
Morpho-Anatomical Properties and Terpene Composition of Picea Omorika (Pančić) Purk. Needles from Bosnia and Herzegovina

 ,
,  , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
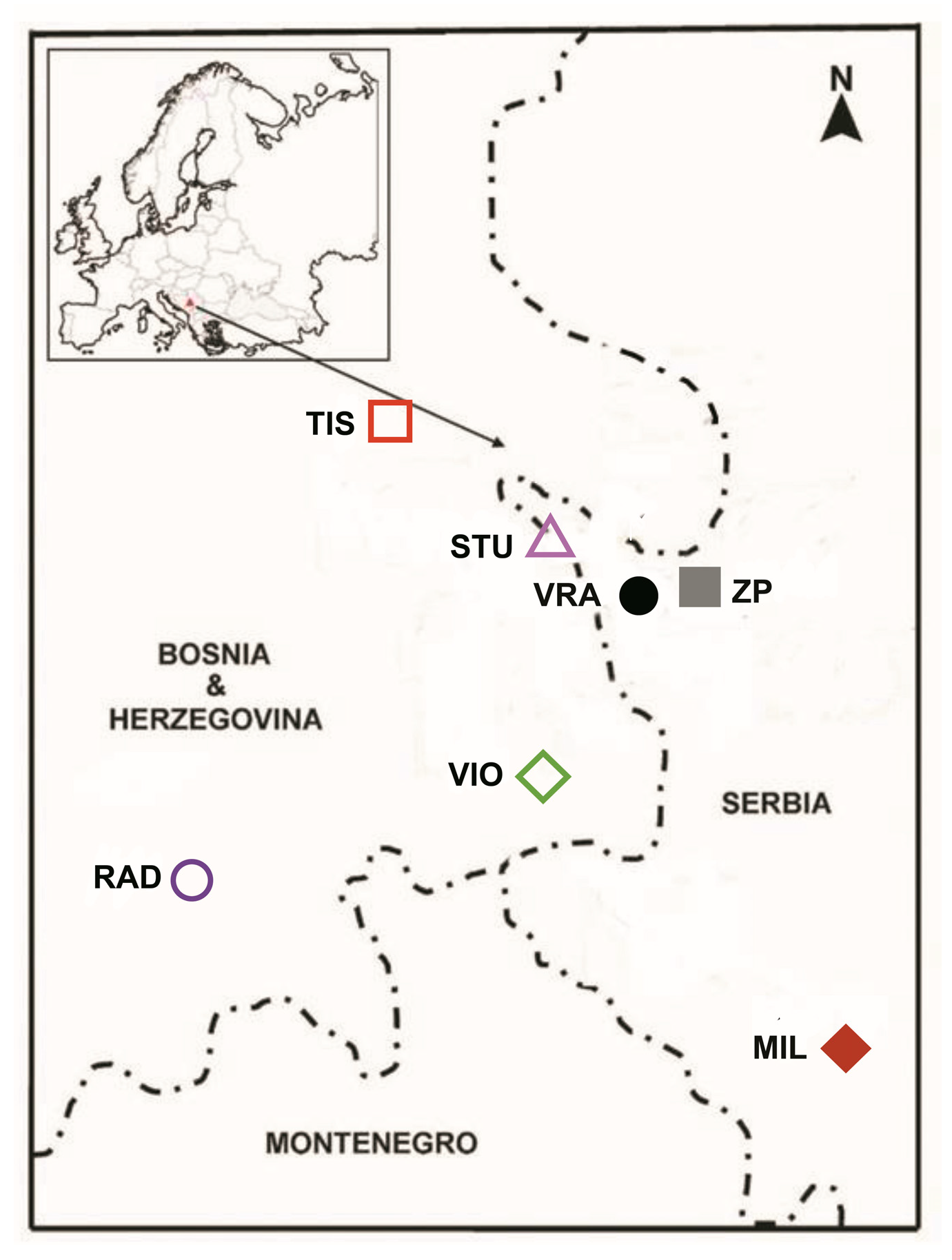
2.1. Plant Material
2.2. Morpho-Anatomical Measurements of Needles
2.3. Isolation of Volatile Compounds
2.4. GC-FID-MS Analysis
2.5. Statistical Analyses
3. Results
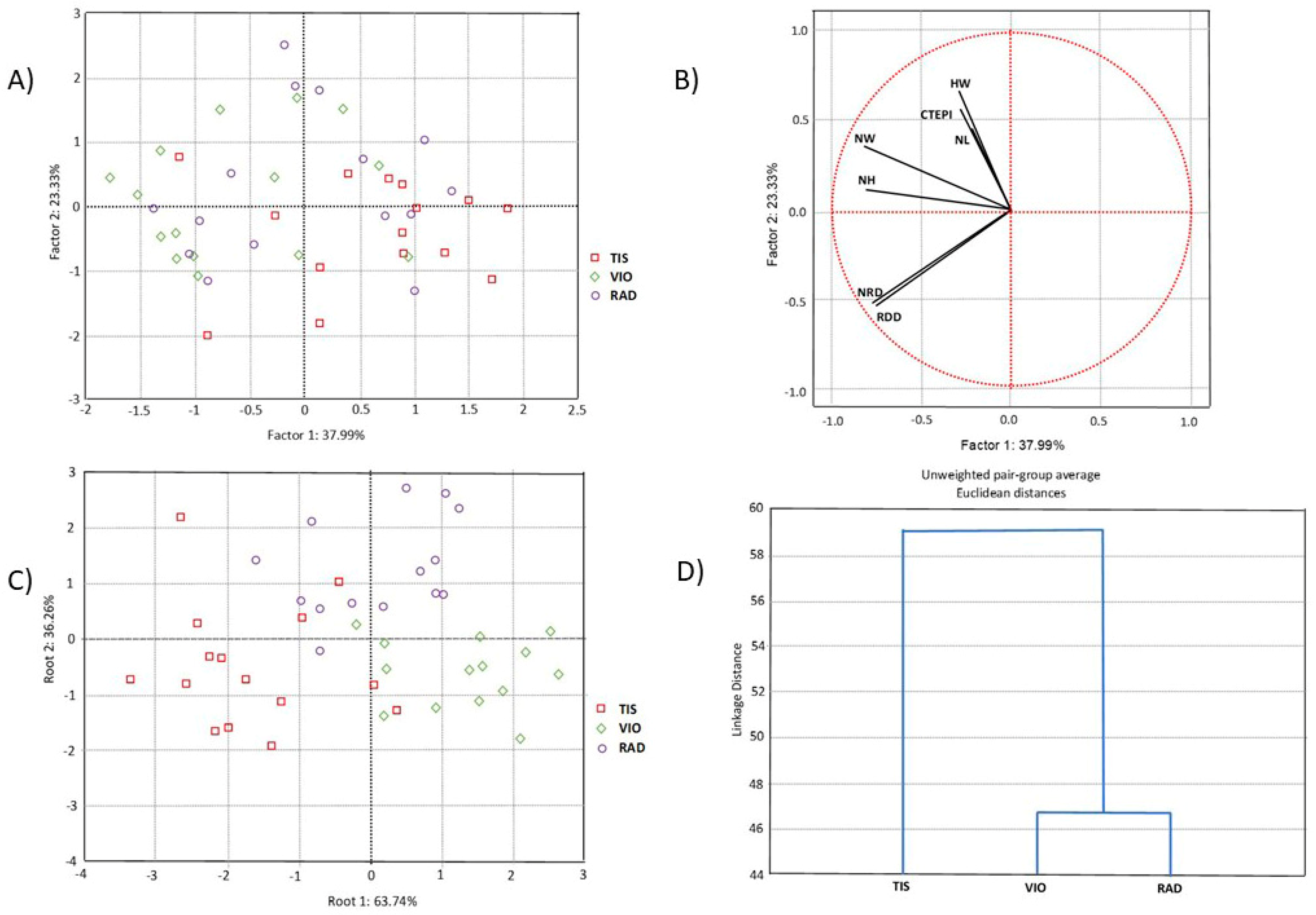
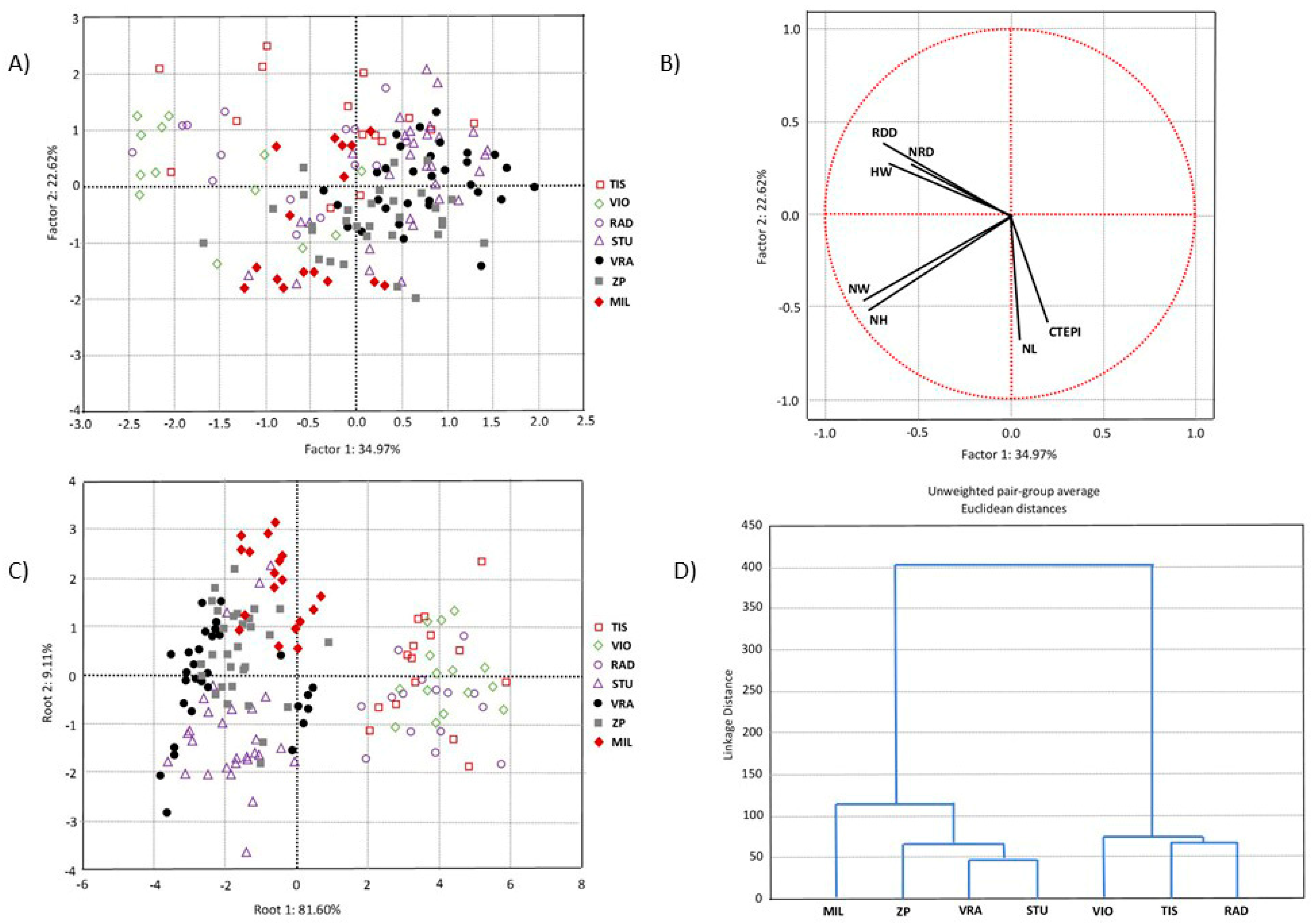
3.1. Morpho-Anatomical Characteristics of Needles and Population Variability
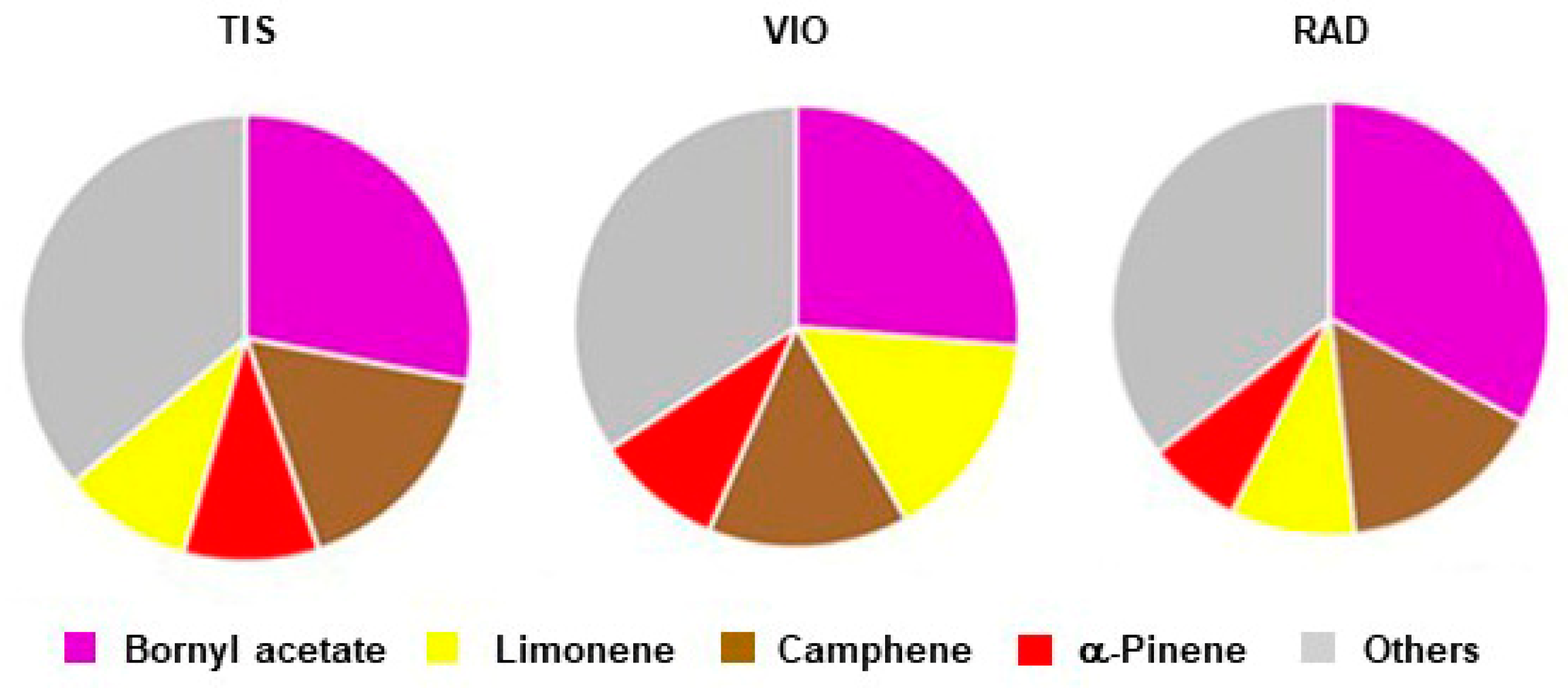
3.2. Terpene Composition of Needles and Population Variability
4. Discussion
4.1. Morpho-Anatomy of Serbian Spruce Needles
4.2. Terpene Composition of Serbian Spruce Needles
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vidaković, M.; Franjić, J. Golosjemenjače; Šumarski Fakultet Sveučilišta u Zagrebu, Hrvatske Šume, Akademija Šumarskih Znanosti: Zagreb, Croatia, 2004. [Google Scholar]
- Mataruga, M.; Milanović, Đ. Natural populations of Serbian spruce in the Republic of Srpska (Bosnia and Herzegovina). Glas. Šumarskog Fak. Univ. U Banjoj Luci 2020, 30, 77–113. [Google Scholar] [CrossRef]
- Fukarek, P. Staništa Pančićeve omorike nakon šumskih požara u 1946/47 godini. Šumarski List. 1951, 75, 61–75. [Google Scholar]
- Chandler, J.W.; Dale, J.E. Needle growth in Sitka spruce (Picea sitchensis): Effects of nutrient deficiency and needle position within shoots. Tree Physiol. 1990, 6, 41–56. [Google Scholar] [CrossRef]
- Vidaković, M. Četinjače. In Morfologija i Varijabilnost. Zagreb; JAZU i Sveučilišna Naklada Liber: Zagreb, Croatia, 1982. [Google Scholar]
- Weng, C.; Jackson, S.T. Species differentiation of North American spruce (Picea) based on morphological and anatomical characteristics of needles. Can. J. Bot. 2000, 78, 1367–1383. [Google Scholar] [CrossRef]
- Marco, H.F. The anatomy of spruce needles. J. Agric. Res. 1939, 58, 357–368. [Google Scholar]
- Ishii, H.; Ooishi, M.; Maruyama, Y.; Koike, T. Acclimation of shoot and needle morphology and photosynthesis of two Picea species to differences in soil nutrient availability. Tree Physiol. 2003, 23, 453–461. [Google Scholar] [CrossRef]
- Garneau, F.X.; Collin, G.; Gagnon, H.; Pichette, A. Chemical composition of the hydrosol and the essential oil of three different species of the Pinaceae family: Picea glauca (Moench) Voss., Picea mariana (Mill.) B.S.P., and Abies balsamea (L.) Mill. J. Essent. Oil-Bear. Plants 2012, 15, 227–236. [Google Scholar] [CrossRef]
- Wajs-Bonikowska, A.; Szoka, Ł.; Karna, E.; Wiktorowska-Owczarek, A.; Sienkiewicz, M. Composition and biological activity of Picea pungens and Picea orientalis seed and cone essential oils. Chem. Biodiv. 2017, 14, e1600264. [Google Scholar] [CrossRef]
- Champagne, E.; Bonin, M.; Royo, A.; Tremblay, J.-P.; Raymond, P. Predicting terpene content in dried conifer shoots using near infrared spectroscopy. J. Near Infrared Spec. 2020, 28, 308–314. [Google Scholar] [CrossRef]
