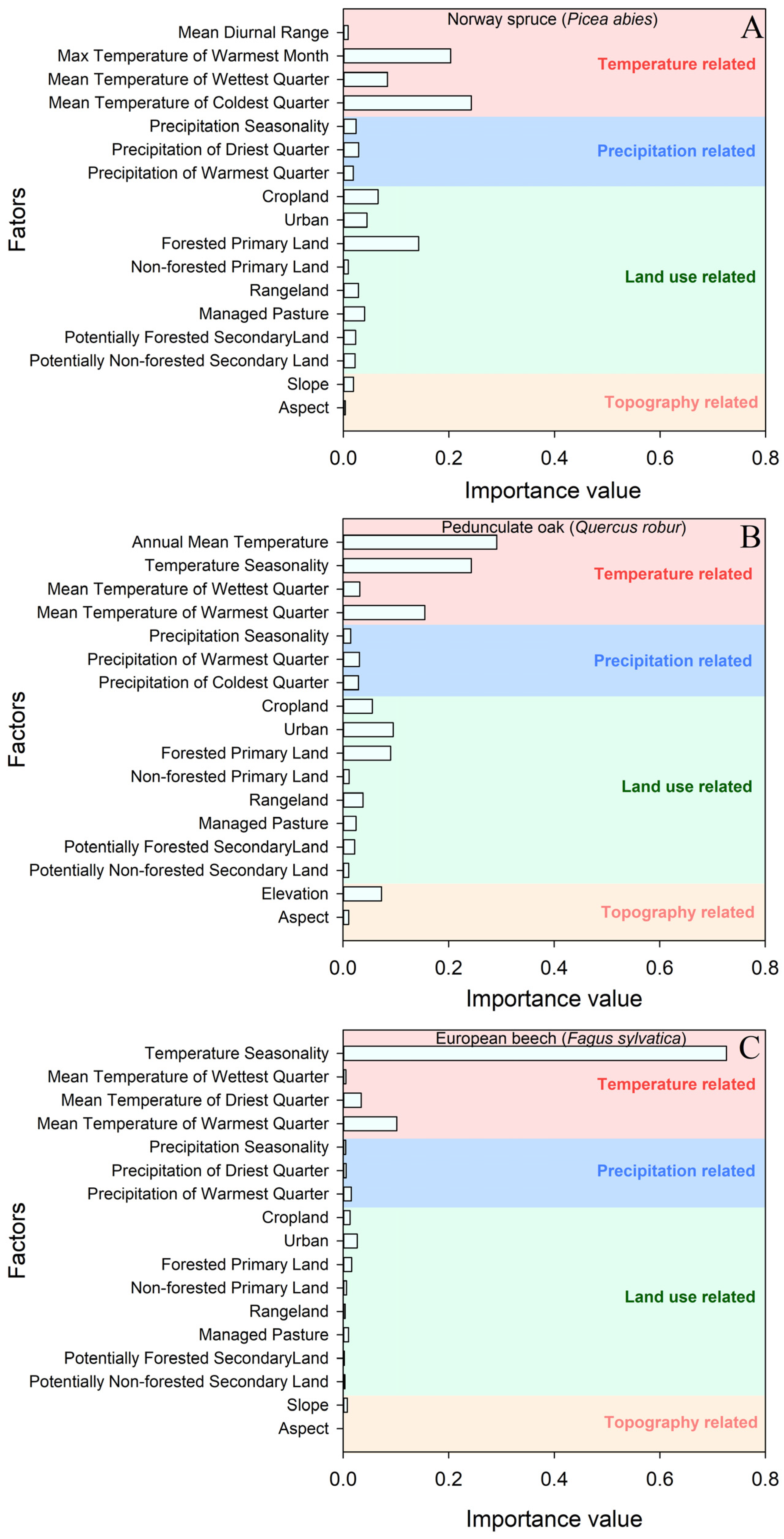
3.2. Driving Factors Affecting Taxa Distributions
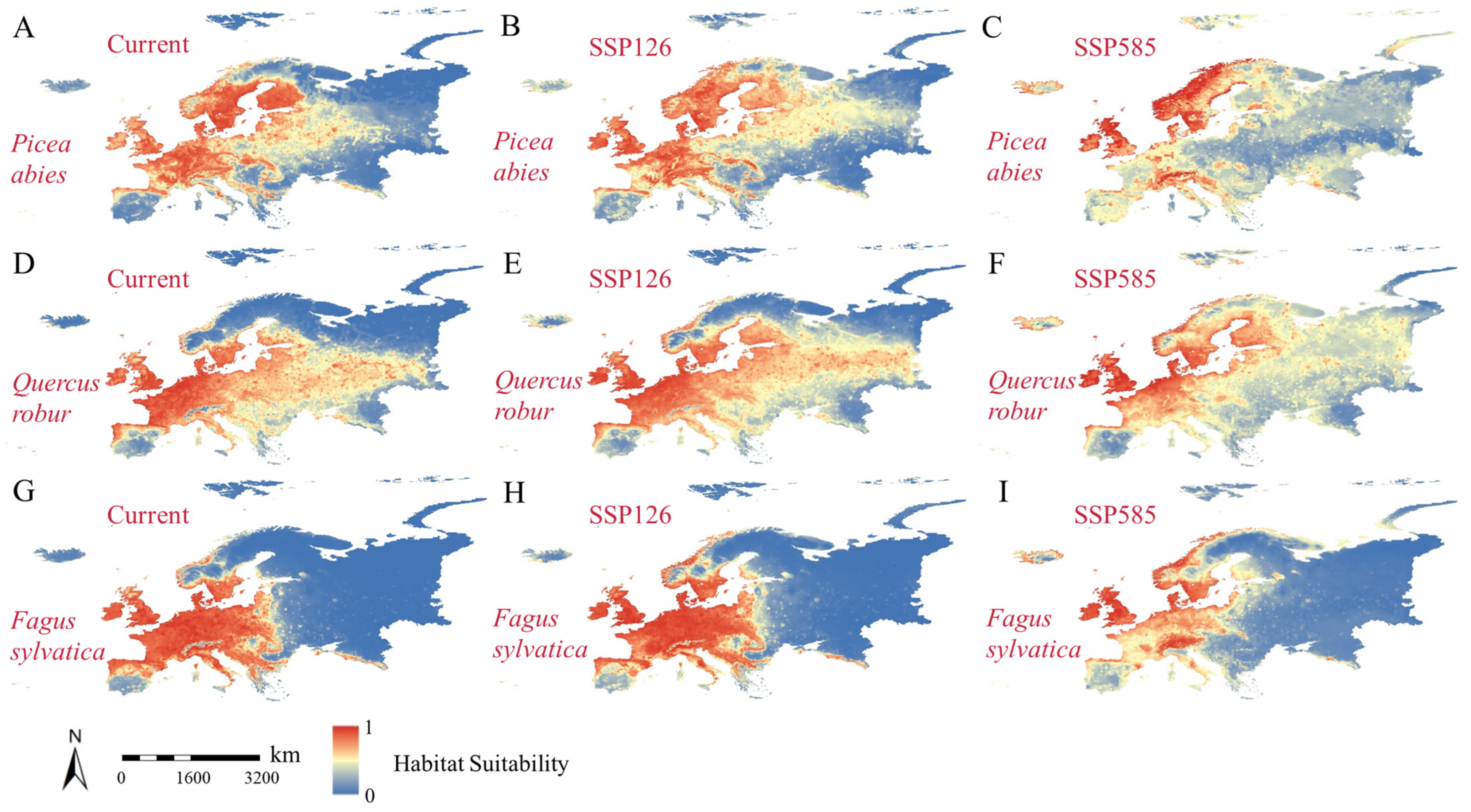
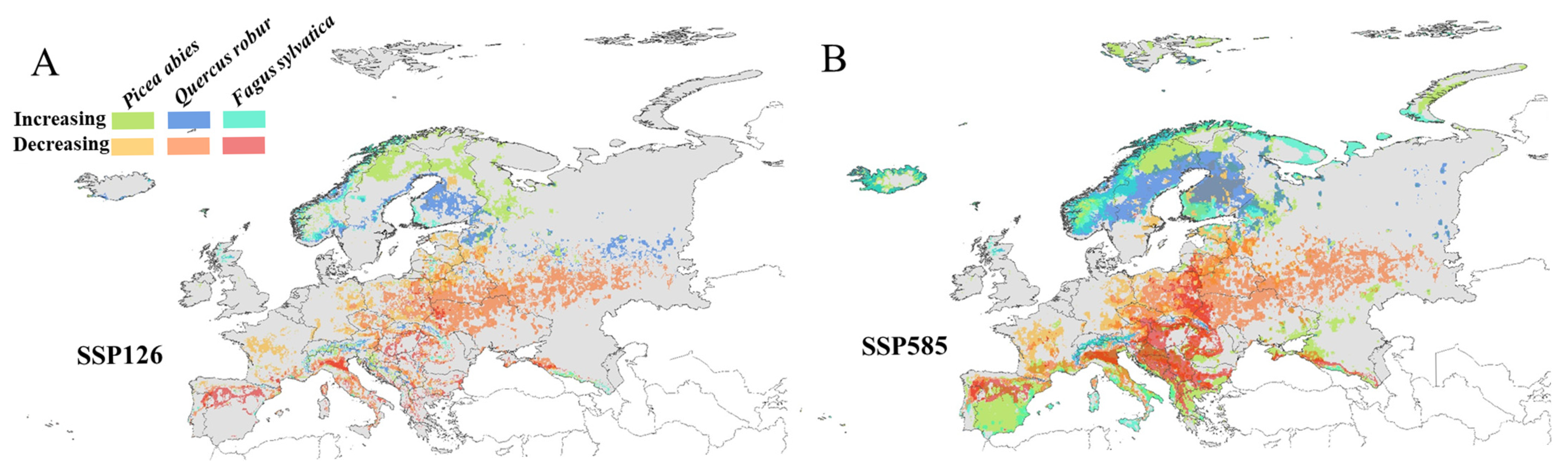
3.3. Changes in Habitat Suitability Under Different Future Scenarios
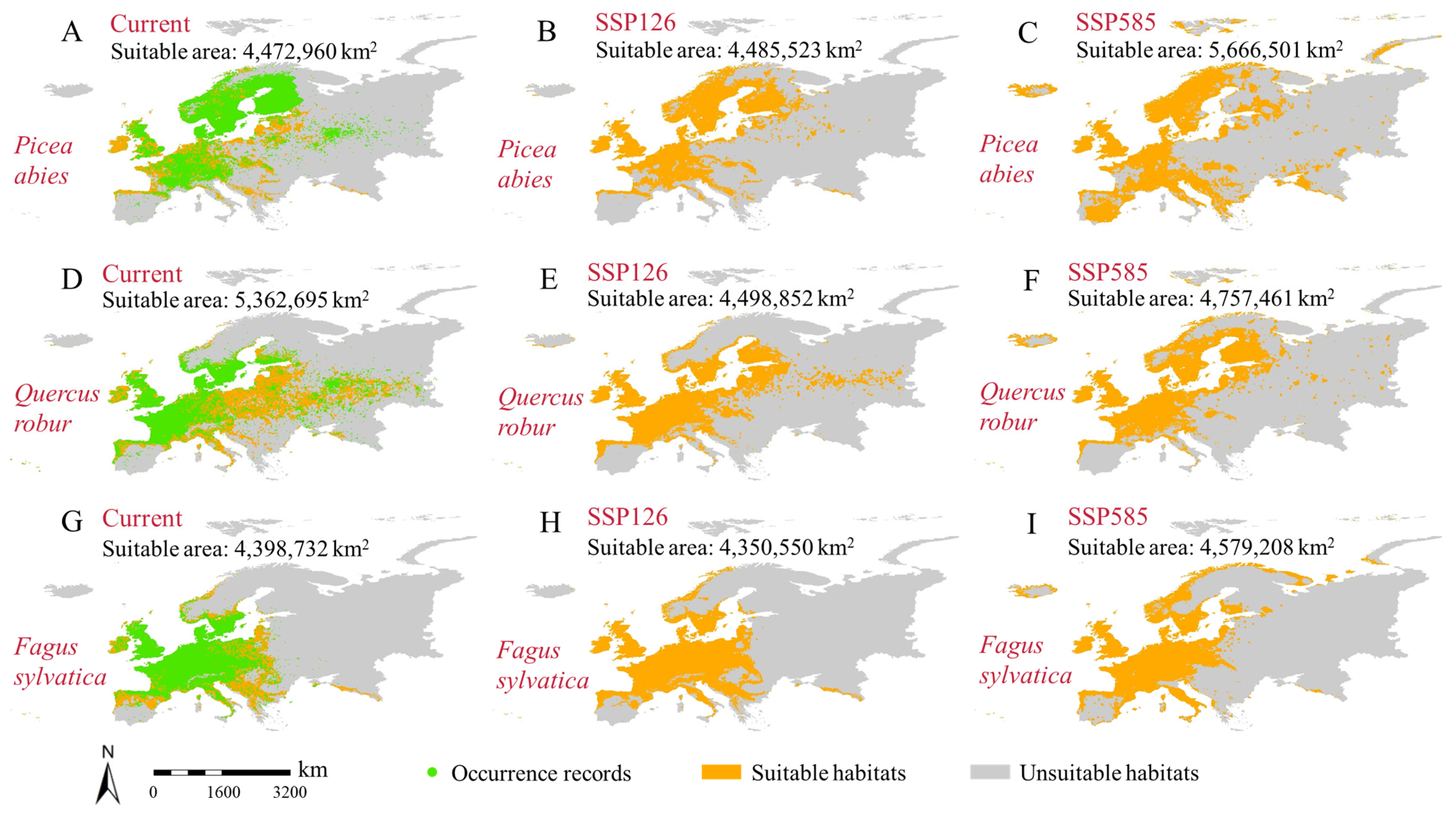
3.4. Distribution and Changes in Suitable Habitats Under Different Future Scenarios
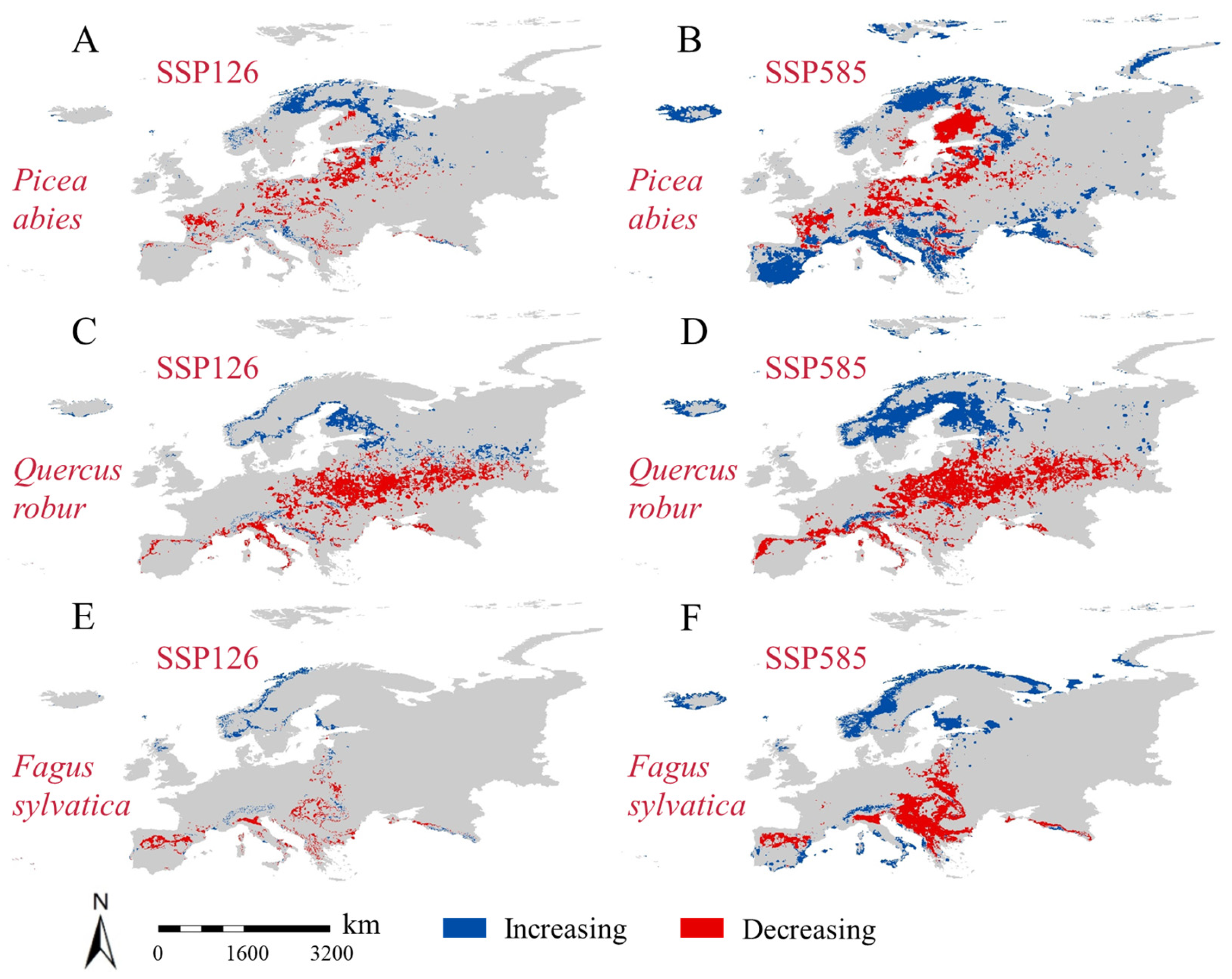
3.5. Changes in Suitable Future Habitats in European Countries for the Three Tree Taxa
4. Discussion
4.1. Driving Factors of Different Tree Taxa Distributions
4.2. Changing Forest Distribution Conditions Under Different Future Scenarios
4.3. Responses of Different European Countries to Future Changes in Suitable Habitat
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Smith, J.B.; Schneider, S.H.; Oppenheimer, M.; Yohe, G.W.; Hare, W.; Mastrandrea, M.D.; Patwardhan, A.; Burton, I.; Corfee-Morlot, J.; Magadza, C.H.D.; et al. Assessing dangerous climate change through an update of the Intergovernmental Panel on Climate Change (IPCC) “reasons for concern”. Proc. Natl. Acad. Sci. USA 2009, 106, 4133–4137. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, J.S.; Lutscher, F.; Bourgault, Y. Climate change fluctuations can increase population abundance and range size. Ecol. Lett. 2024, 27, e14453. [Google Scholar] [CrossRef] [PubMed]
