
Physiological and Transcriptomic Insights into Lead Uptake and Tolerance in Moso Bamboo (Phyllostachys edulis) Highlight Its Strong Lead Tolerance Capacity
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Growth Conditions
2.2. Growth Phenotype of Moso Bamboo Under Pb Stress
2.3. Determination of Element Concentrations of Moso Bamboo Under Pb Stress
2.4. Pb Uptake Kinetics
2.5. Xylem Sap Collection and Pb Concentration Determination
2.6. Cellular Distribution of Pb in Moso Bamboo Root
2.7. Root Cellular Morphology After Pb Stress
2.8. Pb Distribution and Accumulation in Leaves
2.9. RNA Extraction, Transcriptome Sequencing, and qRT-PCR Analysis
2.10. Statistical Analysis
3. Results
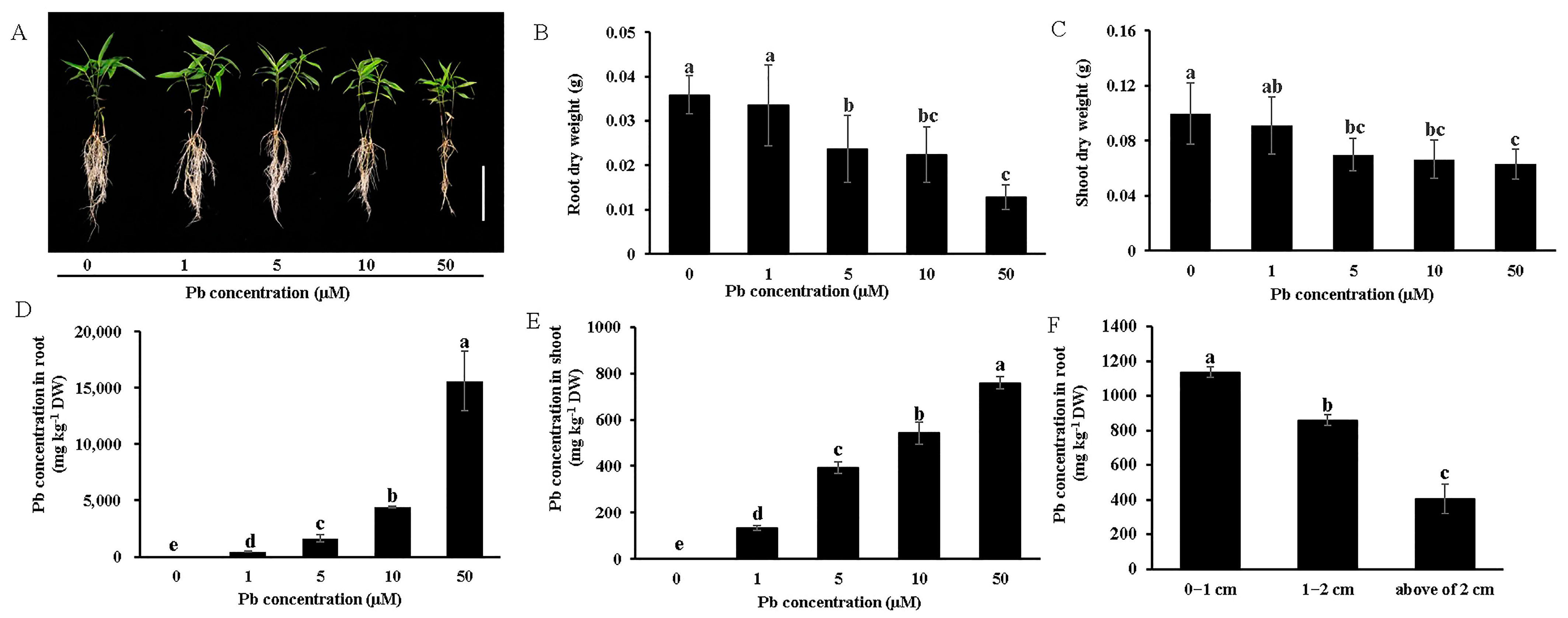
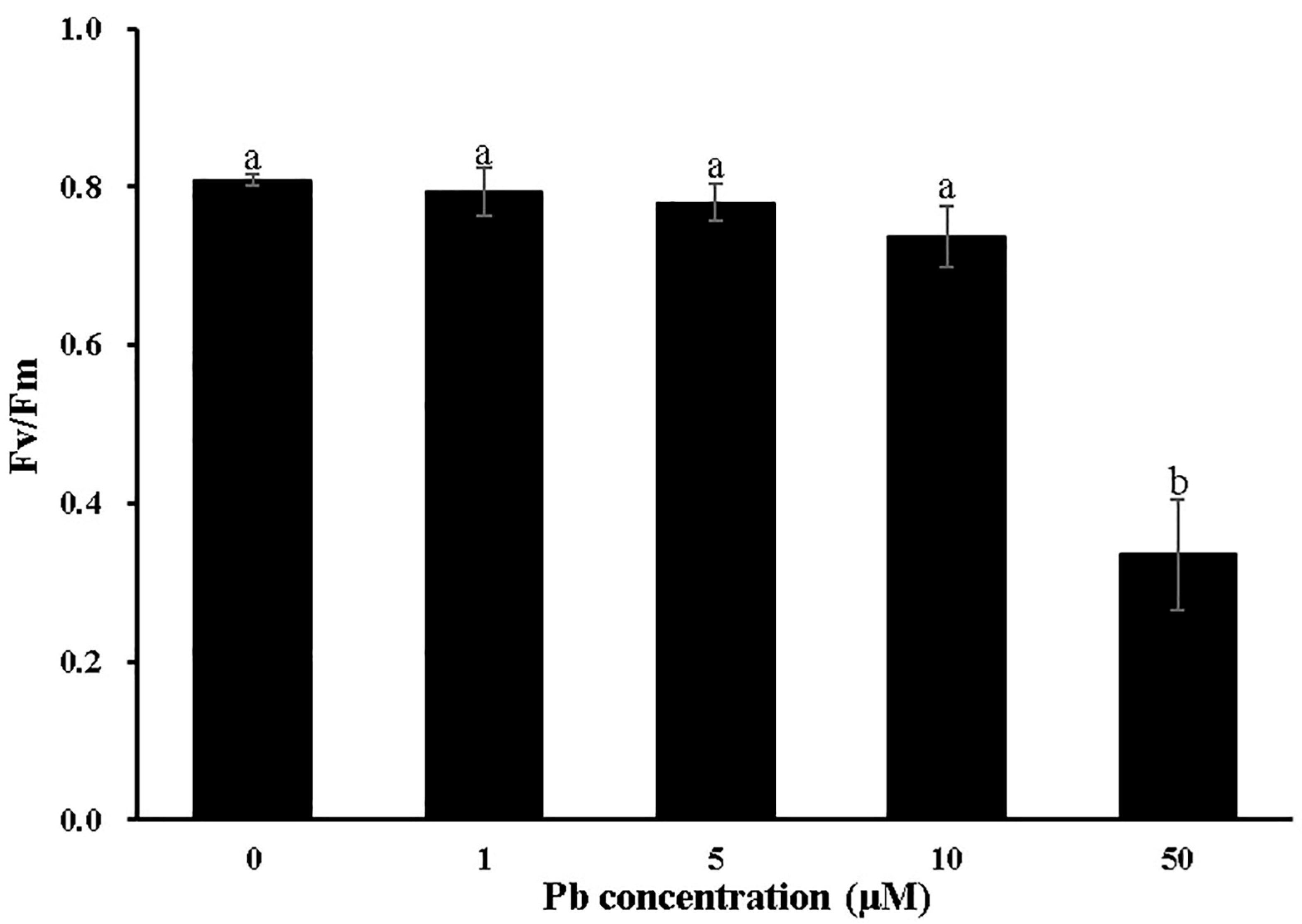
3.1. Effects of Pb Stress on the Growth of Moso Bamboo
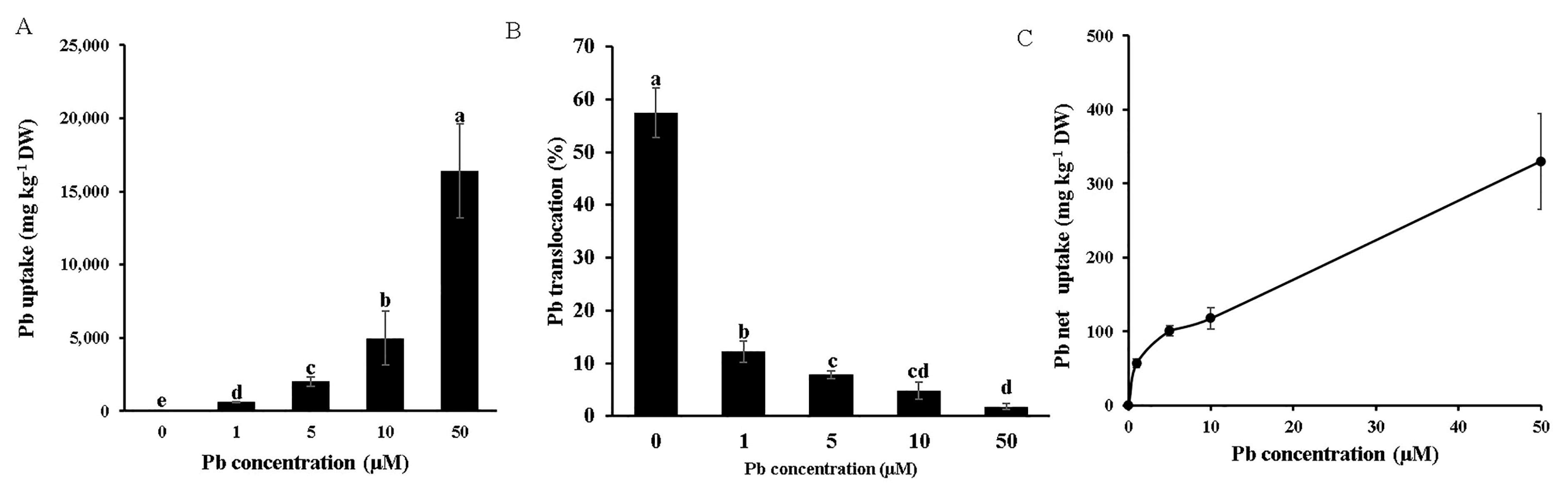
3.2. Pb Uptake, Translocation, and Distribution
3.3. Xylem Loading of Pb
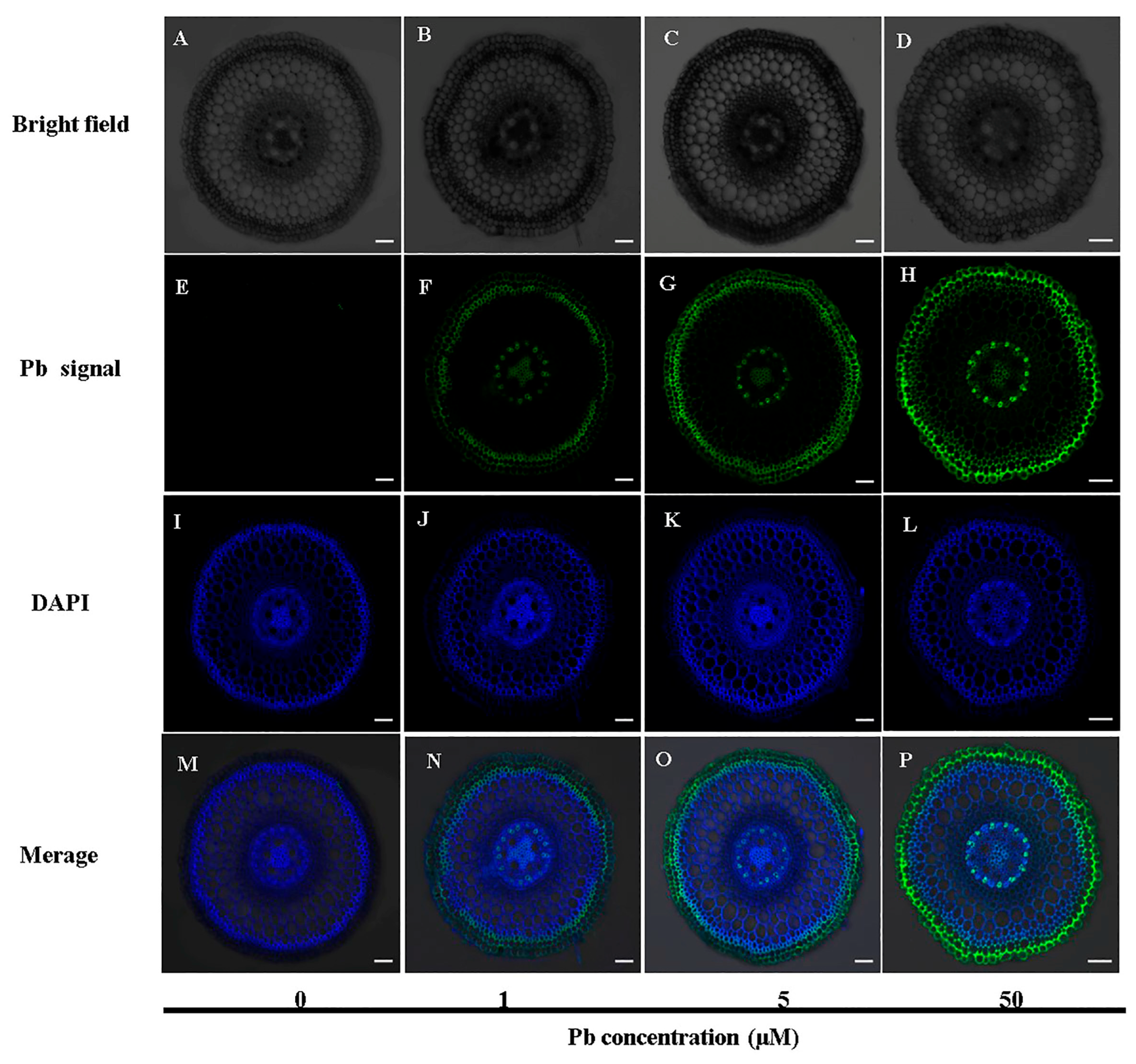
3.4. Cellular Localization of Pb in Roots
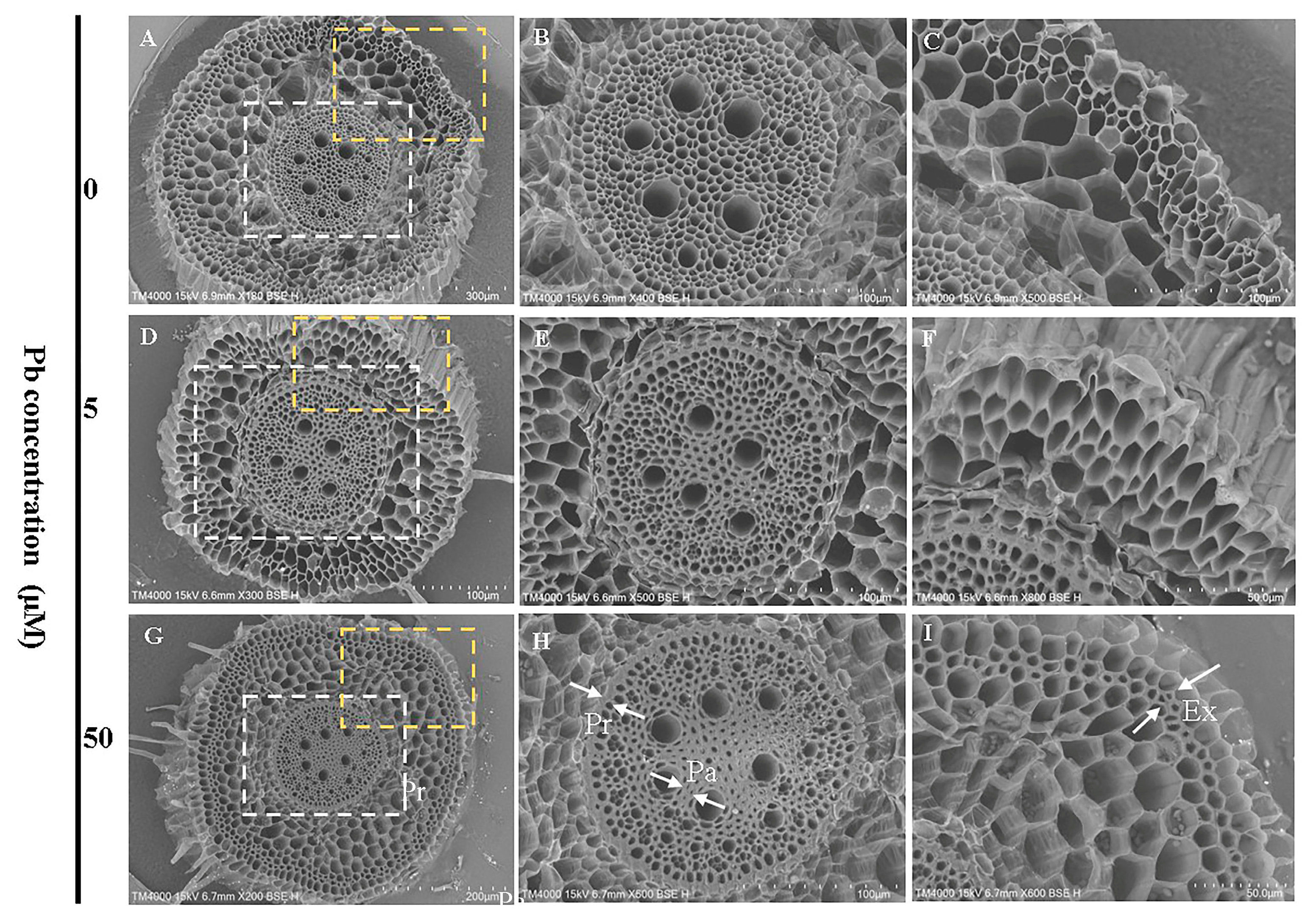
3.5. Effects of Pb Stress on Root Cellular Morphology
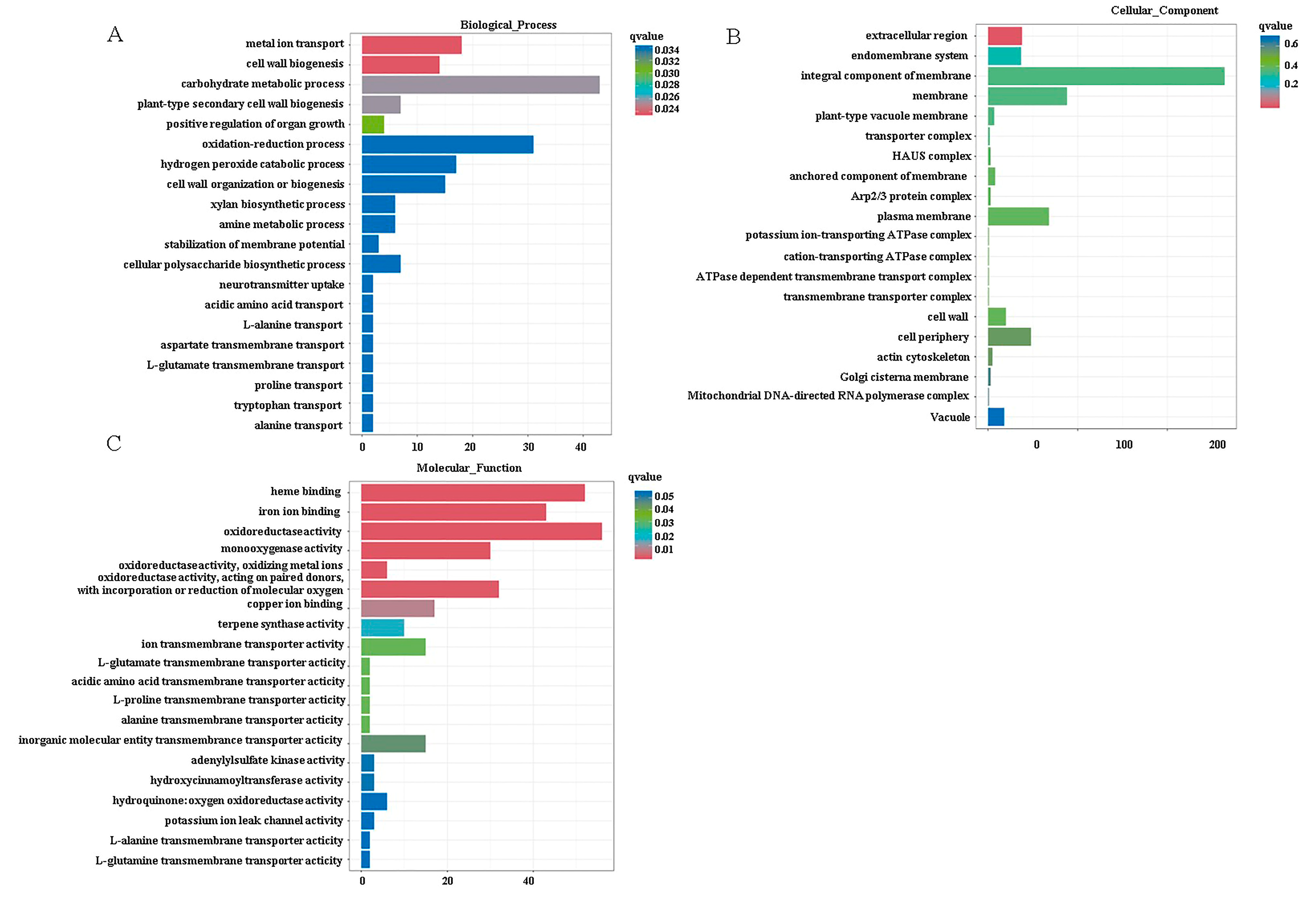
3.6. Sequencing Quality Overview and DEG Analysis
4. Discussion
4.1. Moso Bamboo Exhibits High Pb Tolerance and Accumulation Through Physiological and Anatomical Adaptations
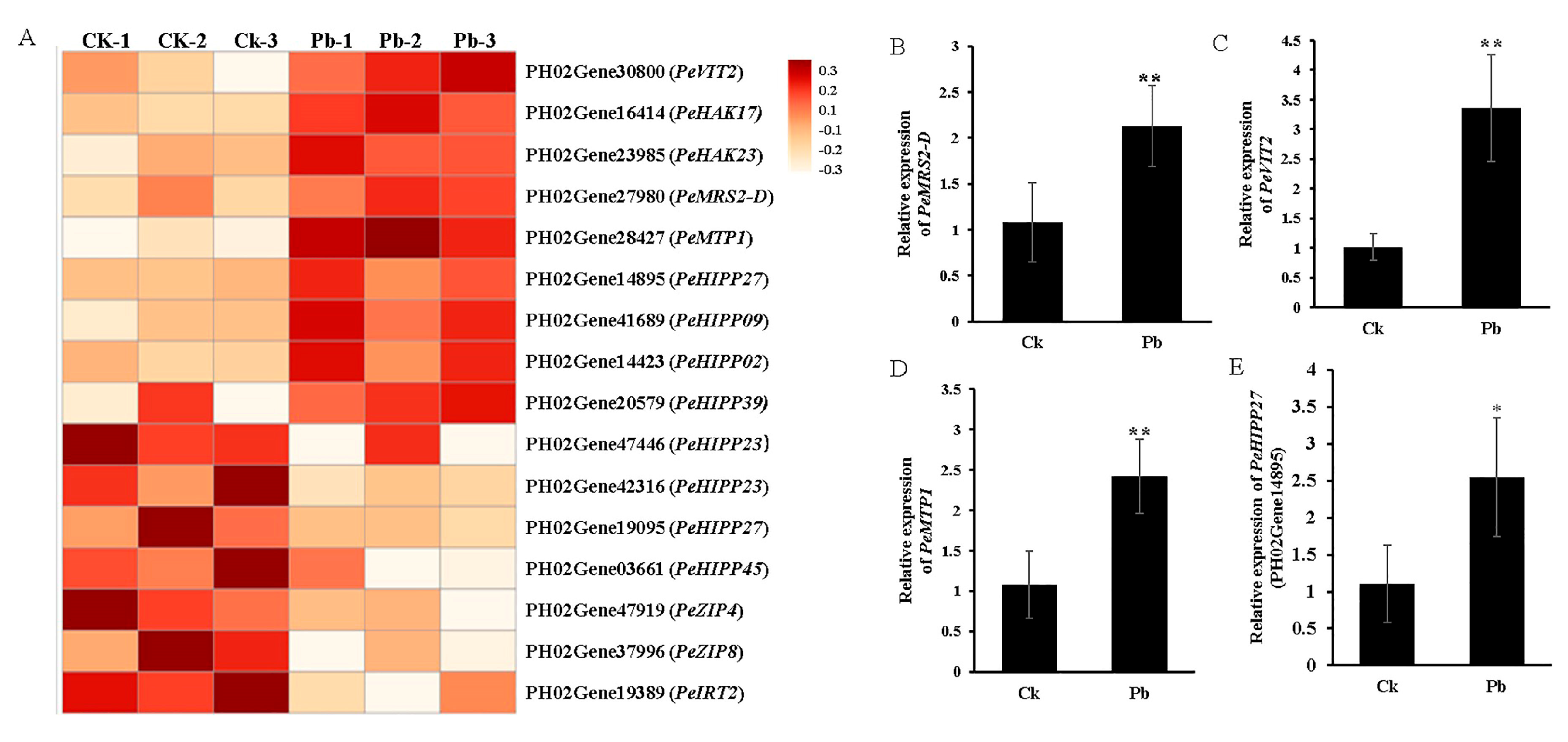