- Tang, M.; Ai, Y.; Song, N.; Geng, L.; Ren, L.; Zhu, S.; Ai, Y.; Cai, S.; Zhang, Z.; Zhang, L.; et al. Chemical composition and hypnotic effect of Picea mariana essential oils. J. Essent. Oil Bear. Plants 2023, 26, 362–377. [Google Scholar] [CrossRef]
- Stevanović Janežić, T.; Isajev, V.; Lange, W. Relations of needle terpene compositions for selected species from genus Picea. J. Serb. Chem. Soc. 1994, 59, 359–365. [Google Scholar] [CrossRef]
- Ledig, F.T.; Hodgskiss, P.D.; Krutovskii, K.V.; Neale, D.B.; Eguiluz Piedra, T. Relationships among the spruces (Picea, Pinaceae) of southwestern North America. Syst. Bot. 2004, 29, 275–295. [Google Scholar] [CrossRef]
- Smith, D.E.; Klein, A.S. Phylogenetic inferences on the relationship of North American and European Picea species based on nuclear ribosomal 18S sequences and the internal transcribed spacer 1 region. Mol. Phylogenet. Evol. 1994, 3, 17–26. [Google Scholar] [CrossRef]
- Campbell, C.S.; Wright, W.A.; Cox, M.; Vining, T.F.; Major, C.S.; Arsenault, M.P. Internal transcribed spacer 1 (ITS1) in Picea (Pinaceae). Sequence divergence and structure. Mol. Phylogenet. Evol. 2005, 35, 165–185. [Google Scholar] [CrossRef] [PubMed]
- Mitić, Z.S.; Nikolić, B.M.; Stojković, J.P.; Jevtović, S.Č.; Stojanović, G.S.; Zlatković, B.K.; Marin, P.D. Morpho-anatomical characteristics and volatile profiles of Pinus nigra J. F. Arnold from the Balkan Peninsula and Southern Carpathians. Forests 2024, 15, 739. [Google Scholar] [CrossRef]
- Nikolić, B.; Tešević, V.; Bajić, D.; Bojović, S.; Marin, P.D. Needle essential oil composition of Picea omorika var. vukomanii. Chem. Nat. Compd. 2008, 44, 526–527. [Google Scholar] [CrossRef]
- Nikolić, B.; Tešević, V.; Đorđević, I.; Marin, P.D.; Bojović, S. Essential oil variability in natural populations of Picea omorika, a rare European conifer. Chem. Biodiv. 2009, 6, 193–203. [Google Scholar] [CrossRef]
- Nikolić, B.; Bojović, S.; Marin, P.D. Variability of needle morpho-anatomical characters of Picea omorika from natural populations in Serbia. Plant Biosyst. 2015, 149, 61–67. [Google Scholar] [CrossRef]
- Nikolić, B.M.; Ballian, D.A.; Đorđević, I.Z.; Rajčević, N.F.; Todosijević, M.M.; Stanković Jeremić, J.M.; Mitić, Z.S.; Bojović, S.R.; Tešević, V.V. n-Alkanes variability in natural populations of Picea omorika (Pančić) Purk. from Bosnia and Herzegovina. Biochem. Syst. Ecol. 2023, 106, 104544. [Google Scholar] [CrossRef]
- Blaženčić, J. Praktikum iz Anatomije Biljaka sa Osnovama Mikroskopske Tehnike; Naučna knjiga: Belgrade, Serbia, 1979. [Google Scholar]
- Liao, P.H.; Yang, H.H.; Wu, P.C.; Abu Bakar, N.H.; Urban, P.L. On-line coupling of simultaneous distillation-extraction using the Likens-Nickerson apparatus with gas chromatography. Anal. Chem. 2020, 92, 1228–1235. [Google Scholar] [CrossRef]
- Petrakis, P.V.; Tsitsimpikou, C.; Tzakou, O.; Couladis, M.; Vagias, C.; Roussis, V. Needle volatiles from five Pinus species growing in Greece. Flavour Fragr. J. 2001, 16, 249. [Google Scholar] [CrossRef]
