
Assessing Climate Change Risks and Conservation Needs for Carpinus Species in China Using Ensemble Distribution Modeling

 ,
,
Abstract
1. Introduction
2. Material and Methods
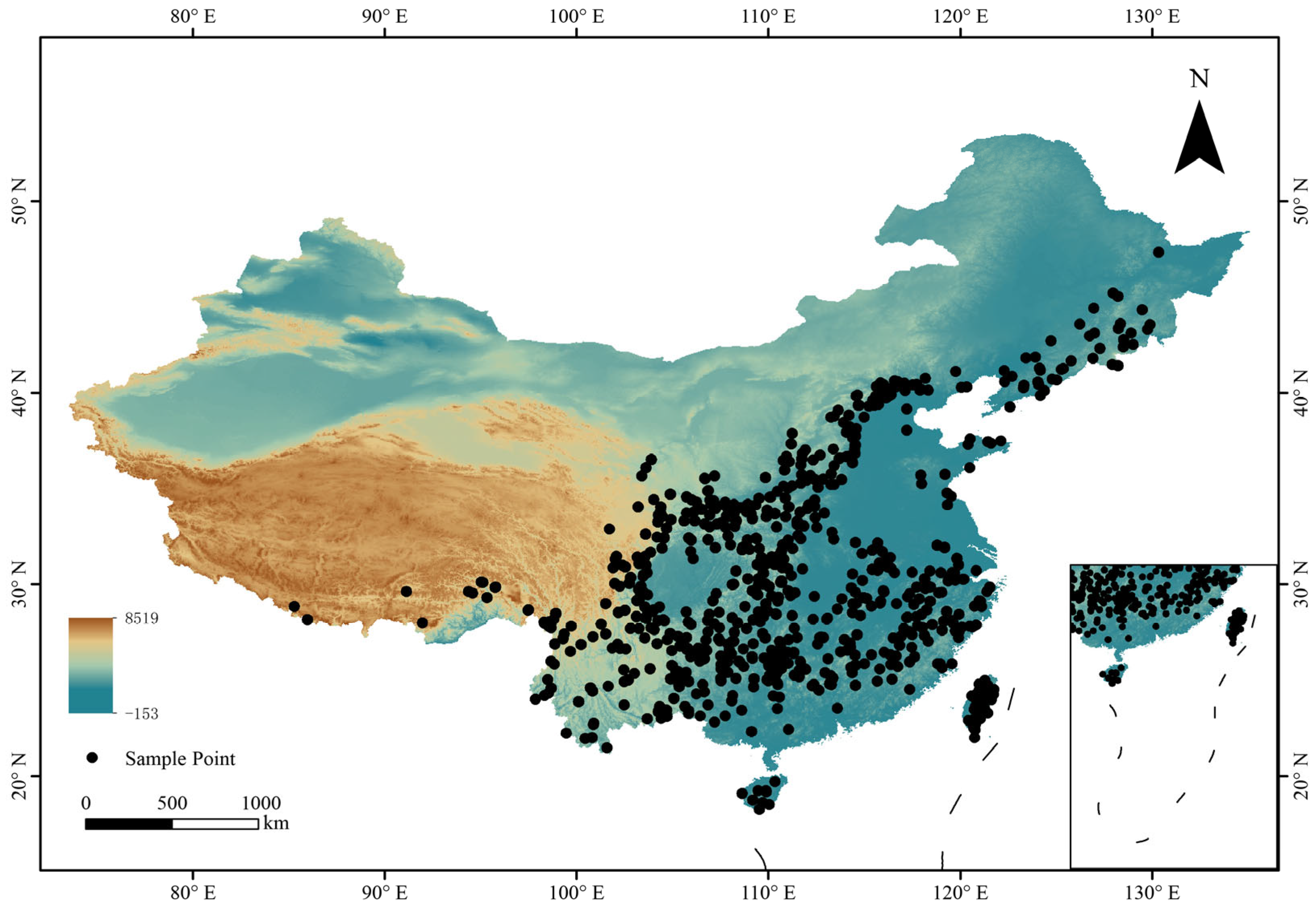
2.1. Species Distribution Data
2.2. Environmental Variables
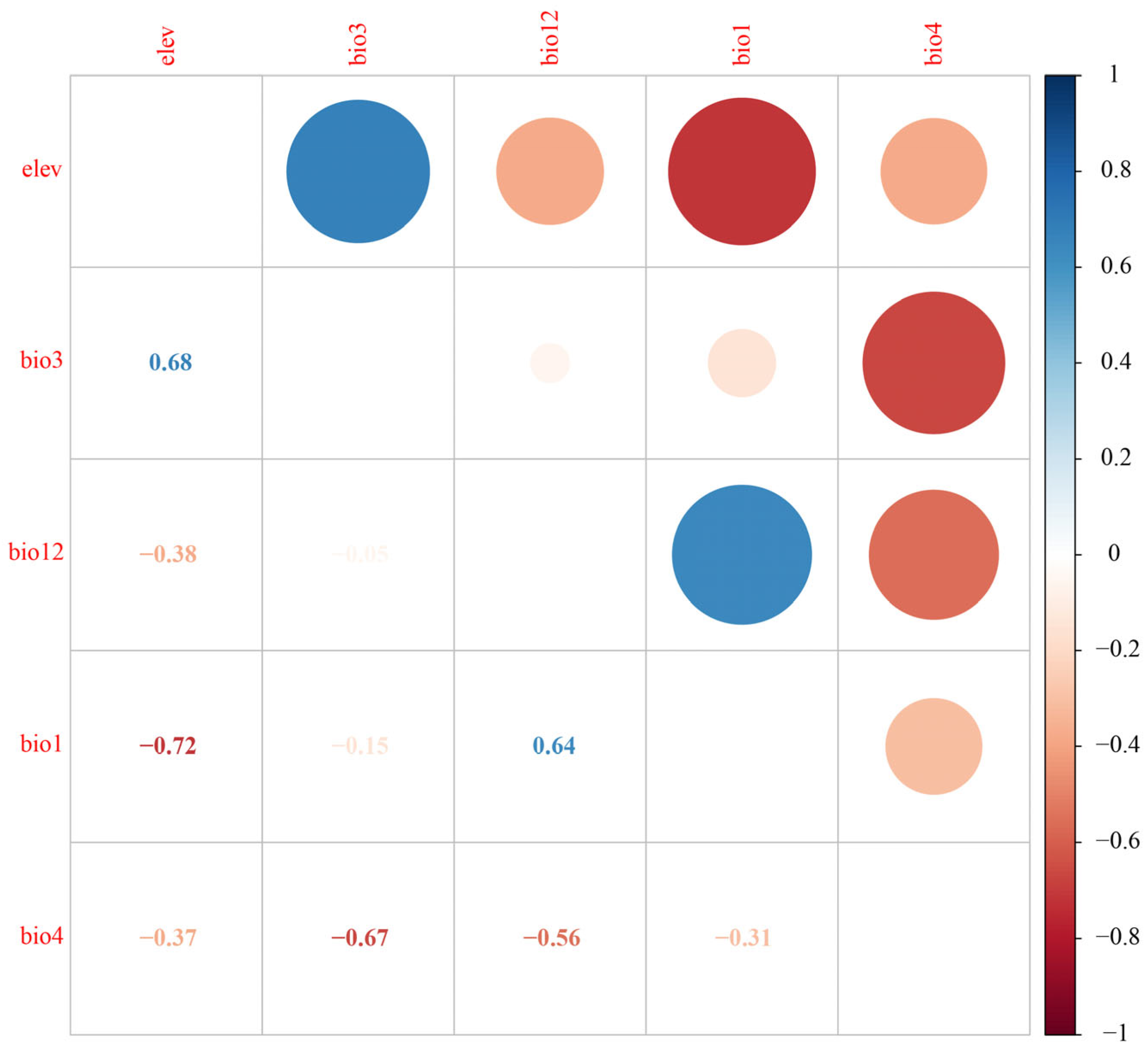
2.3. Key Environmental Variable Selection
2.4. Species Distribution Modeling
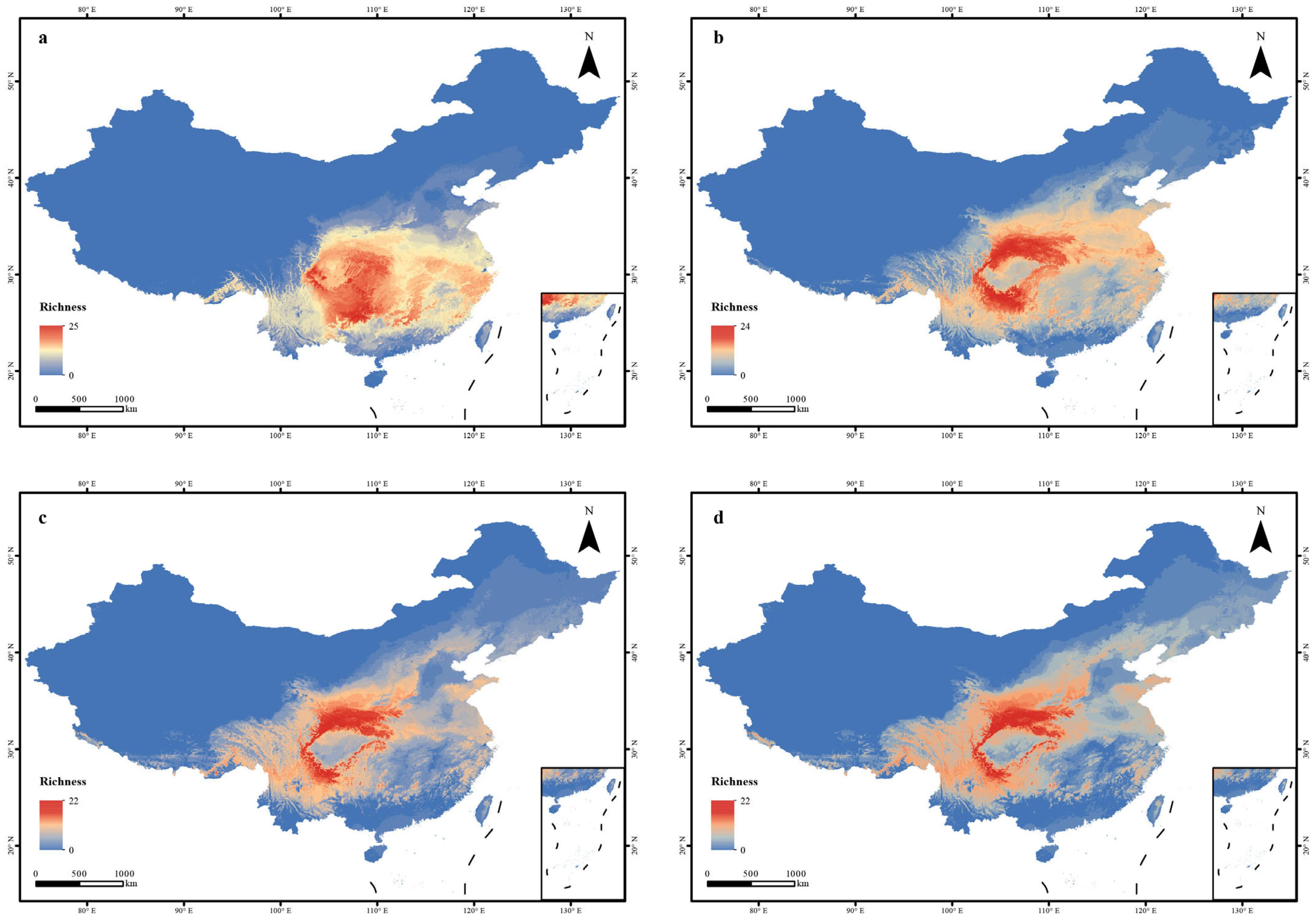
2.5. Species Richness Visualization
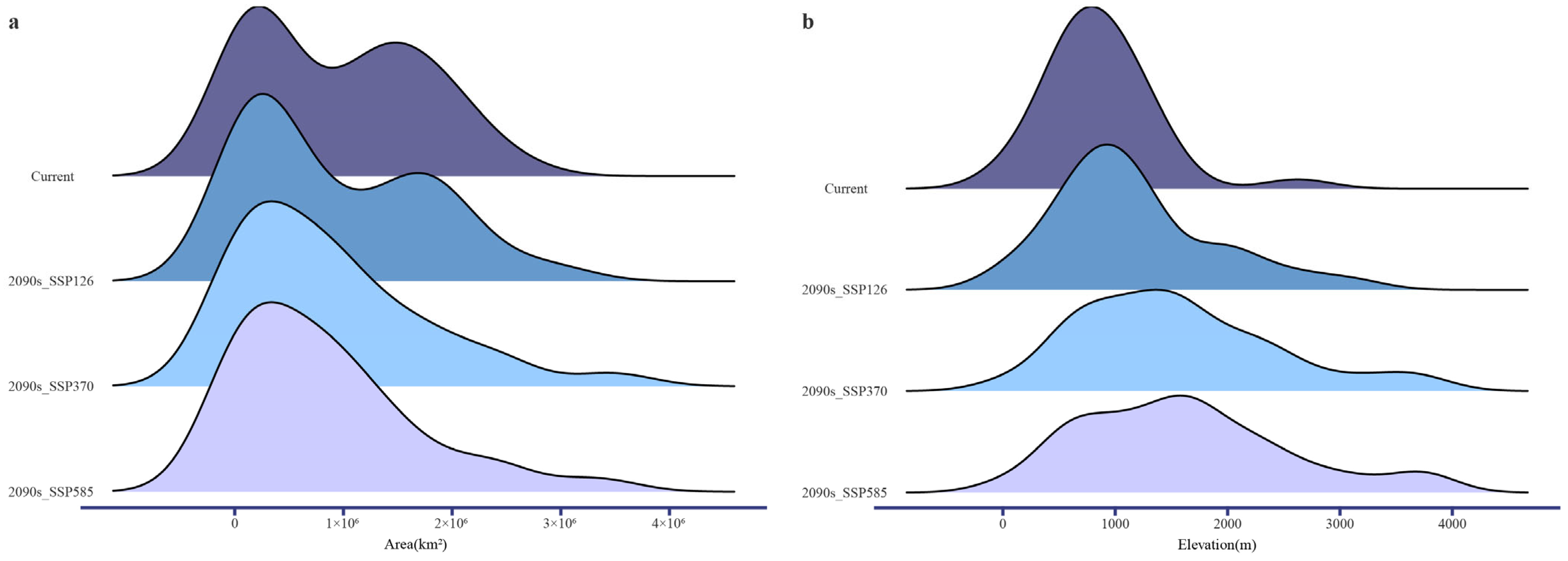
2.6. Analysis of Projected Range Area and Elevational Change
2.7. Centroid Shift Analysis
3. Results
3.1. Model Accuracy and Key Environmental Variables
3.2. Predicted Changes in Species Distributions Under Future Climate Scenarios
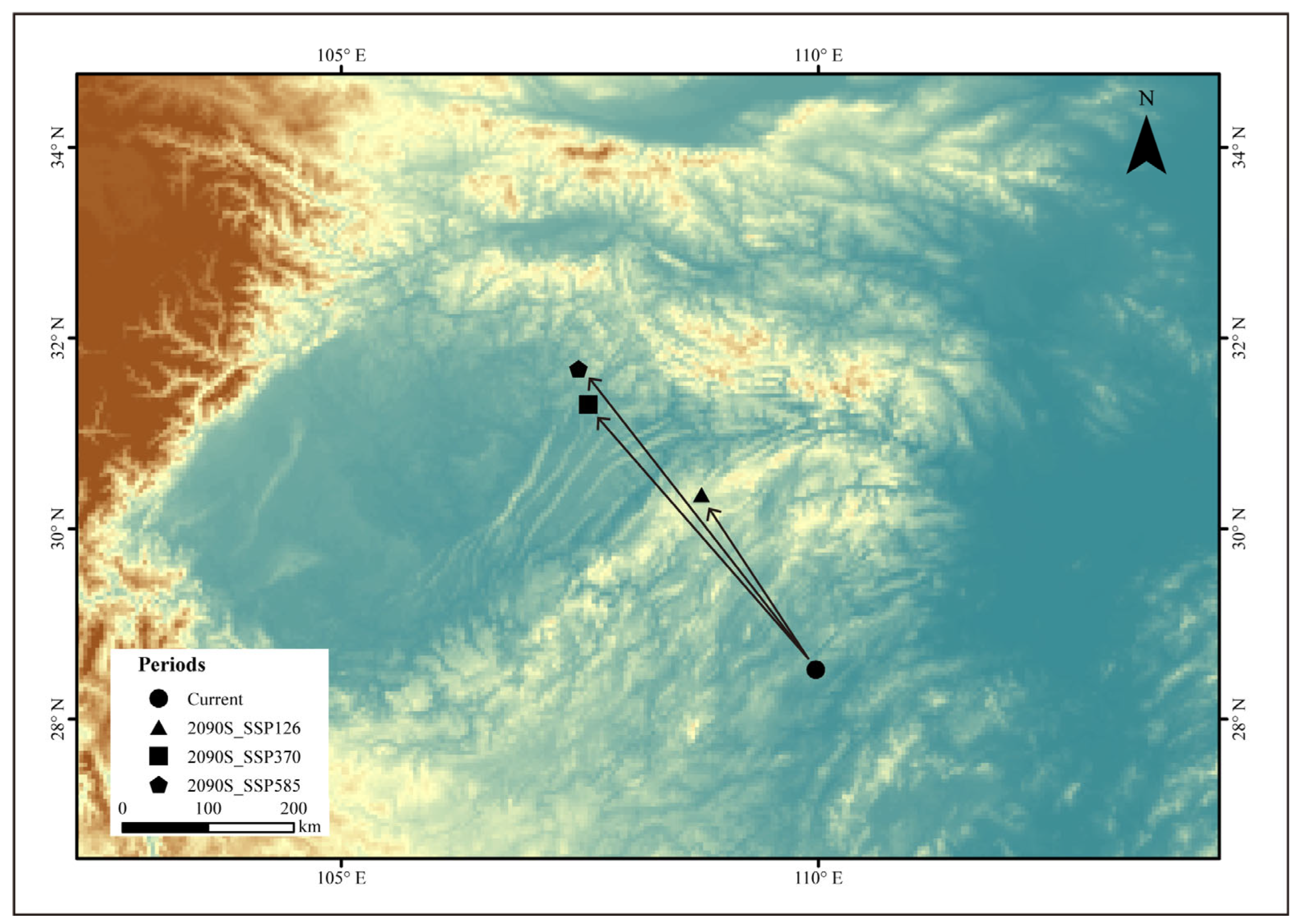
3.3. Centroid Shift Under Climatic Scenarios
4. Discussion
4.1. Model Performance and Advantages of Ensemble Modeling
4.2. Climate-Driven Shifts in Suitable Habitat and Key Environmental Drivers
4.3. Climate-Associated Habitat Loss and Conservation Risk in Vulnerable Species
4.4. Limitations and Future Research Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Intergovernmental Panel On Climate Change (IPCC). Climate Change 2021—The Physical Science Basis: Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change, 1st ed.; Cambridge University Press: Cambridge, UK, 2023; ISBN 978-1-009-15789-6. [Google Scholar]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid Range Shifts of Species Associated with High Levels of Climate Warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef] [PubMed]
