
Projected Spatial Distribution Patterns of Three Dominant Desert Plants in Xinjiang of Northwest China



Abstract
1. Introduction
2. Materials and Methods
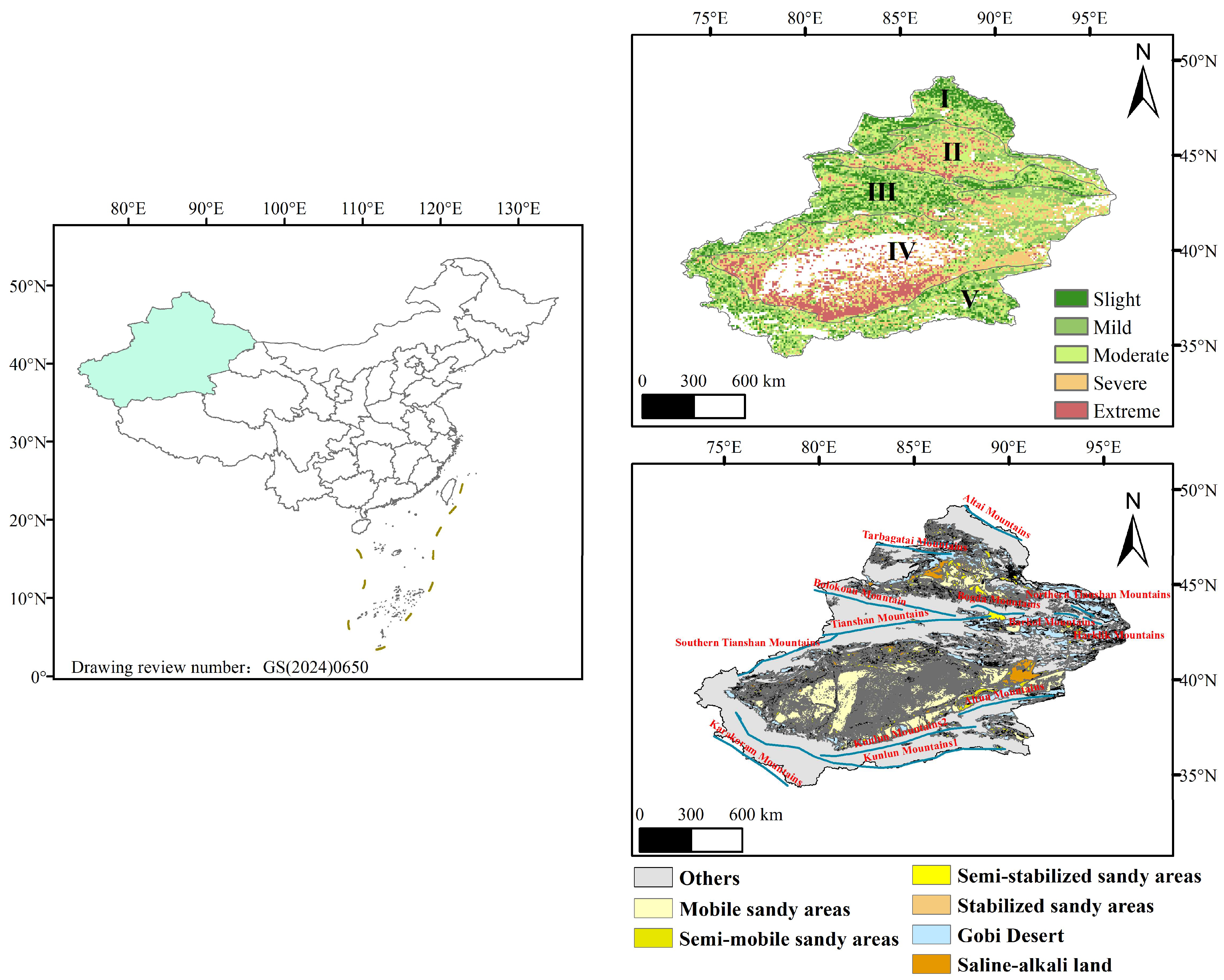
2.1. Study Area Overview
2.2. Species Occurrence Data Collection
2.3. Environmental Variable Acquisition
2.4. Variable Screening
2.5. Model Parameterization and Performance Evaluation
2.6. Habitat Suitability Zonation
3. Results
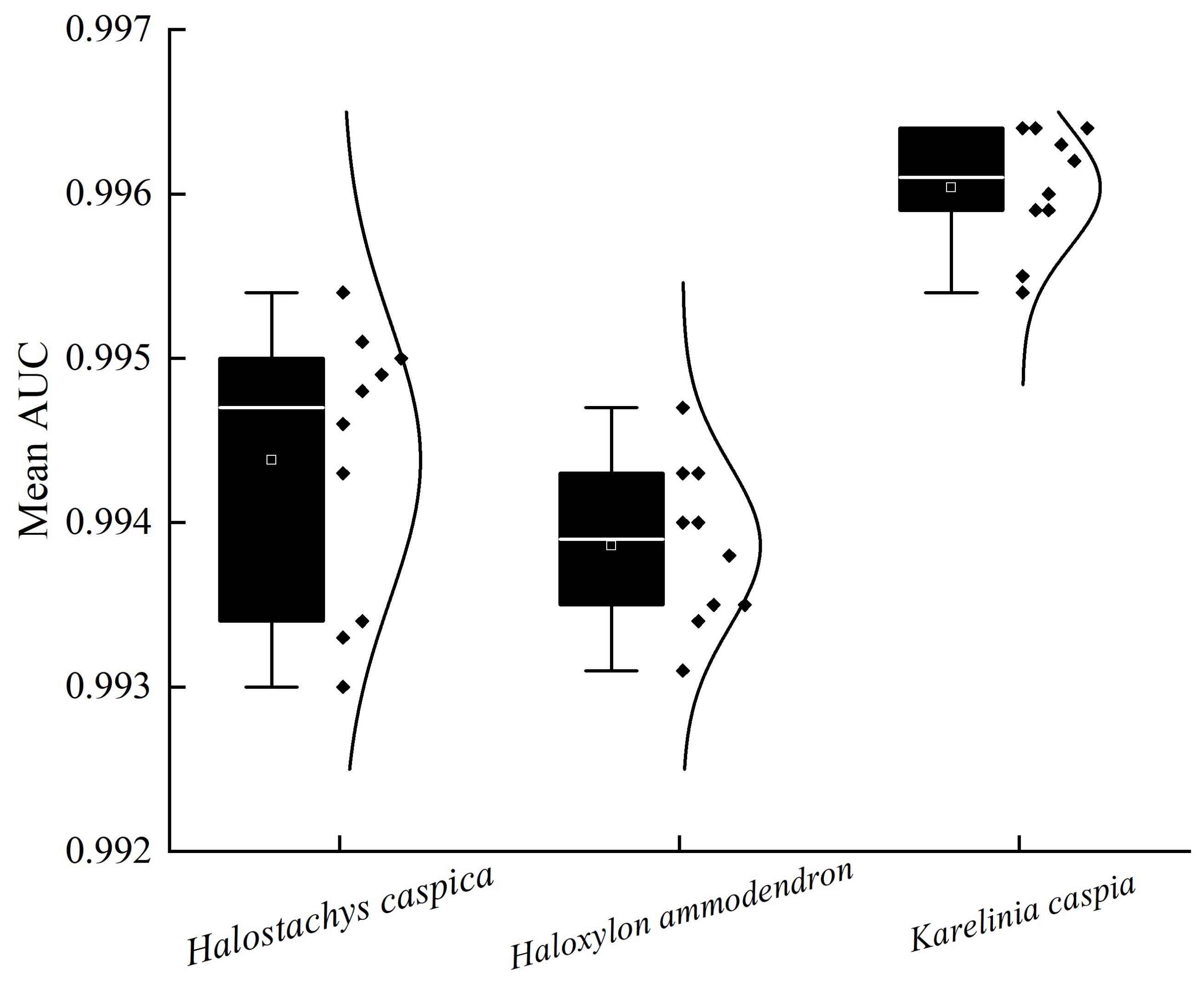
3.1. Model Predictive Accuracy Evaluation
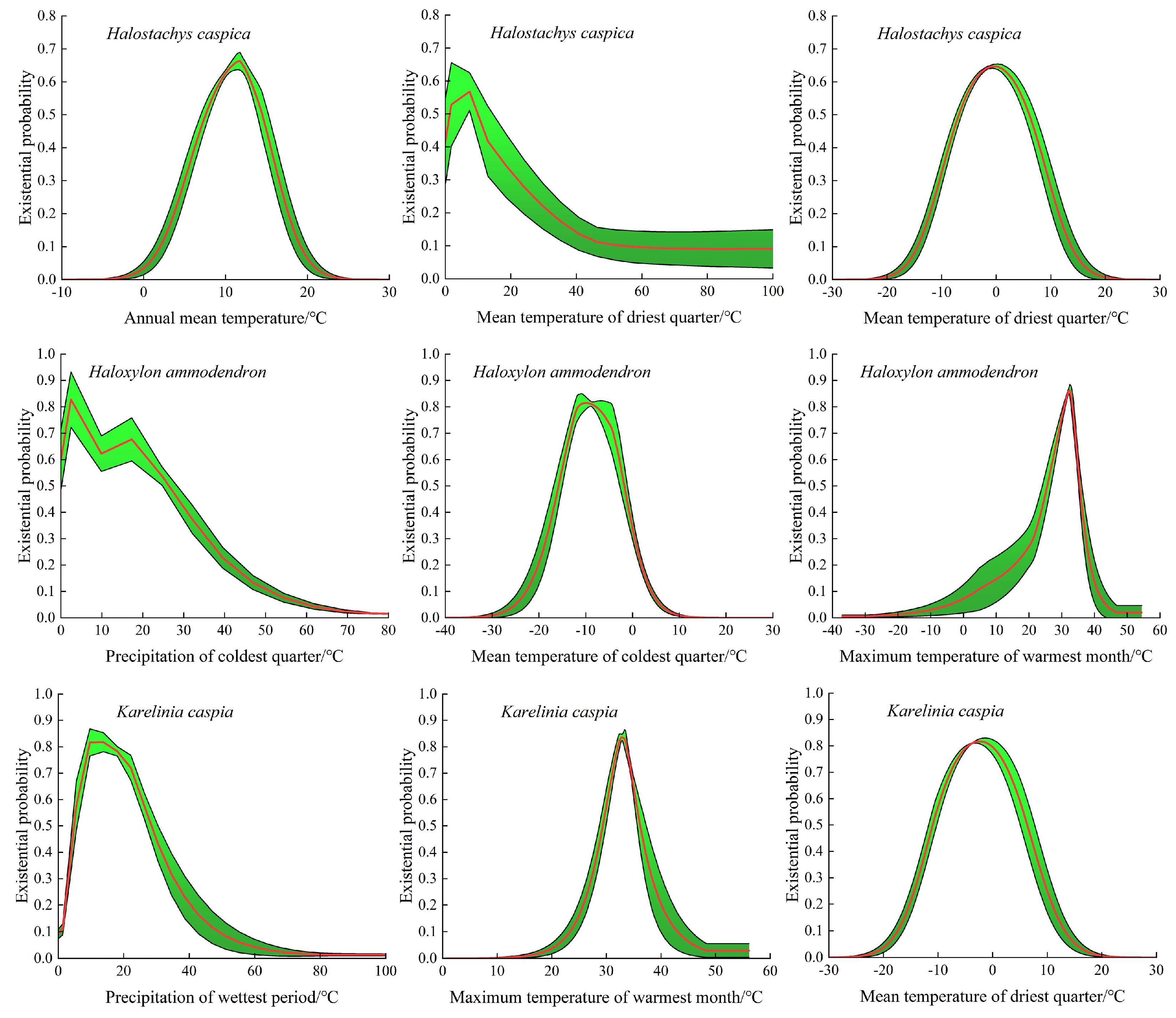
3.2. Analysis of Dominant Environmental Factors
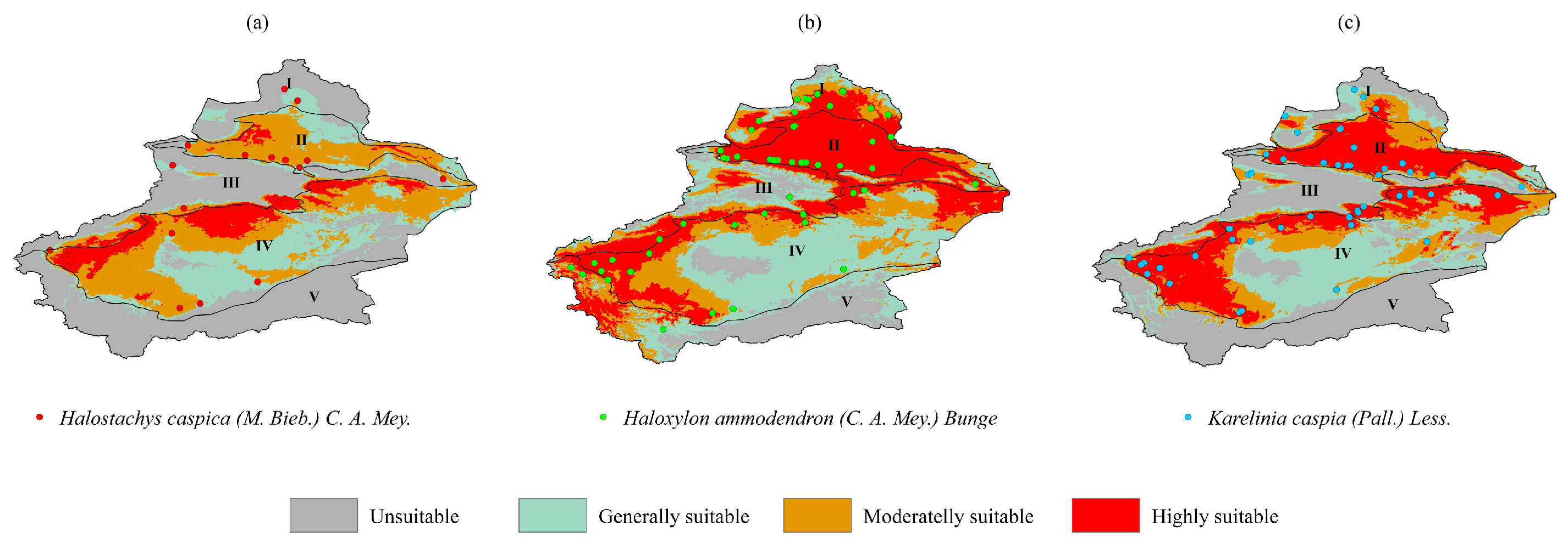
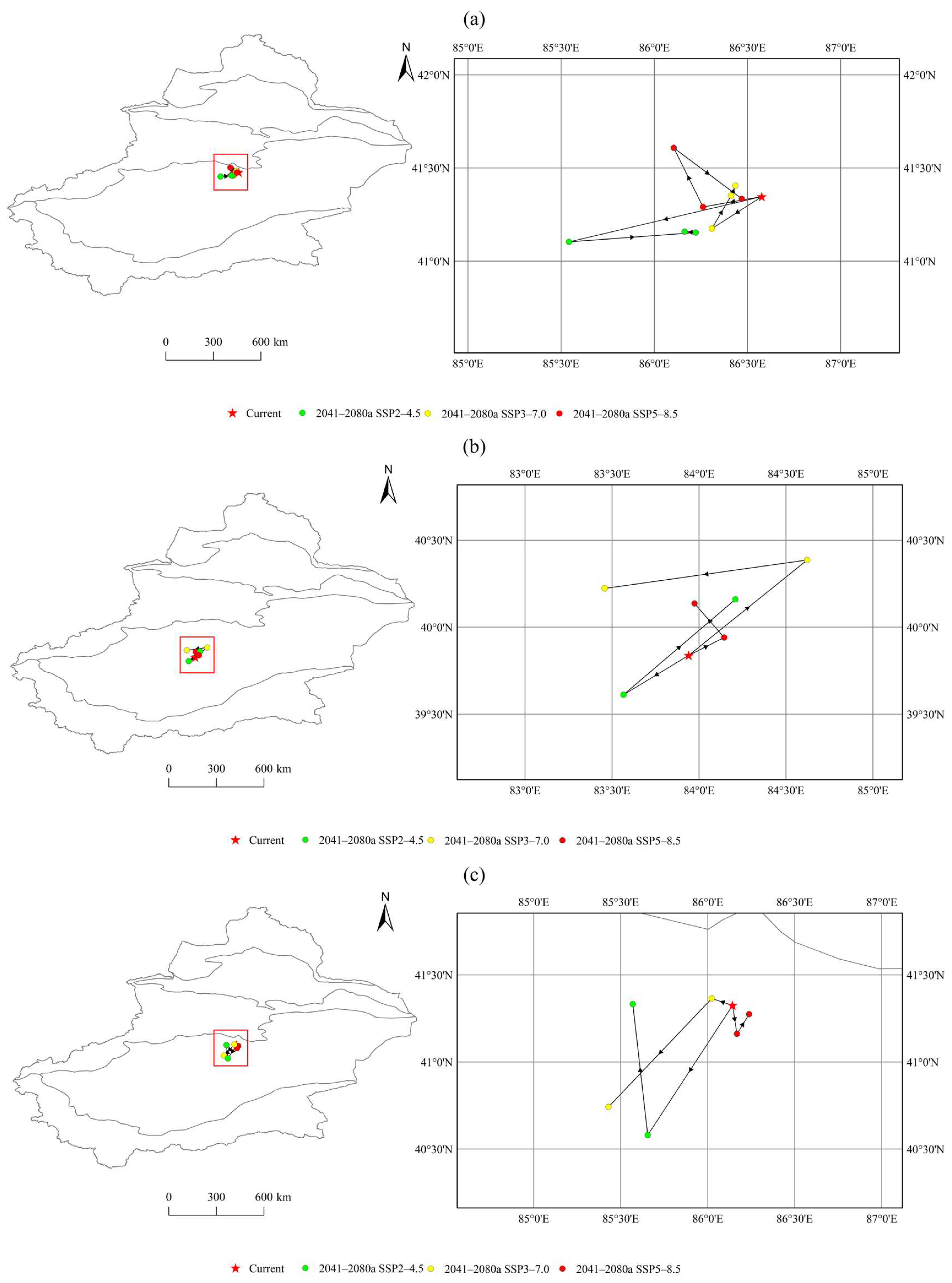
3.3. Spatial Pattern Changes in Three Species Under Current Climate Conditions
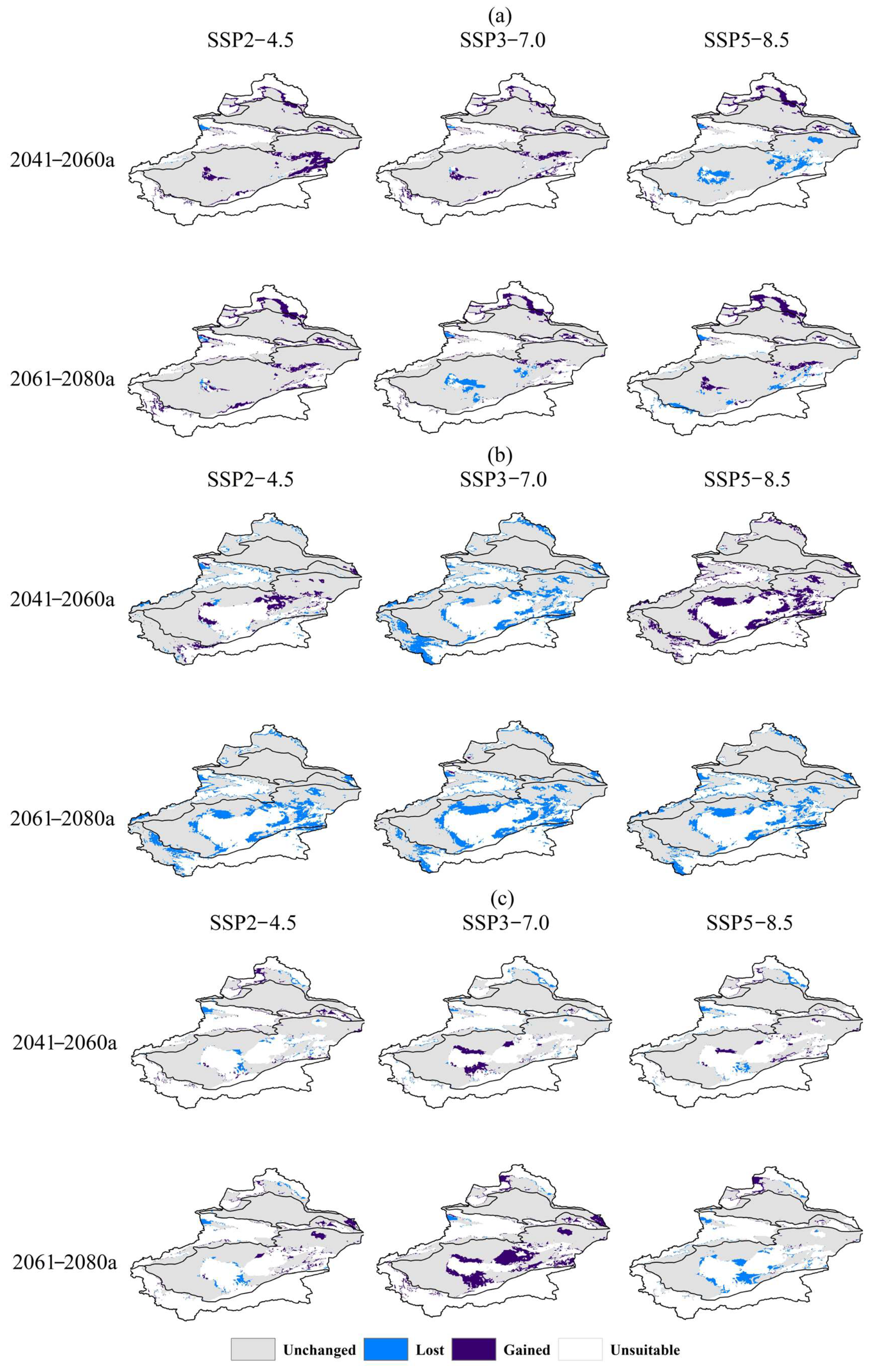
3.4. Spatial Pattern Changes in Three Species Under Different Scenarios
3.4.1. Future Spatial Pattern Changes in Halos. caspica
3.4.2. Future Spatial Pattern Changes in Halox. ammodendrum
3.4.3. Future Spatial Pattern Changes in K. caspia
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.; Ge, Q.; Geng, X.; Wang, Z.; Gao, L.; Bryan, B.A.; Chen, S.; Su, Y.; Cai, D.; Ye, J.; et al. Unintended Consequences of Combating Desertification in China. Nat. Commun. 2023, 14, 1139. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Liu, H.; Huang, J.; Gao, Z.; Wang, G.; Li, D.; Yu, H.; Chen, X. Accelerated Dryland Expansion Regulates Future Variability in Dryland Gross Primary Production. Nat. Commun. 2020, 11, 1665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yang, J.; Wang, W.; Ma, P.; Lu, G.; Liu, X.; Yu, H.; Fang, F. Climatic Warming and Humidification in the Arid Region of Northwest China: Multi-Scale Characteristics and Impacts on Ecological Vegetation. J. Meteorol. Res. 2021, 35, 113–127. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of Climate Change on the Future of Biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef]
- Bonan, G.B.; Doney, S.C. Climate, Ecosystems, and Planetary Futures: The Challenge to Predict Life in Earth System Models. Science 2018, 359, eaam8328. [Google Scholar] [CrossRef]