4.2. Transcriptomic Regulation of Metal Cation Transporters Under Pb Stress in Moso Bamboo
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Küpper, H. Lead toxicity in plants. Met. Ions Life Sci. 2017, 17. [Google Scholar] [CrossRef]
- Zhao, F.J.; Tang, Z.; Song, J.J.; Huang, X.Y.; Wang, P. Toxic metals and metalloids: Uptake, transport, detoxification, phytoremediation, and crop improvement for safer food. Mol. Plant. 2022, 15, 27–44. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.K.; Huang, H.G.; Corpas, F.J. Lead tolerance in plants: Strategies for phytoremediation. Environ. Sci. Pollut. Res. Int. 2013, 4, 2150–2161. [Google Scholar] [CrossRef]
- Pourrut, B.; Shahid, M.; Dumat, C.; Winterton, P.; Pinelli, E. Lead uptake, toxicity, and detoxification in plants. Rev. Environ. Contam. Toxicol. 2011, 213, 113–136. [Google Scholar] [PubMed]
- Zhang, H.; Sun, X.; Hwarari, D.; Du, X.; Wang, Y.; Xu, H.; Lv, S.; Wang, T.; Yang, L.; Hou, D. Oxidative stress response and metal transport in roots of Macleaya cordata exposed to lead and zinc. Plants 2023, 12, 516. [Google Scholar] [CrossRef]
- Kaur, G.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Adaptations to oxidative stress in Zea mays root under short-term Pb2+ exposure. Biologia 2015, 70, 190–197. [Google Scholar] [CrossRef]