- Pauchard, A.; Milbau, A.; Albihn, A.; Alexander, J.; Burgess, T.; Daehler, C.; Englund, G.; Essl, F.; Evengård, B.; Greenwood, G.B.; et al. Non-native and native organisms moving into high elevation and high latitude ecosystems in an era of climate change: New challenges for ecology and conservation. Biol. Invasions 2016, 18, 345–353. [Google Scholar] [CrossRef]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef]
- Chen, I.C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Leroux, S.J.; Larrivée, M.; Boucher-Lalonde, V.; Hurford, A.; Zuloaga, J.; Kerr, J.T.; Lutscher, F. Mechanistic models for the spatial spread of species under climate change. Ecol. Appl. 2013, 23, 815–828. [Google Scholar] [CrossRef]
- Calama, R.; de-Dios-García, J.; del Río, M.; Madrigal, G.; Gordo, J.; Pardos, M. Mixture mitigates the effect of climate change on the provision of relevant ecosystem services in managed Pinus pinea L. forests. For. Ecol. Manag. 2021, 481, 118782. [Google Scholar] [CrossRef]
- Girdziusas, S.; Lof, M.; Hanssen, K.H.; Lazdina, D.; Madsen, P.; Saksa, T.; Liepins, K.; Floistad, I.S.; Metslaid, M. Forest regeneration management and policy in the Nordic–Baltic region since 1900. Scand. J. For. Res. 2021, 36, 513–523. [Google Scholar] [CrossRef]
- Fuchs, R.; Herold, M.; Verburg, P.H.; Clevers, J.G.; Eberle, J. Gross changes in reconstructions of historic land cover/use for Europe between 1900 and 2010. Glob. Change Biol. 2015, 21, 299–313. [Google Scholar] [CrossRef]
- Kauppi, P.E.; Sandström, V.; Lipponen, A. Forest resources of nations in relation to human well-being. PLoS ONE 2018, 13, e0196248. [Google Scholar] [CrossRef]