- Kaufmann, R.K.; Zhou, L.; Myneni, R.B.; Tucker, C.J.; Slayback, D.; Shabanov, N.V.; Pinzon, J. The Effect of Vegetation on Surface Temperature: A Statistical Analysis of NDVI and Climate Data. Geophys. Res. Lett. 2003, 30, 2003GL018251. [Google Scholar] [CrossRef]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India Lead in Greening of the World through Land-Use Management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef]
- Piao, S.; Wang, X.; Park, T.; Chen, C.; Lian, X.; He, Y.; Bjerke, J.W.; Chen, A.; Ciais, P.; Tømmervik, H.; et al. Characteristics, Drivers and Feedbacks of Global Greening. Nat. Rev. Earth Environ. 2020, 1, 14–27. [Google Scholar] [CrossRef]
- Lian, X.; Piao, S.; Chen, A.; Huntingford, C.; Fu, B.; Li, L.Z.X.; Huang, J.; Sheffield, J.; Berg, A.M.; Keenan, T.F.; et al. Multifaceted Characteristics of Dryland Aridity Changes in a Warming World. Nat. Rev. Earth Environ. 2021, 2, 232–250. [Google Scholar] [CrossRef]
- Yin, G.; Hu, Z.; Chen, X.; Tiyip, T. Vegetation Dynamics and Its Response to Climate Change in Central Asia. J. Arid Land 2016, 8, 375–388. [Google Scholar] [CrossRef]
- Warren, R.; Price, J.; Graham, E.; Forstenhaeusler, N.; VanDerWal, J. The Projected Effect on Insects, Vertebrates, and Plants of Limiting Global Warming to 1.5 °C Rather than 2 °C. Science 2018, 360, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Din, J.U.; Bosso, L.; Hameed, S.; Kabir, M.; Younas, M.; Nawaz, M.A. Expanding or Shrinking? Range Shifts in Wild Ungulates under Climate Change in Pamir-Karakoram Mountains, Pakistan. PLoS ONE 2022, 16, e0260031. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Wang, R.; Hou, K.; Wang, X.; Wu, W. Predicting the Current and Future Cultivation Regions of Carthamus Tinctorius L. Using MaxEnt Model under Climate Change in China. Glob. Ecol. Conserv. 2018, 16, e00477. [Google Scholar] [CrossRef]
- Paim, F.P.; Carretero-Pinzón, X.; Guzmán-Caro, D.C.; Alfaro, J.L.; Calouro, A.M. The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2021. [Google Scholar]
- Gaston, K.J. Species-Range-Size Distributions: Patterns, Mechanisms and Implications. Trends Ecol. Evol. 1996, 11, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Wei-Yao, K.; Xin-Hai, L.I.; Hong-Fei, Z.O.U. Optimizing MaxEnt Model in the Prediction of Species Distribution. Yingyong Shengtai Xuebao 2019, 30, 2116. [Google Scholar]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Hannah, L.; Roehrdanz, P.R.; Ikegami, M.; Shepard, A.V.; Shaw, M.R.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R.J. Climate Change, Wine, and Conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar] [CrossRef]