- Liu, D.; Islam, E.; Li, T.Q.; Yang, X.; Jin, X.F.; Mahmood, Q. Comparison of synthetic chelators and low molecular weight organic acids in enhancing phytoextraction of heavy metals by two ecotypes of Sedum alfredii Hance. J. Hazard. Mater. 2008, 153, 114–122. [Google Scholar] [CrossRef]
- Shahid, M.; Pourrut, B.; Dumat, C.; Nadeem, M.; Aslam, M.; Pinelli, E. Heavy-metal-induced reactive oxygen species: Phytotoxicity and physicochemical changes in plants. Rev. Environ. Contam. Toxicol. 2014, 232, 1–44. [Google Scholar]
- Gupta, D.K.; Nicoloso, F.T.; Schetinger, M.R.; Rossato, L.V.; Pereira, L.B.; Castro, G.Y.; Srivastava, S.; Tripathi, R.D. Antioxidant defense mechanism in hydroponically grown Zea mays seedlings under moderate lead stress. J. Hazard. Mater. 2009, 172, 479–484. [Google Scholar] [CrossRef]
- Li, X.; Bu, N.; Li, Y.; Ma, L.; Xin, S.; Zhang, L. Growth, photosynthesis and antioxidant responses of endophyte infected and non-infected rice under lead stress conditions. J. Hazard. Mater. 2012, 213, 55–61. [Google Scholar] [CrossRef]
- Shao, J.F.; Che, J.; Yamaji, N.; Shen, R.F.; Ma, J.F. Silicon reduces cadmium accumulation by suppressing expression of transporter genes involved in cadmium uptake and translocation in rice. J. Exp. Bot 2017, 68, 5641–5651. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, H.Y.; Zhang, Y.; Huang, J.; Chen, Z.; Shen, R.F.; Zhu, X.F. Auxin is involved in cadmium accumulation in rice through controlling nitric oxide production and the ability of cell walls to bind cadmium. Sci. Total Environ. 2023, 904, 166644. [Google Scholar] [CrossRef] [PubMed]