- Vitasse, Y.; Ursenbacher, S.; Klein, G.; Bohnenstengel, T.; Chittaro, Y.; Delestrade, A.; Monnerat, C.; Rebetez, M.; Rixen, C.; Strebel, N.; et al. Phenological and Elevational Shifts of Plants, Animals and Fungi under Climate Change in the European Alps. Biol. Rev. 2021, 96, 1816–1835. [Google Scholar] [CrossRef] [PubMed]
- Pacifici, M.; Foden, W.B.; Visconti, P.; Watson, J.E.M.; Butchart, S.H.M.; Kovacs, K.M.; Scheffers, B.R.; Hole, D.G.; Martin, T.G.; Akçakaya, H.R.; et al. Assessing Species Vulnerability to Climate Change. Nat. Clim. Change 2015, 5, 215–224. [Google Scholar] [CrossRef]
- He, F.; Zarfl, C.; Bremerich, V.; David, J.N.W.; Hogan, Z.; Kalinkat, G.; Tockner, K.; Jähnig, S.C. The Global Decline of Freshwater Megafauna. Glob. Change Biol. 2019, 25, 3883–3892. [Google Scholar] [CrossRef]
- Chichorro, F.; Urbano, F.; Teixeira, D.; Väre, H.; Pinto, T.; Brummitt, N.; He, X.; Hochkirch, A.; Hyvönen, J.; Kaila, L.; et al. Species Traits Predict Extinction Risk across the Tree of Life. bioRxiv 2020, bioRxiv:2020.07.01.183053. [Google Scholar]
- Wan, J.-N.; Mbari, N.J.; Wang, S.-W.; Liu, B.; Mwangi, B.N.; Rasoarahona, J.R.E.; Xin, H.-P.; Zhou, Y.-D.; Wang, Q.-F. Modeling Impacts of Climate Change on the Potential Distribution of Six Endemic Baobab Species in Madagascar. Plant Divers. 2021, 43, 117–124. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, Y.; Yue, J.; Wang, Z.; Zou, H.; Ji, X.; Zhang, S.; Liu, Z. Prediction of Potential Suitable Areas and Priority Protection for Cupressus gigantea on the Tibetan Plateau. Plants 2024, 13, 896. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The Effect of Sample Size and Species Characteristics on Performance of Different Species Distribution Modeling Methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Franklin, J.; Miller, J.A. Mapping Species Distributions: Spatial Inference and Prediction, 5th ed.; Ecology, biodiversity and conservation; Cambridge University Press: Cambridge, UK, 2011; ISBN 978-0-521-70002-3. [Google Scholar]
- Araújo, M.B.; Peterson, A.T. Uses and Misuses of Bioclimatic Envelope Modeling. Ecology 2012, 93, 1527–1539. [Google Scholar] [CrossRef]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibañez, I.; Miller, L.P.; et al. Global Threats from Invasive Alien Species in the Twenty-First Century and National Response Capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef]
- Raitsos, D.E.; Beaugrand, G.; Georgopoulos, D.; Zenetos, A.; Pancucci-Papadopoulou, A.M.; Theocharis, A.; Papathanassiou, E. Global Climate Change Amplifies the Entry of Tropical Species into the Eastern Mediterranean Sea. Limnol. Oceanogr. 2010, 55, 1478–1484. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Wang, R.; Guo, L.; Ji, Y.; Chen, Y.; Hao, L.; Lin, K. Impacts of Human Activity and Climate Change on the Suitable Habitats for Xanthium Spinosum in China. Plants 2025, 14, 306. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists: Statistical Explanation of MaxEnt. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble Forecasting of Species Distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Cassey, P.; Blackburn, T.M. The More You Introduce the More You Get: The Role of Colonization Pressure and Propagule Pressure in Invasion Ecology. Divers. Distrib. 