- Khanum, R.; Mumtaz, A.S.; Kumar, S. Predicting Impacts of Climate Change on Medicinal Asclepiads of Pakistan Using Maxent Modeling. Acta Oecol. 2013, 49, 23–31. [Google Scholar] [CrossRef]
- Hosseini, N.; Ghorbanpour, M.; Mostafavi, H. Habitat Potential Modelling and the Effect of Climate Change on the Current and Future Distribution of Three Thymus Species in Iran Using MaxEnt. Sci. Rep. 2024, 14, 3641. [Google Scholar] [CrossRef]
- Su, B.; Huang, J.; Mondal, S.K.; Zhai, J.; Wang, Y.; Wen, S.; Gao, M.; Lv, Y.; Jiang, S.; Jiang, T.; et al. Insight from CMIP6 SSP-RCP Scenarios for Future Drought Characteristics in China. Atmos. Res. 2021, 250, 105375. [Google Scholar] [CrossRef]
- Li, S.-Y.; Miao, L.-J.; Jiang, Z.-H.; Wang, G.-J.; Gnyawali, K.R.; Zhang, J.; Zhang, H.; Fang, K.; He, Y.; Li, C. Projected Drought Conditions in Northwest China with CMIP6 Models under Combined SSPs and RCPs for 2015–2099. Adv. Clim. Change Res. 2020, 11, 210–217. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, S.; He, Y. Projection of Summer Rainfall in the Yangtze River Basin in the Next 30 Years under Different Shared Socioeconomic Pathways. Chin. J. Atmos. Sci. 2023, 47, 1405–1420. [Google Scholar] [CrossRef]
- Wu, X.; Zheng, X.J.; Yin, X.W.; Yue, Y.M.; Li, Y. Seasonal Variation in the Groundwater Dependency of Two Dominant Woody Species in a Desert Region of Central Asia. Plant Soil 2019, 444, 39–55. [Google Scholar] [CrossRef]
- Xiao, F.; Liu, Q.; Qin, Y. Predicting the Potential Distribution of Haloxylon Ammodendron under Climate Change Scenarios Using Machine Learning of a Maximum Entropy Model. Biology 2024, 13, 3. [Google Scholar] [CrossRef]
- Zeng, Y.; Li, L.; Yang, R.; Yi, X.; Zhang, B. Contribution and Distribution of Inorganic Ions and Organic Compounds to the Osmotic Adjustment in Halostachys Caspica Response to Salt Stress. Sci. Rep. 2015, 5, 13639. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, H.; Wang, L.; Zeng, Y. Effect of Salt-Alkali Stress on Seed Germination of the Halophyte Halostachys Caspica. Sci. Rep. 2024, 14, 13199. [Google Scholar] [CrossRef]