- Morel, M.; Crouzet, J.; Gravot, A.; Auroy, P.; Leonhardt, N.; Vavasseur, A.; Richaud, P. AtHMA3, a P1B-ATPase allowing Cd/Zn/Co/Pb vacuolar storage in Arabidopsis. Plant Physiol. 2009, 149, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Krzesłowska, M. The cell wall in plant cell response to trace metals: Polysaccharide remodeling and its role in defense strategy. Acta Physiol. Plant. 2011, 33, 35–51. [Google Scholar] [CrossRef]
- Wang, F.; Zhao, W.; Chen, Y. Spatial Variations of Soil Heavy Metal Potential Ecological Risks in Typical Moso Bamboo Forests of Southeast China. Bull. Environ. Contam. Toxicol. 2019, 102, 224–230. [Google Scholar] [CrossRef]
- Zhong, B.; Chen, J.; Shafi, M.; Guo, J.; Wang, Y.; Wu, J.; Ye, Z.; He, L.; Liu, D. Effect of lead (Pb) on antioxidation system and accumulation ability of Moso bamboo (Phyllostachys pubescens). Ecotoxicol. Environ. Saf. 2017, 138, 71–77. [Google Scholar] [CrossRef]
- Liu, D.; Li, S.; Islam, E.; Chen, J.R.; Wu, J.S.; Ye, Z.Q.; Peng, D.L.; Yan, W.B.; Lu, K.P. Lead accumulation and tolerance of Moso bamboo (Phyllostachys pubescens) seedlings: Applications of phytoremediation. J. Zhejiang Univ. Sci. B 2015, 16, 123–130. [Google Scholar] [CrossRef]