- Pavlović, B.P.; Matović, M. Vukoman’s spruce-new variety of Serbian spruce in the Mileševka kanyon—Picea omorika var. vukomanii. Arch. Biol. Sci. 1994, 3–4, 27–28. [Google Scholar]
- Milovanović, J.; Ivetić, V.; Vilotić, D.; Šijačić-Nikolić, M. Morfo-anatomske karakteristike četina različitih fenogrupa omorike. Acta Herbol. 2005, 14, 41–49. [Google Scholar]
- Isajev, V.; Šijačić, M.; Vilotić, D. Promenljivost nekih svojstava dvogodišnjih četina omorike iz šest populacija sa različitim stepenom aerozagadenja. Šumarstvo 1999, 2–3, 147–157. [Google Scholar]
- Ilijin-Jug, M. Histološko-Citološke i Morfološke Promene Kod Vrste Picea Omorica na Području Pančeva u Hortikulturi, Izazvane Aerozagadenjem. Master’s Thesis, Biološki fakultet, University of Belgrade, Belgrade, Serbia, 1995; p. 153. [Google Scholar]
- Radotić, S.; Topuzović, M. Uporedna analiza anatomskih karakteristika listova vrsta Picea omorica (Pančić) Purk. i Picea excelsa (Lam.) Link. (Pinaceae) sa istog staništa. In Proceedings of the Simpozijum Proučavanje Biljnog i Životinjskog Sveta sa Aspekta Problema Zaš Tite i Unapredenja Životne Sredine-100 Godina bez Pančića 1888–1988, Kragujevac, Serbia, 29–30 September 1988; pp. 83–89. [Google Scholar]
- Vilotić, D. Anatomska grada stabala omorike Picea omorika (Pančić) Purkyne sa područja Nacionalnog parka Tara. In Nacionalnog Vegetacija Parka Tara; Gajić, M., Kojić, M., Karadžić, D., Vasiljević, M., Stanić, M., Eds.; Šumarski Fakultet Beograd: Belgrade, Serbia, 1994; pp. 33–49. [Google Scholar]
- Popović, V.; Nikolić, B.; Lučić, A.; Rakonjac, L.; Šešlija Jovanović, D.; Miljković, D. Morpho anatomical trait variability of the Norway spruce (Picea abies (L.) Karst.) needles in natural populations along elevational diversity gradient. Trees 2022, 36, 1131–1147. [Google Scholar] [CrossRef]
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. BioScience 2013, 58, 501–517. [Google Scholar] [CrossRef]
- Kännaste, A.; Copolovici, L.; Niinemets, Ü. Gas chromatography–mass spectrometry method for determination of biogenic volatile organic compounds emitted by plants. Methods Mol. Biol. 2013, 1009, 161–178. [Google Scholar] [CrossRef]
- Zhao, T.; Krokene, P.; Hu, J.; Christiansen, E.; Björklund, N.; Långström, B.; Solheim, H.; Borg-Karlson, A.K. Induced terpene accumulation in Norway spruce inhibits bark beetle colonization in a dose-dependent manner. PLoS ONE 2011, 6, e26649. [Google Scholar] [CrossRef]
- Keeling, C.I.; Bohlmann, J. Genes, enzymes and chemicals of terpenoid diversity in the constitutive and induced defence of conifers against insects and pathogens. New Phytol. 2006, 170, 657–675. [Google Scholar] [CrossRef]
- Hall, D.E.; Robert, J.A.; Keeling, C.I.; Domanski, D.; Quesada, A.L.; Jancsik, S.; Kuzyk, M.A.; Hamberger, B.; Borchers, C.H.; Bohlmann, J. An integrated genomic, proteomic, and biochemical analysis of 3-carene biosynthesis in Sitka spruce (Picea sitchensis). Plant Physiol. 2011, 157, 1034–1050. [Google Scholar] [CrossRef]