- Lü, X.-P.; Gao, H.-J.; Zhang, L.; Wang, Y.-P.; Shao, K.-Z.; Zhao, Q.; Zhang, J.-L. Dynamic Responses of Haloxylon Ammodendron to Various Degrees of Simulated Drought Stress. Plant Physiol. Biochem. 2019, 139, 121–131. [Google Scholar] [CrossRef]
- Islam, W.; Zeng, F.; Mubarak Alwutayd, K.; Ali Khan, K. Beneath the Surface: Investigating Soil Microbial and Metazoa Communities at Various Depths in a Natural Desert Ecosystem Inhabited by Karelinia Caspia. Ecol. Indic. 2024, 159, 111745. [Google Scholar] [CrossRef]
- Du, H.; Zhou, C.; Tang, H.; Jin, X.; Chen, D.; Jiang, P.; Li, M. Correction to: Simulation and Estimation of Future Precipitation Changes in Arid Regions: A Case Study of Xinjiang, Northwest China. Clim. Change 2021, 167, 43. [Google Scholar] [CrossRef]
- Tian, Y.; Li, J.G.; Pan, L.P.; Zhao, Y. The Key Factors Affecting Haloxylon Ammodendron Germination and Survival at Very Early Stage. Acta Ecol. Sin. 2010, 30, 4898–4904. [Google Scholar]
- Hua, Y.C.; Zhong, D.J.; Mei, S.Q.; Ke, Z.; Yong, Z.Z.; Yan, T.C. Salt Island Effect of Halophytic Shrubs in Different Habitats and Its Ecological Implication. Acta Pedol. Sin. 2012, 49, 289–295. [Google Scholar]
- Chen, L.; Li, M.; Li, C.; Zheng, W.; Liu, R. Different Physiological Responses to Continuous Drought between Seedlings and Younger Individuals of Haloxylon Ammodendron. Plants 2023, 12, 3683. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Jun, C.; Sai, L.; Rong, X.U.; Chang-qing, X.U.; Tong-ning, L.I.U. Influence of Dehydration and Diurnal Variation on Characteristics of Chlorophyll Fluorescence of Leaves in Haloxylon Ammodendron and H. Persicum. Yingyong Shengtai Xuebao 2015, 26, 2321. [Google Scholar]
- Yan, K.; Ablimit, M.; Liu, S.; Liu, Z.; Wang, Y. A Novel Metallothionein Gene HcMT from Halophyte Shrub Halostachys Caspica Respond to Cadmium and Sodium Stress. Plant Physiol. Biochem. 2023, 201, 107763. [Google Scholar] [CrossRef]
- Mielke, K.P.; Claassen, T.; Busana, M.; Heskes, T.; Huijbregts, M.A.J.; Koffijberg, K.; Schipper, A.M. Disentangling Drivers of Spatial Autocorrelation in Species Distribution Models. Ecography 2020, 43, 1741–1751. [Google Scholar] [CrossRef]
- Wan, G.-Z.; Wang, L.; Jin, L.; Chen, J. Evaluation of Environmental Factors Affecting the Quality of Codonopsis Pilosula Based on Chromatographic Fingerprint and MaxEnt Model. Ind. Crops Prod. 2021, 170, 113783. [Google Scholar] [CrossRef]