- Li, S.; Chen, J.; Islam, E.; Wang, Y.; Wu, J.; Ye, Z.; Yan, W.; Peng, D.; Liu, D. Cadmium-induced oxidative stress, response of antioxidants and detection of intracellular cadmium in organs of moso bamboo (Phyllostachys pubescens) seedlings. Chemosphere 2016, 153, 107–114. [Google Scholar] [CrossRef]
- Gong, X.X.; Yang, F.; Pan, X.Y.; Shao, J.F. Accumulation of silicon in shoot is required for reducing lead uptake in rice. Crop J. 2022, 11, 1261–1271. [Google Scholar] [CrossRef]
- Li, X.; Sun, H.F.; Fan, J.H.; Li, Y.Y.; Ma, L.J.; Wang, L.L.; Li, X.M. Transcriptome modulation by endophyte drives rice seedlings response to Pb stress. Ecotoxicol. Environ. Saf. 2023, 254, 114740. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y.; Wei, X.; You, J.; Wang, W.; Lu, J.; Shi, R. Comparative antioxidative responses and proline metabolism in two wheat cultivars under short term lead stress. Ecotoxicol. Environ. Saf. 2011, 74, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.K.; Huang, H.G.; Yang, X.E.; Razafindrabe, B.H.; Inouhe, M. The detoxification of lead in Sedum alfredii H. is not related to phytochelatins but the glutathione. J. Hazard. Mater. 2010, 177, 437–444. [Google Scholar] [CrossRef]