2009, 15, 904–910. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Guillera-Arroita, G.; Lahoz-Monfort, J.J. A Review of Evidence about Use and Performance of Species Distribution Modelling Ensembles like BIOMOD. Divers. Distrib. 2019, 25, 839–852. [Google Scholar] [CrossRef]
- Hao, T.; Elith, J.; Lahoz-Monfort, J.J.; Guillera-Arroita, G. Testing Whether Ensemble Modelling Is Advantageous for Maximising Predictive Performance of Species Distribution Models. Ecography 2020, 43, 549–558. [Google Scholar] [CrossRef]
- Harris, J.; Pirtle, J.L.; Laman, E.A.; Siple, M.C.; Thorson, J.T. An Ensemble Approach to Species Distribution Modelling Reconciles Systematic Differences in Estimates of Habitat Utilization and Range Area. J. Appl. Ecol. 2024, 61, 351–364. [Google Scholar] [CrossRef]
- Yang, J.; Fu, Z.; Xiao, K.; Dong, H.; Zhou, Y.; Zhan, Q. Climate Change Potentially Leads to Habitat Expansion and Increases the Invasion Risk of Hydrocharis (Hydrocharitaceae). Plants 2023, 12, 4124. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A Platform for Ensemble Forecasting of Species Distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Vasconcelos, R.N.; Cantillo-Pérez, T.; Franca Rocha, W.J.S.; Aguiar, W.M.; Mendes, D.T.; de Jesus, T.B.; de Santana, C.O.; de Santana, M.M.M.; Oliveira, R.P. Advances and Challenges in Species Ecological Niche Modeling: A Mixed Review. Earth 2024, 5, 963–989. [Google Scholar] [CrossRef]
- Cheng, J.; Xiao, J.; Song, N.; Saha, S.; Qin, J.; Nomura, H.; Panhwar, S.K.; Farooq, N.; Shao, K.; Gao, T. Molecular Phylogeny Reveals Cryptic Diversity and Swim Bladder Evolution of Sillaginidae Fishes (Perciformes) across the Indo-West Pacific Ocean. Divers. Distrib. 2021, 27, 82–94. [Google Scholar] [CrossRef]