- Janežić, T.Š.; Isajev, V. Needle oil terpenes of Serbian spruce from three localities. Holz Roh Werkst. 1993, 51, 283–286. [Google Scholar] [CrossRef]
- Chalchat, J.C.; Gorunovic, M.S. Chemotaxonomy of pines native to the Balkans (IV): Variations in the composition of essential oils of Pinus omorika Pancic according to plant part and age of specimens. Pharmazie 1995, 50, 640–641. [Google Scholar]
- Von Ruddlof, E. Volatile leaf oil analysis in chemosystematic studies of North American conifers. Biochem. Syst. Ecol. 1975, 2, 131–167. [Google Scholar] [CrossRef]
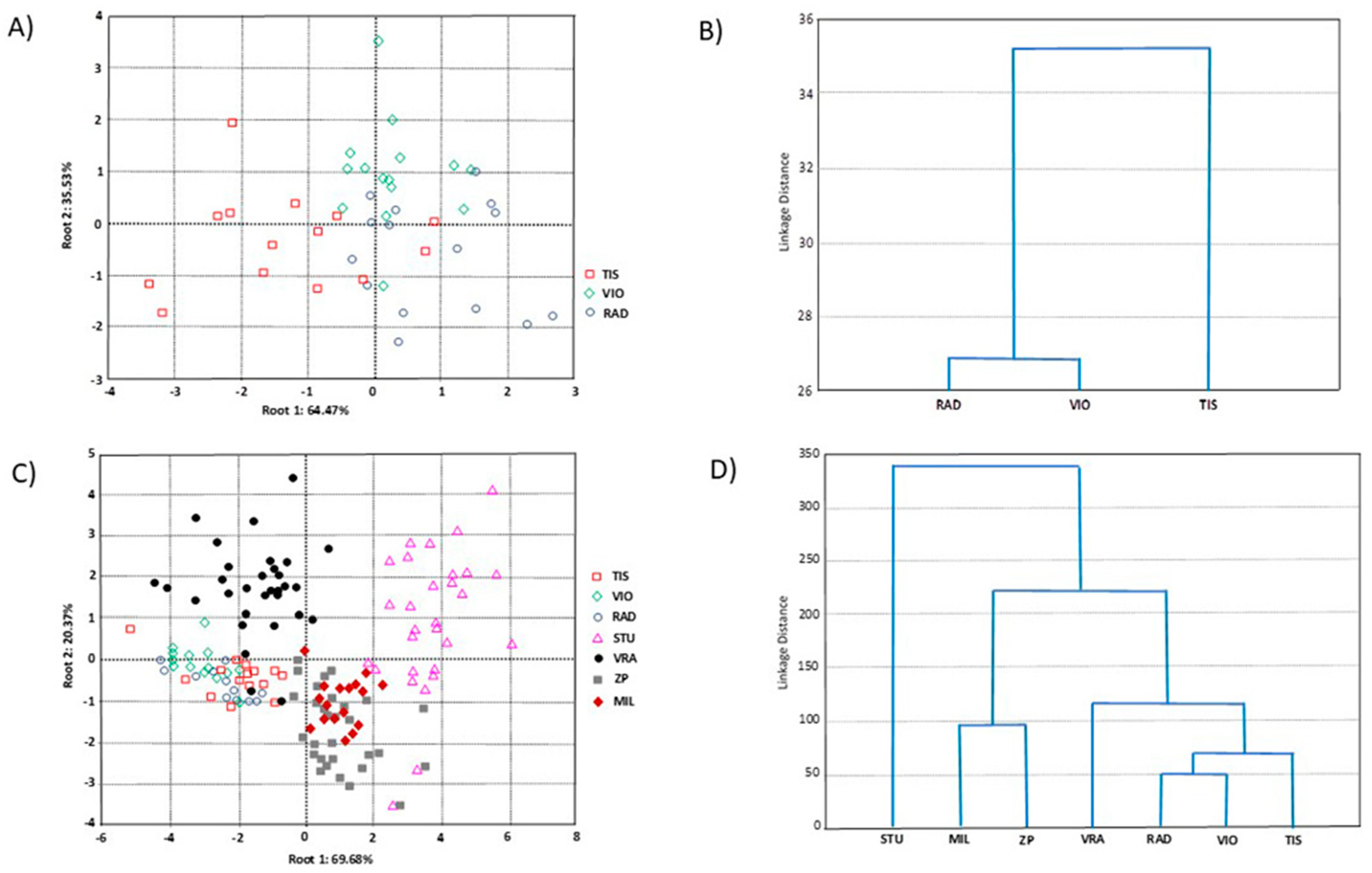

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TIS | VIO | RAD | ||||
|---|---|---|---|---|---|---|
| No. | Morpho-Anatomical Properties | F | p | n = 150 | n = 150 | n = 150 |
| X ± SD | X ± SD | X ± SD | ||||
| 1. | Needle length (mm), NL | 49.73 | *** | 11.5 ± 1.4 a | 13.3 ± 1.9 c | 12.2 ± 1.3 b |
| 2. | Needle width (µm), NW | 47.56 | *** | 1513.7 ± 214.3 a | 1706.4 ± 198.3 b | 1523.1 ± 162.4 a |
| 3. | Needle height (µm), NH | 57.27 | *** | 798.8 ± 118.65 a | 945.4 ± 108.0 c | 906.9 ± 140.3 b |
| 4. | Cuticle + epidermis width (µm), CTEPI | 7.62 | *** | 18.4 ± 3.8 a | 18.5 ± 3.4 a | 19.9 ± 3.7 b |
| 5. | Hypodermis width (µm), HW | 3.48 | * | 25.8 ± 4.3 a | 27.0 ± 3.9 b | 26.6 ± 4.2 ab |
| 6. | Number of resin ducts, NRD | 20.67 | *** | 0.5 ± 0.8 a | 1.1 ± 0.9 c | 0.8 ± 0.8 b |
| 7. | Resin duct diameter (µm), RDD | 69.70 | *** | 168.4 ± 49.4 b | 107.6 ± 26.5 a | 105.9 ± 25.5 b |
| Variables | CA 1 | CA 2 |
|---|---|---|
| Needle length (mm), NL | 0.50694 | −0.02020 |
| Needle width (µm), NW | 0.81699 | 1.12565 |
| Needle height (µm), NH | −0.13849 | −1.41322 |
| Cuticle + epidermis width(µm), CTEPI | −0.24485 | 0.71797 |
| Hypodermis width (µm), HW | 0.28981 | −0.56591 |
| Number of resin ducts, NRD | 1.89068 | 0.26420 |
| Resin duct diameter (µm), RDD | −1.72543 | −0.23710 |
| Eigenval. | 1.69895 | 0.96633 |
| Cum. Prop. | 0.63744 | 1.00000 |
| Variables | C1 | C2 | C3 | C4 | C5 | C6 |
|---|---|---|---|---|---|---|
| Needle length, NL | −0.291497 | −0.400480 | 0.02363 | −0.449454 | −0.360216 | −0.50206 |
| Needle width, NW | −0.016916 | −0.489000 | −1.49153 | 0.333592 | 0.150833 | −0.16168 |
| Needle height, NH | 0.174468 | 1.192573 | 0.73281 | −0.685388 | 0.377807 | −0.29397 |
| Cuticle + epidermis width, CTEPI | −0.367450 | 0.460102 | −0.00875 | 0.385076 | −0.745772 | −0.03169 |
| Hypodermis width, HW | 0.925749 | −0.042867 | 0.14062 | 0.101634 | −0.368507 | 0.03713 |
| Number of resin ducts, NRD | −0.60236 | −0.42842 | −0.10944 | −703417 | −0.566290 | 0.98349 |
| Resin duct diameter, RDD | 0.057210 | 0.154574 | 0.31091 | 0.196992 | 0.035784 | −1.39181 |
| Eigenvalue | 6.870889 | 0.767062 | 0.49859 | 0.213854 | 0.066353 | 0.00301 |
| Cum. Prop. | 0.816044 | 0.907146 | 0.96636 | 0.991762 | 0.999643 | 1.00000 |