- Yang, F.; Chang, Y.Z.; Zheng, Y.T.; Pan, X.Y.; Ji, H.B.; Shao, J.F. Physiological and transcriptomic characterization of cadmium toxicity in Moso bamboo (Phyllostachys edulis), a non-timber forest species. Tree Physiol. 2023, 43, 1250–1264. [Google Scholar] [CrossRef]
- Gao, P.P.; Liang, H.; Dong, Y.; Xue, P.Y.; Zhao, Q.L.; Yan, J.S.; Ma, W.; Zhao, J.J.; Liu, W.J. Transcriptomic mechanisms of reduced PM2.5-Pb retention in the leaves of the low-Pb-accumulation genotype of Chinese cabbage. J. Hazard. Mater. 2023, 444, 130385. [Google Scholar] [CrossRef] [PubMed]
- Ur Rahman, S.; Qin, A.; Zain, M.; Mushtaq, Z.; Mehmood, F.; Riaz, L.; Naveed, S.; Ansari, M.J.; Saeed, M.; Ahmad, I.; et al. Pb uptake, accumulation, and translocation in plants: Plant physiological, biochemical, and molecular response: A review. Heliyon 2024, 10, e27724. [Google Scholar] [CrossRef]
- Wang, Z.; Liang, Y.; Liu, H. Editorial: Phytoremediation of heavy metal contaminated soil: Technology, mechanism, and implementation. Front. Plant Sci. 2024, 14, 1347564. [Google Scholar] [CrossRef]
- Uraguchi, S.; Mori, S.; Kuramata, M.; Kawasaki, A.; Arao, T.; Ishikawa, S. Root-to-shoot Cd translocation via the xylem is the major process determining shoot and grain cadmium accumulation in rice. J. Exp. Bot. 2009, 60, 2677–2688. [Google Scholar] [CrossRef]