- Kelly, A.E.; Goulden, M.L. Rapid shifts in plant distribution with recent climate change. Proc. Natl. Acad. Sci. USA 2008, 105, 11823–11826. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Macua, J.; Ninyerola, M.; Zabala, A.; Domingo-Marimon, C.; Pons, X. Factors affecting forest dynamics in the Iberian Peninsula from 1987 to 2012. The role of topography and drought. For. Ecol. Manag. 2017, 406, 290–306. [Google Scholar] [CrossRef]
- Palmero-Iniesta, M.; Pino, J.; Pesquer, L.; Espelta, J.M. Recent forest area increase in Europe: Expanding and regenerating forests differ in their regional patterns, drivers and productivity trends. Eur. J. For. Res. 2021, 140, 793–805. [Google Scholar] [CrossRef]
- Martinez del Castillo, E.; Zang, C.S.; Buras, A.; Hacket-Pain, A.; Esper, J.; Serrano-Notivoli, R.; Hartl, C.; Weigel, R.; Klesse, S.; Resco de Dios, V.; et al. Climate-change-driven growth decline of European beech forests. Commun. Biol. 2022, 5, 163. [Google Scholar] [CrossRef]
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; Gharun, M.; Grams, T.E.E.; Hauck, M.; Hajek, P.; et al. A first assessment of the impact of the extreme 2018 summer drought on Central European forests. Basic Appl. Ecol. 2020, 45, 86–103. [Google Scholar] [CrossRef]
- Meyer, P.; Spînu, A.P.; Mölder, A.; Bauhus, J. Management alters drought-induced mortality patterns in European beech (Fagus sylvatica L.) forests. Plant Biol. 2022, 24, 1157–1170. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Change Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]