- Wang, T.; Tu, X.; Singh, V.P.; Chen, X.; Lin, K. Global Data Assessment and Analysis of Drought Characteristics Based on CMIP6. J. Hydrol. 2021, 596, 126091. [Google Scholar] [CrossRef]
- Su, W.; Wu, D.; Wu, H.; Zhang, M.; Zhang, R. Upscaling Method for Corn Canopy LAI Using MaxEnt Model. Trans. Chin. Soc. Agric. Eng. 2016, 32, 165–172. [Google Scholar]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists: Statistical Explanation of MaxEnt. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Dreiseitl, S. Towards More Efficient Multiclass AUC Computations. In Proceedings of the 31st European Modeling and Simulation Symposium, EMSS 2019, Lisbon, Portugal, 18–20 September 2019; pp. 327–331. [Google Scholar] [CrossRef]
- Niels, J.; Mark, R. Assessing the Performance and Accuracy of Invasive Plant Habitat Suitability Models in Detecting New Observations in Wisconsin. Invasive Plant Sci. Manag. 2021, 14, 214–222. [Google Scholar]
- Zhao, Y.; Deng, X.; Xiang, W.; Chen, L.; Ouyang, S. Predicting Potential Suitable Habitats of Chinese Fir under Current and Future Climatic Scenarios Based on Maxent Model. Ecol. Inform. 2021, 64, 101393. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting Thresholds of Occurrence in the Prediction of Species Distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Zhang, K.; Yao, L.; Meng, J.; Tao, J. Maxent Modeling for Predicting the Potential Geographical Distribution of Two Peony Species under Climate Change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- He, Y.; Huang, W.; Zhao, X.; Lv, P.; Wang, H. Review on the Impact of Climate Change on Plant Diversity. J. Desert Res. 2021, 41, 59. [Google Scholar]
- Wu, J.; Lv, J.; Ai, L. The Impacts of Climate Change on the Biodiversity: Vulnerability and Adaptation. Ecol. Environ. 2009, 18, 693. [Google Scholar]
- Zhu, Y.; Jia, Z.; Wang, G. Response of Seed Germination and Seedling Emergence of Haloxylon Ammodendron to Climate Change in Desert Ecosystem. Res. Gate 2022. [Google Scholar] [CrossRef]
- Guo, N.; Chen, F.; He, C.; Wang, T.; Long, A.; Xu, X. Multi-Year Average Water Vapor Characteristics and Potential Sources and Transport Pathways of Intense Water Vapor during Extreme Precipitation Events in the Ili River Valley, China. J. Hydrol. Reg. Stud. 2025, 58, 102278. [Google Scholar] [CrossRef]