| No. | RT | Compound | RI | TIS | VIO | RAD |
|---|---|---|---|---|---|---|
| X ± SD | X ± SD | X ± SD | ||||
| 1 | 3.223 | Hexanal | 798 | 0.8 ± 0.6 | 0.3 ± 0.5 | 0.2 ± 0.2 |
| 2 | 4.061 | (E)-hex-2-enal | 847 | 1.4 ± 0.9 | 0.4 ± 0.5 | 0.6 ± 0.6 |
| 3 | 4.283 | n-Hexanol | 860 | 0.2 ± 0.1 | 0.2 ± 0.3 | 0.1 ± 0.1 |
| 4 | 4.643 | Santene | 881 | 3.2 ± 0.6 | 5.0 ± 0.9 | 4.1 ± 0.8 |
| 5 | 5.511 | Tricyclene | 919 | 1.5 ± 0.3 | 1.3 ± 0.3 | 1.2 ± 0.3 |
| 6 | 5.801 | α-Pinene | 930 | 9.7 ± 3.7 | 9.4 ± 2.6 | 7.0 ± 1.9 |
| 7 | 6.212 | Camphene | 945 | 16.8 ± 2.2 | 14.7 ± 1.7 | 15.4 ± 2.6 |
| 8 | 6.551 | Benzaldehyde | 961 | 0.3 ± 0.2 | tr | 0.1 ± 0.1 |
| 9 | 6.909 | Sabinene | 969 | 0.1 ± 0.1 | tr | |
| 10 | 7.011 | β-Pinene | 972 | 1.2 ± 0.3 | 1.4 ± 0.4 | 1.3 ± 0.6 |
| 11 | 7.389 | Myrcene | 985 | 4.0 ± 1.3 | 2.9 ± 0.9 | 2.3 ± 0.6 |
| 12 | 7.529 | Mesitylene | 991 | tr | ||
| 13 | 7.735 | n-Octanal | 999 | 0.1 ± 0.1 | ||
| 14 | 7.859 | α-Phellandrene | 1002 | 1.3 ± 0.4 | 1.4 ± 0.6 | 1.1 ± 0.3 |
| 15 | 8.068 | δ-Car-3-ene | 1007 | 0.3 ± 0.2 | 0.1 ± 0.0 | 0.1 ± 0.1 |
| 16 | 8.301 | α-Terpinene | 1013 | 0.1 ± 0.1 | tr | 0.1 ± 0.1 |
| 17 | 8.555 | p-Cymene | 1021 | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 |
| 18 | 8.699 | Limonene | 1025 | 9.3 ± 3.0 | 15.5 ± 7.0 | 9.3 ± 7.9 |
| 19 | 8.786 | 1,8-Cineole | 1027 | tr | 0.1 ± 0.2 | 0.4 ± 0.3 |
| 20 | 9.342 | trans-β-Ocimene | 1040 | tr | ||
| 21 | 9.805 | γ-Terpinene | 1054 | 0.1 ± 0.1 | tr | |
| 22 | 10.940 | Terpinolene | 1085 | 0.6 ± 0.2 | 0.5 ± 0.2 | 0.4 ± 0.1 |
| 23 | 10.993 | 72 (100), 71 (100), 43 (78); MW 140 | 1086 | 0.4 ± 0.1 | 0.7 ± 0.2 | 0.5 ± 0.1 |
| 24 | 11.095 | 108 (100), 93 (47), 109 (13); MW 152 | 1089 | 0.1 ± 0.0 | 0.1 ± 0.1 | 0.1 ± 0.0 |
| 25 | 11.381 | Linalool | 1096 | 0.2 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| 26 | 11.465 | 79 (100), 94 (88), 91 (23); MW 138 | 1099 | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 |
| 27 | 11.476 | n-Nonanal | 1099 | 0.7 ± 0.7 | ||
| 28 | 11.946 | endo-Fenchol | 1110 | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 |
| 29 | 12.064 | 3-Methylbut-3-enyl 3-methylbutanoate | 1114 | tr | ||
| 30 | 12.276 | cis-p-Menth-2-en-1-ol | 1118 | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.1 ± 0.1 |
| 31 | 12.476 | α-Campholenal | 1122 | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 |
| 32 | 13.016 | trans-Pinocarveol | 1135 | 0.1 ± 0.1 | 0.3 ± 0.1 | 0.2 ± 0.1 |
| 33 | 13.283 | Isopulegol | 1141 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| 34 | 13.387 | Camphene hydrate | 1143 | 0.5 ± 0.4 | 0.9 ± 0.6 | 0.9 ± 0.5 |
| 35 | 13.594 | Citronellal | 1148 | 0.7 ± 0.4 | 1.0 ± 0.6 | 1.1 ± 0.6 |
| 36 | 13.751 | Isoborneol | 1152 | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 |
| 37 | 14.130 | Borneol | 1161 | 2.2 ± 1.2 | 4.3 ± 1.5 | 4.1 ± 1.1 |
| 38 | 14.669 | Terpinen-4-ol | 1173 | 0.3 ± 0.2 | 0.2 ± 0.1 | 0.2 ± 0.1 |
| 39 | 15.040 | p-Cymen-8-ol | 1181 | tr | tr | tr |