- Chang, J.D.; Gao, W.; Wang, P.; Zhao, F.J. OsNRAMP5 is a major transporter for lead uptake in rice. Environ. Sci. Technol. 2022, 56, 17481–17490. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.Y.; Belloeil, C.; Ianna, M.L.; Shin, R. Arabidopsis CNGC family members contribute to heavy metal ion uptake in plants. Int. J. Mol. Sci. 2019, 20, 413. [Google Scholar] [CrossRef]
- De Abreu-Neto, J.B.; Turchetto-Zolet, A.C.; De Oliveira, L.F.; Zanettini, M.H.; Margis-Pinheiro, M. Heavy metal-associated isoprenylated plant protein (HIPP): Characterization of a family of proteins exclusive to plants. FEBS J. 2013, 280, 1604–1616. [Google Scholar] [CrossRef]
- Wei, Y.; Peng, X.; Wang, X.; Wang, C. The heavy metal-associated isoprenylated plant protein (HIPP) gene family plays a crucial role in cadmium resistance and accumulation in the tea plant (Camellia sinensis L.). Ecotoxicol. Environ. Saf. 2023, 260, 115077. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, Y.H.; Yi, H.Y.; Gong, J.M. Vacuolar membrane transporters OsVIT1 and OsVIT2 modulate iron translocation between flag leaves and seeds in rice. Plant J. 2012, 72, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Yang, S.; Liu, B.; Zhang, M.; Wu, K. Molecular characterization of a rice metal tolerance protein, OsMTP1. Plant Cell Rep. 2012, 31, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Menguer, P.K.; Farthing, E.; Peaston, K.A.; Ricachenevsky, F.K.; Fett, J.P.; Williams, L.E. Functional analysis of the rice vacuolar zinc transporter OsMTP1. J. Exp. Bot. 2013, 64, 2871–2883. [Google Scholar] [CrossRef]