- Wu, X.; Luo, M.; Meng, F.; Sa, C.; Yin, C.; Bao, Y. New Characteristics of Spatio-Temporal Evolution of Extreme Climate Events in Xinjiang under the Background of Warm and Humid Climate. Arid. Zone Res. 2022, 39, 1695–1705. [Google Scholar]
- Xie, P.; Zhang, Y.; Qiao, F. Spatial and Temporal Characteristics of Extreme Precipitation in Xinjiang and Its Response to ENSO. J. Meteorol. Res. Appl. 2019, 40, 5–12. [Google Scholar]
- Trew, B.T.; Maclean, I.M.D. Vulnerability of Global Biodiversity Hotspots to Climate Change. Glob. Ecol. Biogeogr. 2021, 30, 768–783. [Google Scholar] [CrossRef]
- Adapting to the Impacts of Climate Change|The National Academies Press. Available online: https://nap.nationalacademies.org/catalog/12783/adapting-to-the-impacts-of-climate-change (accessed on 26 May 2025).
- Zhang, H.-X.; Zhang, M.-L. Insight into Distribution Patterns and Conservation Planning in Relation to Woody Species Diversity in Xinjiang, Arid Northwestern China. Biol. Conserv. 2014, 177, 165–173. [Google Scholar] [CrossRef]
- McDowell, N.G.; Anderson-Teixeira, K.; Biederman, J.A.; Breshears, D.D.; Fang, Y.; Fernández-de-Uña, L.; Graham, E.B.; Mackay, D.S.; McDonnell, J.J.; Moore, G.W.; et al. Ecohydrological Decoupling under Changing Disturbances and Climate. One Earth 2023, 6, 251–266. [Google Scholar] [CrossRef]
- Yu, H.; Bian, Z.; Mu, S.; Yuan, J.; Chen, F. Effects of Climate Change on Land Cover Change and Vegetation Dynamics in Xinjiang, China. Int. J. Environ. Res. Public Health 2020, 17, 4865. [Google Scholar] [CrossRef]
- Carrillo, J.; Hernández-Barrera, S.; Expósito, F.J.; Díaz, J.P.; González, A.; Pérez, J.C. The Uneven Impact of Climate Change on Drought with Elevation in the Canary Islands. npj Clim. Atmos. Sci. 2023, 6, 31. [Google Scholar] [CrossRef]
- Jiang, K.; Teuling, A.J.; Chen, X.; Huang, N.; Wang, J.; Zhang, Z.; Gao, R.; Men, J.; Zhang, Z.; Wu, Y.; et al. Global Land Degradation Hotspots Based on Multiple Methods and Indicators. Ecol. Indic. 2024, 158, 111462. [Google Scholar] [CrossRef]
- Wang, J.; Peng, J.; Li, H.; Yin, C.; Liu, W.; Wang, T.; Zhang, H. Soil Salinity Mapping Using Machine Learning Algorithms with the Sentinel-2 MSI in Arid Areas, China. Remote Sens. 2021, 13, 305. [Google Scholar] [CrossRef]
