
Induction of Somatic Embryogenesis in Araucaria araucana (Molina) K. Koch: Considerations for Ex Situ Conservation of Ancient Tree in Chile

 , ,
, ,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Material and Methods
2.1. Plant Material
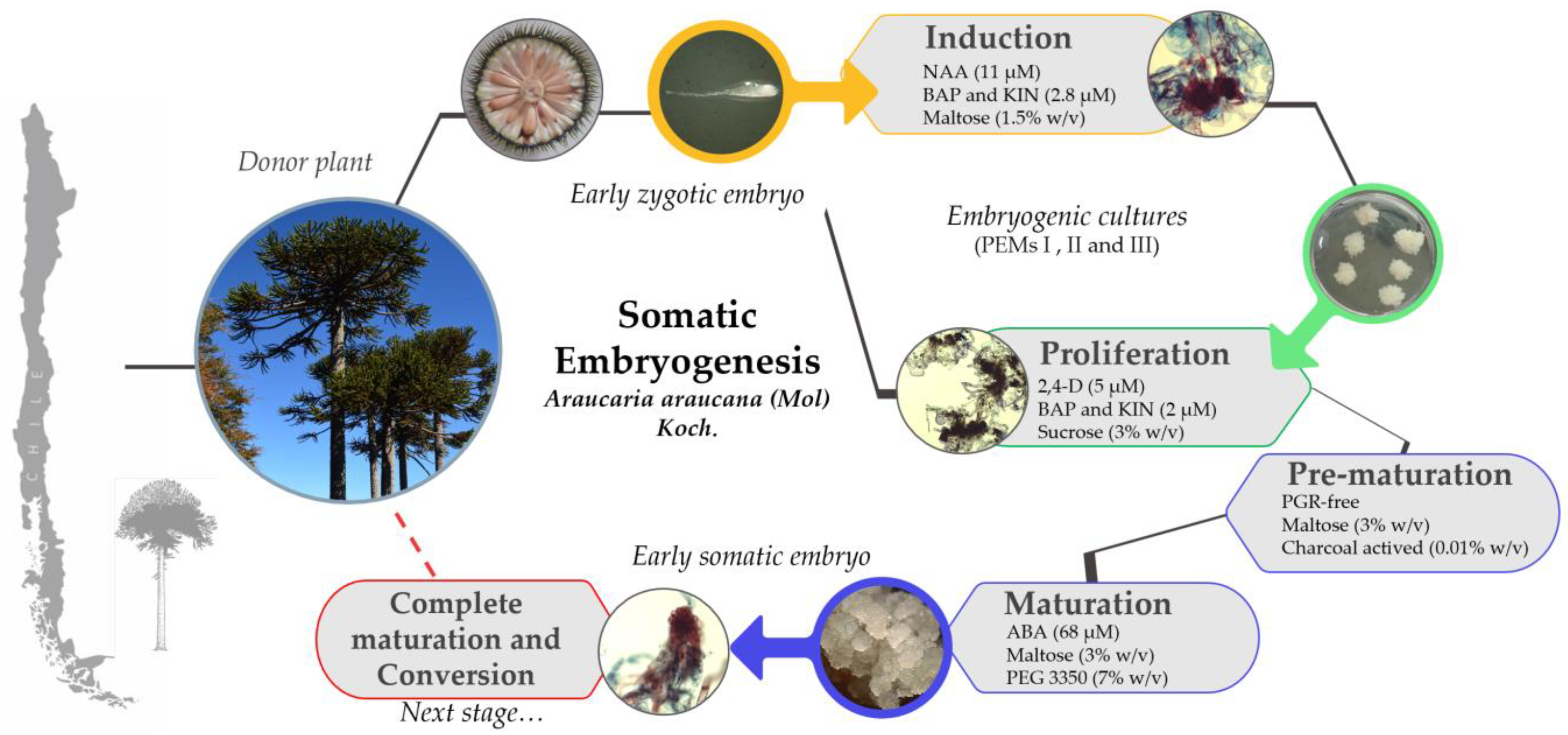
2.2. Induction of Embryogenic Culture
2.3. Embryogenic Culture Proliferation
2.4. Somatic Embryo Development in Pre-Maturation and Maturation
2.5. Morpho-Cytochemical Analysis
2.6. Statistical Analysis
3. Results
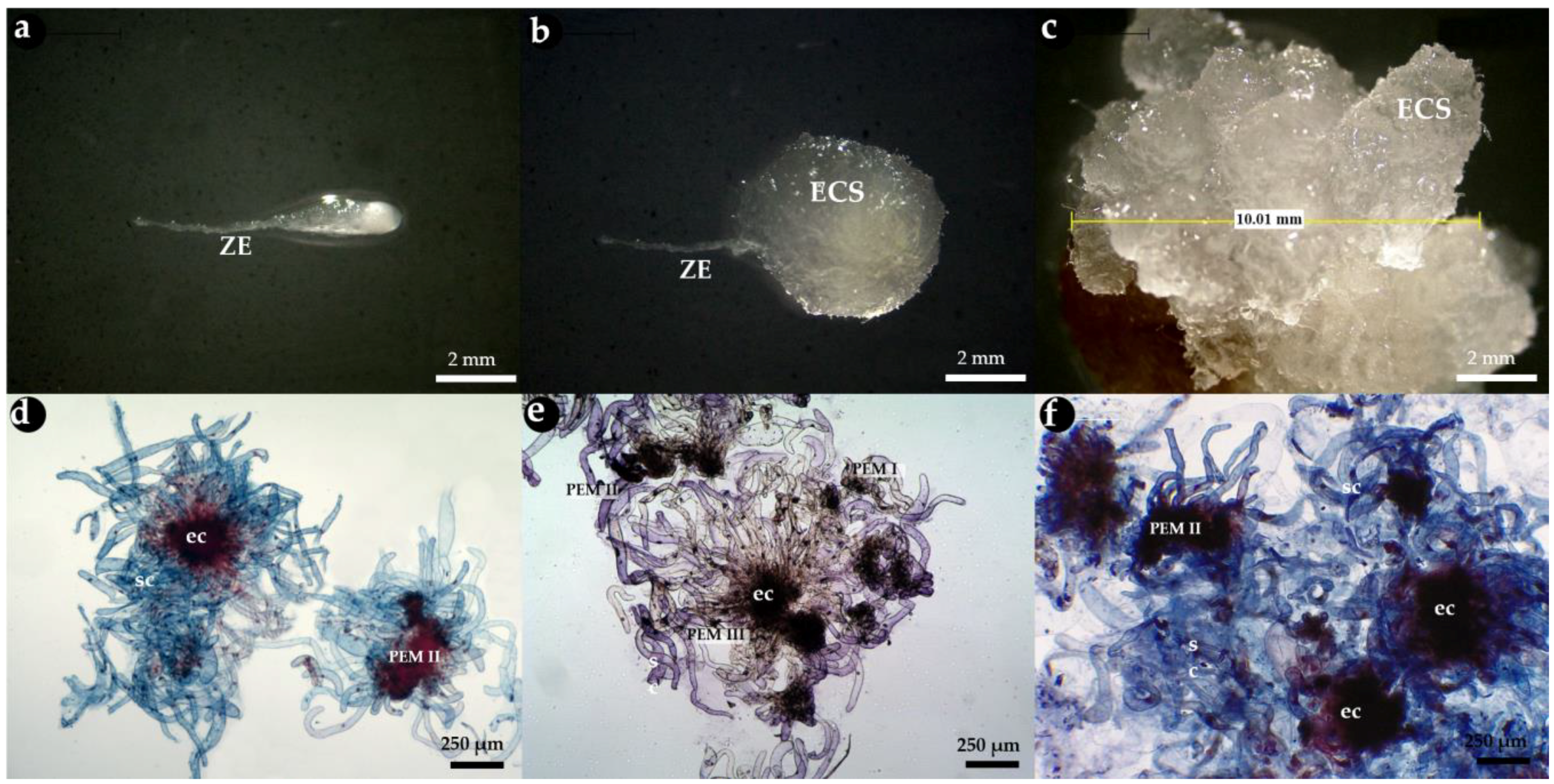
3.1. Zygotic Embryo Development Stage Is Affected by Provenance and Embryogenic Culture Induction
3.2. Embryogenic Culture Proliferation and Characterization of Pro-Embryogenic Masses
3.3. Pre-Maturation and Maturation of Embryogenic Culture
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| BM | Basal medium [7] |
| Ec | Embryogenic cell |
| Sc | Suspensor cell |
| ECLs | Embryogenic cell lines |
| FM | Fresh mass |
| IC | Induction medium control (PGR-free BM) |
| Kin | Kinetin |
| MA | Malalcahuello |
| MIP | Induction medium (BM with 11 μM NAA and others) |
| MIN | Induction medium (BM with 23 μM 2,4-D and others) |
| MP1 | Proliferation medium 1 (BM with 5 μM 2,4-D and others) |
| MP2 | Proliferation medium 2 (BM with 23 μM 2,4-D and others) |
| NAA | 1-naftalenacetic acid |
| NaClO | Sodium hypochlorite |