- Li, P.C.; Skvortsov, A.K. Betulaceae. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press & Missouri Botanical Garden Press: Beijing, China; St. Louis, MO, USA, 1999; Volume 4, pp. 289–300. [Google Scholar]
- Xue, L.; Jia, L.; Nam, G.; Huang, Y.; Zhang, S.; Wang, Y.; Zhou, Z.; Chen, Y. Involucre Fossils of Carpinus, a Northern Temperate Element, from the Miocene of China and the Evolution of Its Species Diversity in East Asia. Plant Divers. 2020, 42, 155–167. [Google Scholar] [CrossRef]
- Dong, C.; Lu, Z.; Zhang, H.; Liu, J.; Li, M. Delimiting 33 Carpinus (Betulaceae) Species with a Further Phylogenetic Inference. AoB Plants 2022, 14, plac006. [Google Scholar] [CrossRef]
- Zhao, R.; Chu, X.; He, Q.; Tang, Y.; Song, M.; Zhu, Z. Modeling Current and Future Potential Geographical Distribution of Carpinus tientaiensis, a Critically Endangered Species from China. Forests 2020, 11, 774. [Google Scholar] [CrossRef]
- Sheng, Q.; Song, M.; Zhu, Z.; Cao, F. Physiological and Biochemical Responses of Two Precious Carpinus Species to High-Concentration NO2 Stress and Their Natural Recovery. Sci. Rep. 2021, 11, 9500. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Cai, L.; Liu, D.; Chen, G.; Gratzfeld, J.; Sun, W. China’s Conservation Program on Plant Species with Extremely Small Populations (PSESP): Progress and Perspectives. Biol. Conserv. 2020, 244, 108535. [Google Scholar] [CrossRef]
- He, Q.; Zhao, R.; Zhu, Z. Geographical Distribution Simulation and Comparative Analysis of Carpinus viminea and C. londoniana. Glob. Ecol. Conserv. 2020, 21, e00825. [Google Scholar] [CrossRef]