- Wang, E.; Huang, T.; Liu, Z.; Bao, L.; Guo, B.; Yu, Z.; Feng, Z.; Luo, H.; Ou, G. Improving Forest Above-Ground Biomass Estimation Accuracy Using Multi-Source Remote Sensing and Optimized Least Absolute Shrinkage and Selection Operator Variable Selection Method. Remote Sens. 2024, 16, 4497. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Names | Classification (Family) | Habitat Characteristics | Species Growth Habits | Distribution Range in Central Asia |
|---|---|---|---|---|
| Halostachys capsica (M. Bieb.) C. A. Mey | Amaranthaceae | Saline–alkali flats, river valleys, salt lake shores, and saline–alkali soils. | Shrubs, 50–200 cm tall. | Distributed in Afghanistan, Russia, Mongolia, Iran, and China; in China, it is mainly found in Xinjiang and northern Gansu. |
| Haloxylon ammodendron (C. A. Mey.) Bunge | Chenopodiaceae | Dunes, saline–alkali deserts, riverside sandy areas, sandy soils and saline–alkali soils. | Small trees, 1–9 m tall, with a ground diameter of up to 50 cm on their trunks. | Distributed in Central Asia, Xinjiang, western Gansu, Inner Mongolia, and other regions; suitable habitats are found in areas such as the Tarim Basin, the northern slopes of the Tianshan Mountains, and the western edge of the Taklamakan Desert in Central Asia. |
| Karelinia caspia (Pall.) Less | Asteraceae | Halophytic meadows and salinized lowlands in desert zones, along farmland edges, with soils ranging from slightly to moderately salinized or severely salinized. | Perennial herb, 50–100 cm tall, sometimes up to 150 cm. | Distributed in Central Asia, Mongolia, Iran, Turkey, and regions such as Inner Mongolia, Ningxia, and Gansu in China; it is also found in countries like Kazakhstan and Uzbekistan in Central Asia. |
| Numbering | Environmental Variables | Unit |
|---|---|---|
| Bio1 | Annual mean temperature | °C |
| Bio2 | Mean diurnal range | °C |
| Bio3 | Isothermality | |
| Bio4 | Standard deviation of seasonal variation in temperature | |
| Bio5 | Maximum temperature of warmest month | |
| Bio6 | Minimum temperature of coldest month | °C |
| Bio7 | Temperature annual range | °C |
| Bio8 | Mean temperature of wettest quarter | °C |
| Bio9 | Mean temperature of driest quarter | °C |
| Bio10 | Mean temperature of warmest quarter | °C |
| Bio11 | Mean temperature of coldest quarter | °C |
| Bio12 | Annual precipitation | mm |