- Antonucci, S.; Santopuoli, G.; Marchetti, M.; Tognetti, R.; Chiavetta, U.; Garfì, V. What is known about the management of European beech forests facing climate change? A review. Curr. For. Rep. 2021, 7, 321–333. [Google Scholar] [CrossRef]
- Boavida, J.; Assis, J.; Silva, I.; Serrão, E.A. Overlooked habitat of a vulnerable gorgonian revealed in the Mediterranean and Eastern Atlantic by ecological niche modelling. Sci. Rep. 2016, 6, 36460. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Gillett, N.P.; Shiogama, H.; Funke, B.; Hegerl, G.; Knutti, R.; Matthes, K.; Santer, B.D.; Stone, D.; Tebaldi, C. The detection and attribution model Intercomparison Project (DAMIP v1.0) contribution to CMIP6. Geosci. Model. Dev. 2016, 9, 3685–3697. [Google Scholar] [CrossRef]
- Li, S.Y.; Miao, L.J.; Jiang, Z.H.; Wang, G.J.; Gnyawali, K.R.; Zhang, J.; Zhang, H.; Fang, K.; He, Y.; Li, C. Projected drought conditions in Northwest China with CMIP6 models under combined SSPs and RCPs for 2015–2099. Adv. Clim. Change Res. 2020, 11, 210–217. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Xu, Z.F.; Han, Y.; Guo, W.D. Evaluation of CMIP6 Models toward dynamical downscaling over 14 CORDEX domains. Clim. Dyn. 2024, 62, 4475–4489. [Google Scholar] [CrossRef]
- Hurtt, G.C.; Chini, L.; Sahajpal, R.; Frolking, S.; Bodirsky, B.L.; Calvin, K.; Doelman, J.C.; Fisk, J.; Fujimori, S.; Goldewijk, K.K.; et al. Harmonization of global land-use change and management for the period 850–2100 (LUH2) for CMIP6. Geosci. Model. Dev. 2020, 13, 5425–5464. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Liu, T.M.; Wang, J.M.; Hu, X.K.; Feng, J.M. Land-use change drives present and future distributions of Fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae). Sci. Total Environ. 2020, 706, 135872. [Google Scholar] [CrossRef]
- Tang, Q.H.; Feng, J.M.; Zong, D.L.; Zhou, J.; Hu, X.K.; Wang, B.R.; Wang, T. Potential spread of desert locust Schistocerca gregagia (Orthoptera: Acrididae) under climate change scenarios. Diversity 2023, 15, 1038. [Google Scholar] [CrossRef]
- Moles, A.T.; Perkins, S.E.; Laffan, S.W.; Flores-Moreno, H.; Awasthy, M.; Tindall, M.L.; Sack, L.; Pitman, A.; Kattge, J.; Aarssen, L.W.; et al. Which is a better predictor of plant traits: Temperature or precipitation? J. Veg. Sci. 2014, 25, 1167–1180. [Google Scholar] [CrossRef]
- Clarke, A.; Gaston, K.J. Climate, energy and diversity. P. Roy. Soc. B.-Biol. Sci. 2006, 273, 2257–2266. [Google Scholar] [CrossRef]
- van Ommen Kloeke, A.E.E.; Douma, J.C.; Ordoñez, J.C.; Reich, P.B.; van Bodegom, P.M. Global quantification of contrasting leaf life span strategies for deciduous and evergreen species in response to environmental conditions. Glob. Ecol. Biogeogr. 2012, 21, 224–235. [Google Scholar] [CrossRef]
- Moles, A.T.; Warton, D.I.; Warman, L.; Swenson, N.G.; Laffan, S.W.; Zanne, A.E.; Pitman, A.; Hemmings, F.A.; Leishman, M.R. Global patterns in plant height. J. Ecol. 2009, 97, 923–932. [Google Scholar] [CrossRef]