- Gülçin, D.; Arslan, E.S.; Örücü, Ö.K. Effects of Climate Change on the Ecological Niche of Common Hornbeam (Carpinus betulus L.). Ecol. Inform. 2021, 66, 101478. [Google Scholar] [CrossRef]
- Sun, J.; Feng, L.; Wang, T.; Tian, X.; He, X.; Xia, H.; Wang, W. Predicting the Potential Habitat of Three Endangered Species of Carpinus Genus under Climate Change and Human Activity. Forests 2021, 12, 1216. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A Review of Methods to Deal with It and a Simulation Study Evaluating Their Performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial Filtering to Reduce Sampling Bias Can Improve the Performance of Ecological Niche Models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological Niche Modeling in Maxent: The Importance of Model Complexity and the Performance of Model Selection Criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Marmion, M.; Parviainen, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. Evaluation of Consensus Methods in Predictive Species Distribution Modelling. Divers. Distrib. 2009, 15, 59–69. [Google Scholar] [CrossRef]
- El Alaoui, O.; Idri, A. Predicting the Potential Distribution of Wheatear Birds Using Stacked Generalization-Based Ensembles. Ecol. Inform. 2023, 75, 102084. [Google Scholar] [CrossRef]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A Significant Upward Shift in Plant Species Optimum Elevation During the 20th Century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- Moran, E.V.; Thuiller, W.; Angert, A.L.; Benito Garzón, M. Editorial: Predicting and Managing Climate-Driven Range Shifts in Plants. Front. Ecol. Evol. 2022, 10, 856213. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N.E. Predictive Habitat Distribution Models in Ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Hirzel, A.H.; Le Lay, G. Habitat Suitability Modelling and Niche Theory. J. Appl. Ecol. 2008, 45, 1372–1381. [Google Scholar] [CrossRef]
- Freeman, B.G.; Scholer, M.N.; Ruiz-Gutierrez, V.; Fitzpatrick, J.W. Climate Change Causes Upslope Shifts and Mountaintop Extirpations in a Tropical Bird Community. Proc. Natl. Acad. Sci. USA 2018, 115, 11982–11987. [Google Scholar] [CrossRef] [PubMed]
- Steinbauer, M.J.; Grytnes, J.-A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated Increase in Plant Species Richness on Mountain Summits Is Linked to Warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Variable | Unit |
|---|---|---|
| bio1 | Annual mean temperature | °C |
| bio3 | Isothermality | |
| bio4 | Temperature seasonality | |
| bio12 | Annual precipitation | mm |
| elev | Elevation | m |
| Taxa | Number | Taxa | Number |
|---|---|---|---|
| C. chuniana | 25 | C. monbeigiana | 41 |
| C. cordata | 161 | C. oblongifolia | 5 |
| C. cordata var. chinensis | 82 | C. omeiensis | 6 |
| C. cordata var. mollis | 28 | C. polyneura | 85 |
| C. dayongiana | 5 | C. polyneura var. sunpanensis | 9 |
| C. fangiana | 66 | C. pubescens | 101 |
| C. fargesiana | 96 | C. purpurinervis | 9 |
| C. fargesiana var. hwai | 15 | C. rankanensis | 23 |
| C. firmifolia | 5 | C. rupestris | 9 |
| C. henryana | 40 | C. shensiensis | 14 |