| 40 | 15.234 | α-Terpineol | 1186 | 0.4 ± 0.2 | 0.5 ± 0.2 | 0.4 ± 0.2 |
| 41 | 15.391 | cis-Piperitol | 1192 | tr | tr | |
| 42 | 15.471 | Dihydro carveol | 1191 | tr | ||
| 43 | 15.499 | γ-Terpineol | 1192 | tr | ||
| 44 | 15.702 | Homomyrtenol | 1198 | 0.1 ± 0.1 | 0.2 ± 0.1 | 0.1 ± 0.1 |
| 45 | 15.996 | trans-Piperitol | 1204 | tr | tr | tr |
| 46 | 15.999 | Verbenone | 1204 | tr | ||
| 47 | 16.469 | endo-Fenchyl acetate | 1214 | 0.2 ± 0.2 | 0.2 ± 0.1 | 0.2 ± 0.1 |
| 48 | 16.821 | Citronellol | 1223 | 1.4 ± 0.7 | 2.4 ± 1.2 | 2.1 ± 1.3 |
| 49 | 17.493 | Neral | 1237 | tr | 0.1 ± 0.1 | tr |
| 50 | 17.498 | Nojigiku acetate | 1237 | 0.1 ± 0.0 | 0.1 ± 0.0 | tr |
| 51 | 17.986 | Piperitone | 1249 | 0.1 ± 0.2 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| 52 | 18.010 | Geraniol | 1249 | 0.3 ± 0.2 | 0.6 ± 0.4 | 0.5 ± 0.4 |
| 53 | 18.743 | 69 (100), 41 (68), 119 (30); MW 152 | 1266 | 0.2 ± 0.2 | 0.2 ± 0.2 | tr |
| 54 | 19.075 | (Z)-Undec-6-en-2-one | 1273 | 0.2 ± 0.2 | 0.1 ± 0.1 | |
| 55 | 19.228 | cis-Verbenyl acetate | 1276 | tr | tr | tr |
| 56 | 19.525 | Bornyl acetate | 1285 | 28.2 ± 4.5 | 26.6 ± 4.1 | 33.1 ± 4.9 |
| 57 | 20.010 | 107 (100), 43 (62), 150 (59); MW 150 | 1294 | tr | tr | |
| 58 | 20.038 | trans-Pinocarvyl acetate | 1295 | tr | 0.1 ± 0.1 | 0.2 ± 0.1 |
| 59 | 20.219 | 6-Hydroxycarvotanacetone | 1300 | tr | ||
| 60 | 20.706 | 69 (100), 41 (69), 95 (52); MW 170 | 1310 | tr | 0.2 ± 0.1 | 0.1 ± 0.1 |
| 61 | 20.955 | 81 (100), 67 (29), 153 (26); MW 153 | 1316 | tr | tr | |
| 62 | 21.591 | 121 (100), 93 (62), 108 (56); MW 196 | 1331 | tr | tr | 0.2 ± 0.2 |
| 63 | 22.295 | α-Terpinyl acetate | 1346 | 0.2 ± 0.1 | 0.3 ± 0.1 | 0.5 ± 0.2 |
| 64 | 22.470 | Citronellyl acetate | 1350 | 0.2 ± 0.1 | 0.3 ± 0.2 | 0.3 ± 0.1 |
| 65 | 22.569 | Neric acid | 1353 | tr | tr | |
| 66 | 22.987 | Neryl acetate | 1362 | tr | tr | tr |
| 67 | 23.802 | Geranyl acetate | 1382 | 0.6 ± 0.6 | 1.0 ± 0.8 | 2.2 ± 1.0 |
| 68 | 24.167 | β-Elemene | 1390 | tr | ||
| 69 | 24.821 | Dodecanal | 1403 | tr | tr | |
| 70 | 25.337 | (E)-Caryophyllene | 1417 | 0.1 ± 0.1 | tr | tr |
| 71 | 25.688 | (E)-α-Ionone | 1426 | 0.1 ± 0.1 | tr | 0.1 ± 0.2 |
| 72 | 25.847 | cis-Cinnamic acid | 1429 | tr | tr | |
| 73 | 26.784 | α-Humulene | 1452 | tr | tr | |
| 74 | 27.135 | cis-Muurola-4(15),5-diene | 1461 | tr | ||
| 75 | 27.585 | trans-Cadina-1(6),4-diene | 1470 | tr | tr | |
| 76 | 27.757 | γ-Muurolene | 1475 | 0.1 ± 0.0 | tr | 0.1 ± 0.1 |
| 77 | 27.940 | Germacrene D | 1479 | 0.2 ± 0.1 | 0.4 ± 0.6 | 0.2 ± 0.2 |
| 78 | 28.111 | (E)-β-Ionone | 1484 | 0.2 ± 0.1 | 0.1 ± 0.0 | 0.2 ± 0.1 |
| 79 | 28.406 | trans-Muurola-4(14),5-diene | 1491 | tr | ||
| 80 | 28.460 | α-Selinene | 1492 | tr | tr | |
| 81 | 28.501 | epi-Cubebol | 1493 | tr | ||
| 82 | 28.738 | α-Muurolene | 1498 | 0.2 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.1 |
| 83 | 29.303 | γ-Cadinene | 1512 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.2 ± 0.1 |
| 84 | 29.685 | δ-Cadinene | 1522 | 0.7 ± 0.2 | 0.5 ± 0.2 | 0.7 ± 0.2 |