- Mosher, E.S.; Silander, J.A.; Latimer, A.M. The role of land-use history in major invasions by woody plant species in the northeastern North American landscape. Biol. Invasions 2009, 11, 2317–2328. [Google Scholar] [CrossRef]
- Thomas, S.M.; Moloney, K.A. Combining the effects of surrounding land-use and propagule pressure to predict the distribution of an invasive plant. Biol. Invasions 2015, 17, 477–495. [Google Scholar] [CrossRef]
- Herrera-Soto, G.; González-Cásares, M.; Pompa-García, M.; Camarero, J.J.; Solís-Moreno, R. Growth of Pinus cembroides Zucc. in Response to Hydroclimatic Variability in Four Sites Forming the Species Latitudinal and Longitudinal Distribution Limits. Forests 2018, 9, 440. [Google Scholar] [CrossRef]
- Noce, S.; Cipriano, C.; Santini, M. Altitudinal shifting of major forest tree species in Italian mountains under climate change. Front. For. Glob. Change 2023, 6, 1250651. [Google Scholar] [CrossRef]
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.; Schall, P.; Spathelf, P.; Rock, J. Adaptive forest management in central Europe: Climate change impacts, strategies and integrative concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Trumbore, S.; Brando, P.; Hartmann, H. Forest health and global change. Science 2015, 349, 814–818. [Google Scholar] [CrossRef]
- Guiot, J.; Cramer, W. Climate change: The 2015 Paris Agreement thresholds and Mediterranean basin ecosystems. Science 2016, 354, 465–468. [Google Scholar] [CrossRef]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Change 2018, 8, 972–980. [Google Scholar] [CrossRef]