| Bio13 | Precipitation of wettest period | mm |
| Bio14 | Precipitation of driest period | mm |
| Bio15 | Precipitation of wettest quarter | |
| Bio16 | Precipitation of driest quarter | mm |
| Bio17 | Precipitation seasonality | mm |
| Bio18 | Precipitation of warmest quarter | mm |
| Bio19 | Precipitation of coldest quarter | mm |
| Numbering | Environmental Variables | Percent Contribution | Unit |
|---|---|---|---|
| Bio1 | Annual mean temperature | 30.5 | °C |
| Bio2 | Mean diurnal range | 2.6 | °C |
| Bio3 | Isothermality | 0.7 | |
| Bio4 | Standard deviation of seasonal variation in temperature | 14.6 | |
| Bio16 | Precipitation of driest quarter | 17 | mm |
| Bio19 | Precipitation of coldest quarter | 13.9 | mm |
| Bio16 | Precipitation of driest quarter | 1.7 | mm |
| Bio19 | Precipitation of coldest quarter | 19 | mm |
| Period–Climate Scenario | Area Change/104 km2 | |||||
|---|---|---|---|---|---|---|
| Halostachys capsica | Haloxylon ammodendron | Karelinia caspia | ||||
| Increase | Lost | Increase | Lost | Increase | Lost | |
| 2041–2060 SSP-2.45 | 10.38 | 0.47 | 6.74 | 4.15 | 3.47 | 2.5 |
| 2041–2060 SSP-3.70 | 8.44 | 0.3 | 0.07 | 20.26 | 4.07 | 2.23 |
| 2041–2060 SSP-5.85 | 5.33 | 7.03 | 18.93 | 0.22 | 3.27 | 2.57 |
| 2061–2080 SSP-2.45 | 10 | 0.46 | 0.06 | 12.2 | 2.63 | 1.31 |
| 2061–2080 SSP-3.70 | 6.73 | 3.48 | 0.23 | 13.28 | 9.15 | 0.44 |
| 2061–2080 SSP-5.85 | 7.92 | 3.22 | 0.07 | 10.75 | 1.05 | 3.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, H.; Tao, H.; Zhang, Z. Projected Spatial Distribution Patterns of Three Dominant Desert Plants in Xinjiang of Northwest China. Forests 2025, 16, 1031. https://doi.org/10.3390/f16061031
Cao H, Tao H, Zhang Z. Projected Spatial Distribution Patterns of Three Dominant Desert Plants in Xinjiang of Northwest China. Forests. 2025; 16(6):1031. https://doi.org/10.3390/f16061031
Chicago/Turabian StyleCao, Hanyu, Hui Tao, and Zengxin Zhang. 2025. "Projected Spatial Distribution Patterns of Three Dominant Desert Plants in Xinjiang of Northwest China" Forests 16, no. 6: 1031. https://doi.org/10.3390/f16061031
APA StyleCao, H., Tao, H., & Zhang, Z. (2025). Projected Spatial Distribution Patterns of Three Dominant Desert Plants in Xinjiang of Northwest China. Forests, 16(6), 1031. https://doi.org/10.3390/f16061031