| 85 | 30.117 | 177 (100), 159 (82), 220 (70); MW 220 | 1532 | tr | tr | |
| 86 | 30.253 | α-Cadinene | 1536 | tr | tr | |
| 87 | 31.177 | (E)-Nerolidol | 1559 | 0.2 ± 0.1 | tr | |
| 88 | 31.231 | Dodecanoic acid | 1560 | tr | ||
| 89 | 31.776 | Germacrene D-4-ol | 1573 | 0.1 ± 0.1 | 0.3 ± 0.3 | 0.2 ± 0.1 |
| 90 | 31.849 | Spathulenol | 1575 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.1 ± 0.0 |
| 91 | 32.092 | 93 (100), 147 (96), 105 (96); MW 222 | 1580 | tr | ||
| 92 | 33.229 | 43 (100), 119 (68), 109 (52); MW 254 | 1609 | tr | tr | tr |
| 93 | 33.343 | 1,10-di-epi-Cubenol | 1612 | tr | tr | |
| 94 | 33.860 | 1-epi-Cubenol | 1626 | 0.1 ± 0.1 | tr | 0.1 ± 0.0 |
| 95 | 34.380 | τ-Cadinol | 1638 | 1.3 ± 0.5 | 0.8 ± 0.3 | 1.1 ± 0.3 |
| 96 | 34.553 | α-Muurolol | 1644 | 0.3 ± 0.1 | 0.1 ± 0.1 | 0.2 ± 0.1 |
| 97 | 34.874 | α-Cadinol | 1652 | 2.2 ± 0.9 | 1.2 ± 0.5 | 1.8 ± 0.6 |
| 98 | 35.103 | 191 (100), 119 (37), 121 (24); MW 234 | 1658 | tr | tr | 0.1 ± 0.0 |
| 99 | 36.091 | Eudesma-4(15),7-dien-1-β-ol | 1684 | tr | ||
| 100 | 36.745 | 177 (100), 159 (76), 220 (36); MW 220 | 1701 | tr | 0.1 ± 0.1 | 0.1 ± 0.1 |
| 101 | 37.527 | (2Z,6E)-Farnesol | 1723 | 0.1 ± 0.1 | ||
| 102 | 38.014 | Oplopanone | 1736 | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.3 ± 0.1 |
| 103 | 38.810 | Tetradecanoic acid | 1757 | tr | tr | |
| 104 | 40.854 | Hexadecanal | 1813 | 0.4 ± 0.3 | tr | tr |
| 105 | 41.058 | 93 (100), 81 (86), 95 (83); MW 220 | 1819 | tr | tr | |
| 106 | 41.664 | Neophytadiene | 1836 | tr | ||
| 107 | 44.420 | Heptadecanal | 1915 | 0.1 ± 0.1 | ||
| 108 | 45.836 | Hexadecanoic acid | 1836 | 0.2 ± 0.2 | tr | tr |
| 109 | 46.471 | Hex-5-enyl dodecanoate | 1976 | tr | tr | |
| 110 | 46.452 | Hexyl dodecanoate | 1979 | tr | tr | |
| 111 | 46.802 | Octadec-9-enal | 1986 | 0.5 ± 0.5 | tr | |
| 112 | 46.952 | Manool oxide | 1990 | 0.1 ± 0.1 | tr | tr |
| 113 | 47.688 | 13-epi-Manool oxide | 2012 | 0.1 ± 0.2 | tr | |
| 114 | 47.839 | Octadecanal | 2018 | 0.3 ± 0.5 | ||
| 115 | 48.147 | (E,E)-Geranyl linalool | 2026 | tr | ||
| 116 | 48.944 | 81 (100), 105 (83), 55 (81); MW 272 | 2051 | tr | ||
| 117 | 50.925 | Phytol | 2114 | 1.2 ± 1.0 | 0.5 ± 0.3 | 0.9 ± 0.8 |
| 118 | 54.131 | 83 (100), 95 (34), 55 (33); MW 286 | 2219 | tr | ||
| 119 | 54.575 | Palustral | 2235 | tr | ||
| 120 | 55.469 | Dehydroabietal | 2266 | tr | ||
| 121 | 55.968 | 109 (100), 149 (98), 119 (91); MW 290 | 2283 | tr | ||
| 122 | 56.350 | Tricosane | 2296 | 0.1 ± 0.1 | tr | 0.1 ± 0.1 |
| 123 | 60.019 | 95 (100), 69 (98), 181 (95); MW 306 | 2429 | tr | ||
| 124 | 61.745 | Pentacontane | 2500 | 0.1 ± 0.1 | tr | tr |
| 125 | 63.588 | Dodecyl dodecanoate | 2564 | 0.1 ± 0.1 | ||
| 126 | 70.084 | Squalene | 2823 | 0.2 ± 0.2 | 0.1 ± 0.2 | 0.1 ± 0.1 |
| 127 | 75.972 | Nonacosan-10-one | 3082 | 0.1 ± 0.1 | tr | |
| 128 | 76.622 | Nonacosan-10-ol | 3114 | 1.1 ± 1.0 | 0.3 ± 0.3 | 1.0 ± 0.9 |
| Total | 100.0 | 100.0 | 100.0 | |||