- Ruosteenoja, K.; Tuomenvirta, H.; Jylhä, K. GCM-based regional temperature and precipitation change estimates for Europe under four SRES scenarios applying a super-ensemble pattern-scaling method. Clim. Change 2007, 81, 193–208. [Google Scholar] [CrossRef]
- Peltonen-Sainio, P.; Jauhiainen, L.; Palosuo, T.; Hakala, K.; Ruosteenoja, K. Rainfed crop production challenges under European high-latitude conditions. Reg. Environ. Change 2016, 16, 1521–1533. [Google Scholar] [CrossRef]
- Zhou, J.; Tang, Q.H.; Zong, D.L.; Hu, X.K.; Wang, B.R.; Wang, T. Drivers of species distribution and niche dynamics for ornamental plants originating at different latitudes. Diversity 2023, 15, 877. [Google Scholar] [CrossRef]
- Noce, S.; Collalti, A.; Santini, M. Likelihood of changes in forest species suitability, distribution, and diversity under future climate: The case of Southern Europe. Ecol. Evol. 2017, 7, 9358–9375. [Google Scholar] [CrossRef]
- Lindner, M.; Fitzgerald, J.B.; Zimmermann, N.E.; Reyer, C.; Delzon, S.; van der Maaten, E.; Schelhaas, M.; Lasch, P.; Eggers, J.; Maaten-Theunissen, M.; et al. Climate change and European forests: What do we know, what are the uncertainties, and what are the implications for forest management? J. Environ. Manag. 2014, 146, 69–83. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Taxa | Model | AUC Value | TSS Value |
|---|---|---|---|
| Norway spruce | CTA | 0.8053 | 0.5397 |
| FDA | 0.8220 | 0.5120 | |