| C. hupeana | 41 | C. stipulata | 23 |
| C. kawakamii | 77 | C. tsaiana | 10 |
| C. kweichowensis | 64 | C. tschonoskii | 58 |
| C. londoniana | 91 | C. turczaninovii | 229 |
| C. londoniana var. lanceolata | 9 | C. viminea | 282 |
| C. mollicoma | 8 | C. viminea var. chiukiangensis | 8 |
| SSP126 | SSP370 | SSP585 | |
|---|---|---|---|
| Expansion | 12 | 8 | 7 |
| Contraction | 20 | 22 | 23 |
| Extinction | 0 | 2 | 2 |
| SSP126 | SSP370 | SSP585 | |
|---|---|---|---|
| Increase | 29 | 29 | 29 |
| Decrease | 3 | 1 | 1 |
| Extinction | 0 | 2 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.; Fu, C.; Zhao, Z.; Zhang, W.; Yang, X.; Hu, Q.; Wang, Z. Assessing Climate Change Risks and Conservation Needs for Carpinus Species in China Using Ensemble Distribution Modeling. Forests 2025, 16, 888. https://doi.org/10.3390/f16060888
Yang W, Fu C, Zhao Z, Zhang W, Yang X, Hu Q, Wang Z. Assessing Climate Change Risks and Conservation Needs for Carpinus Species in China Using Ensemble Distribution Modeling. Forests. 2025; 16(6):888. https://doi.org/10.3390/f16060888
Chicago/Turabian StyleYang, Wenjie, Chenlong Fu, Zhuang Zhao, Wenjing Zhang, Xiaoyue Yang, Quanjun Hu, and Zefu Wang. 2025. "Assessing Climate Change Risks and Conservation Needs for Carpinus Species in China Using Ensemble Distribution Modeling" Forests 16, no. 6: 888. https://doi.org/10.3390/f16060888
APA StyleYang, W., Fu, C., Zhao, Z., Zhang, W., Yang, X., Hu, Q., & Wang, Z. (2025). Assessing Climate Change Risks and Conservation Needs for Carpinus Species in China Using Ensemble Distribution Modeling. Forests, 16(6), 888. https://doi.org/10.3390/f16060888