| Monoterpenes | 84.8 | 92.6 | 90.0 | |||
| Monoterpene hydrocarbons | 48.6 | 52.8 | 42.8 | |||
| Oxygenated monoterpenes | 36.2 | 39.8 | 47.2 | |||
| Sesquiterpenes | 6.2 | 3.9 | 5.5 | |||
| Sesquiterpene hydrocarbons | 1.7 | 1.2 | 1.6 | |||
| Oxygenated sesquiterpenes | 4.5 | 2.7 | 3.9 | |||
| Diterpenes | 1.4 | 0.5 | 0.9 | |||
| Diterpene hydrocarbons | tr | |||||
| Oxygenated diterpenes | 1.4 | 0.5 | 0.9 | |||
| Triterpenes | 0.2 | 0.1 | 0.1 | |||
| Triterpene hydrocarbons | 0.2 | 0.1 | 0.1 | |||
| Aliphatic aldehydes and alcohols | 5.6 | 1.3 | 1.9 | |||
| Others * | 1.0 | 0.1 | 0.3 | |||
| Non identified (n.i.) | 0.8 | 1.5 | 1.3 |
| Variables | Root 1 | Root 2 |
|---|---|---|
| Tricyclene | −0.81228 | 0.291029 |
| α-Pinene | 0.06454 | 0.091758 |
| Camphene | −0.02618 | −0.387539 |
| β-Pinene | 0.48200 | −0.065597 |
| Bornyl acetate | 0.55503 | −0.505837 |
| δ-Cadinene | 0.88663 | 0.059749 |
| τ-Cadinol | −1.18635 | −0.623445 |
| Eigenval. | 0.85073 | 0.468831 |
| Cum. Prop. | 0.64471 | 1.000000 |
| Variables | Root 1 | Root 2 | Root 3 | Root 4 | Root 5 |
|---|---|---|---|---|---|
| Tricyclene | 0.591065 | −0.083273 | −0.016057 | −0.420899 | −0.506909 |
| α-Pinene | 1.016893 | −0.011394 | −0.178902 | −0.337778 | −0.767944 |
| Camphene | 0.247062 | −0.294177 | 0.012248 | 0.942534 | 0.829394 |
| β-Pinene | −0.187520 | 0.106547 | −0.205821 | −0.303272 | 0.221453 |
| Bornyl acetate | 0.910007 | −0.510325 | 0.603289 | −0.566169 | −0.039241 |
| δ-Cadinene | 0.558879 | 0.841102 | 0.462885 | 0.108121 | 0.153127 |
| τ-Cadinol | 0.899374 | −0.462104 | −0.847024 | −0.370009 | 0.447749 |
| Eigenvalue | 5.626603 | 1.644619 | 0.724582 | 0.055664 | 0.023865 |
| Cum. Prop. | 0.696757 | 0.900414 | 0.990141 | 0.997034 | 0.999989 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolić, B.M.; Mitić, Z.S.; Ballian, D.; Todosijević, M.M.; Nikolić, J.S.; Ivanović, S.; Tešević, V.V. Morpho-Anatomical Properties and Terpene Composition of Picea Omorika (Pančić) Purk. Needles from Bosnia and Herzegovina. Forests 2025, 16, 791. https://doi.org/10.3390/f16050791
Nikolić BM, Mitić ZS, Ballian D, Todosijević MM, Nikolić JS, Ivanović S, Tešević VV. Morpho-Anatomical Properties and Terpene Composition of Picea Omorika (Pančić) Purk. Needles from Bosnia and Herzegovina. Forests. 2025; 16(5):791. https://doi.org/10.3390/f16050791
Chicago/Turabian StyleNikolić, Biljana M., Zorica S. Mitić, Dalibor Ballian, Marina M. Todosijević, Jelena S. Nikolić, Stefan Ivanović, and Vele V. Tešević. 2025. "Morpho-Anatomical Properties and Terpene Composition of Picea Omorika (Pančić) Purk. Needles from Bosnia and Herzegovina" Forests 16, no. 5: 791. https://doi.org/10.3390/f16050791
APA StyleNikolić, B. M., Mitić, Z. S., Ballian, D., Todosijević, M. M., Nikolić, J. S., Ivanović, S., & Tešević, V. V. (2025). Morpho-Anatomical Properties and Terpene Composition of Picea Omorika (Pančić) Purk. Needles from Bosnia and Herzegovina. Forests, 16(5), 791. https://doi.org/10.3390/f16050791