| GBM | 0.8582 | 0.5633 | |
| GLM | 0.8060 | 0.4848 | |
| MAXNET | 0.8330 | 0.5293 | |
| RF | 0.9999 | 0.9952 | |
| Integrated model | 0.8765 | 0.5853 | |
| pedunculate oak | CTA | 0.8145 | 0.5253 |
| FDA | 0.8060 | 0.4567 | |
| GBM | 0.8395 | 0.5163 | |
| GLM | 0.7953 | 0.4287 | |
| MAXNET | 0.8055 | 0.4478 | |
| RF | 0.9999 | 0.9947 | |
| Integrated model | 0.8285 | 0.5058 | |
| European beech | CTA | 0.8438 | 0.6623 |
| FDA | 0.8728 | 0.6473 | |
| GBM | 0.9108 | 0.6800 | |
| GLM | 0.8705 | 0.6433 | |
| MAXNET | 0.8773 | 0.6435 | |
| RF | 0.9999 | 0.9960 | |
| Integrated model | 0.9054 | 0.6819 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Tang, Q.; Zhao, Y.; Hu, X.; Wang, T.; Wang, B. Driving Mechanisms and Changes in Dominant Forest Tree Taxa in Europe Under Climate Change. Forests 2025, 16, 900. https://doi.org/10.3390/f16060900
Zhou J, Tang Q, Zhao Y, Hu X, Wang T, Wang B. Driving Mechanisms and Changes in Dominant Forest Tree Taxa in Europe Under Climate Change. Forests. 2025; 16(6):900. https://doi.org/10.3390/f16060900
Chicago/Turabian StyleZhou, Jing, Qianhong Tang, Yanan Zhao, Xiaokang Hu, Tao Wang, and Bingru Wang. 2025. "Driving Mechanisms and Changes in Dominant Forest Tree Taxa in Europe Under Climate Change" Forests 16, no. 6: 900. https://doi.org/10.3390/f16060900
APA StyleZhou, J., Tang, Q., Zhao, Y., Hu, X., Wang, T., & Wang, B. (2025). Driving Mechanisms and Changes in Dominant Forest Tree Taxa in Europe Under Climate Change. Forests, 16(6), 900. https://doi.org/10.3390/f16060900