- Amini, S.; Arsova, B.; Hanikenne, M. The molecular basis of zinc homeostasis in cereals. Plant Cell Environ. 2022, 455, 1339–1361. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; Xu, R.; Zhu, C.; Ji, H.; Shao, J.F.; Huang, K. Physiological and Transcriptomic Insights into Lead Uptake and Tolerance in Moso Bamboo (Phyllostachys edulis) Highlight Its Strong Lead Tolerance Capacity. Forests 2025, 16, 1007. https://doi.org/10.3390/f16061007
Yang F, Xu R, Zhu C, Ji H, Shao JF, Huang K. Physiological and Transcriptomic Insights into Lead Uptake and Tolerance in Moso Bamboo (Phyllostachys edulis) Highlight Its Strong Lead Tolerance Capacity. Forests. 2025; 16(6):1007. https://doi.org/10.3390/f16061007
Chicago/Turabian StyleYang, Fan, Rong Xu, Chenyang Zhu, Haibao Ji, Ji Feng Shao, and Kangkang Huang. 2025. "Physiological and Transcriptomic Insights into Lead Uptake and Tolerance in Moso Bamboo (Phyllostachys edulis) Highlight Its Strong Lead Tolerance Capacity" Forests 16, no. 6: 1007. https://doi.org/10.3390/f16061007
APA StyleYang, F., Xu, R., Zhu, C., Ji, H., Shao, J. F., & Huang, K. (2025). Physiological and Transcriptomic Insights into Lead Uptake and Tolerance in Moso Bamboo (Phyllostachys edulis) Highlight Its Strong Lead Tolerance Capacity. Forests, 16(6), 1007. https://doi.org/10.3390/f16061007