| PEG | Polyethyleneglycol 3350 |
| PEMs | Pro-embryogenic masses |
| PGR | Plant growth regulator |
| sc | Suspensor-like cell |
| SE | Somatic embryogenesis |
| TR | Trongol |
| VA | Villa las Araucarias |
| ZE | Zygotic embryo |
| 2,4-D | 2,4-diclorofenoxiacetic acid |
| 6-BA | 6-benzylaminopurine |
Appendix A
References
- González, M.; Cortés, M.; Izquierdo, F.; Gallo, L.; Echeverría, C.; Bekessy, S.; Montaldo, P. Araucaria araucana (Molina) K. Koch. Araucaria (o), Pehuén, Pino piñonero, Pino de Neuquén, Monkey Puzzle Tree. In Las especies arbóreas de los bosques templados de Chile y Argentina; Donoso, C., Ed.; Autoecología, Marisa Cuneo Ediciones: Valdivia, Chile, 2006; pp. 36–53. [Google Scholar]
- Zamorano, C.; Cortés, M.; Echeverría, C.; Hechenleitner, P.; Lara, A. Experiencias de Restauración con Especies Forestales Amenazadas En Chile. In Restauración de Bosques en América Latina; FIRE/Mundi-Prensa: Ciudad de México, México, 2008; pp. 18–36. [Google Scholar]
- Herrmann, T. Knowledge, values, uses and management of the Araucaria Araucana forest by the indigenous Mapuche Pewenche people: A basis for collaborative natural resource management in southern Chile. Nat. Resour. Forum 2005, 29, 120–134. [Google Scholar] [CrossRef]
- Veblen, T.; Burns, R.; Kitzberger, T.; Lara, A.; Villalba, R. The ecology of the conifers of southern South America. In Ecology of the Southern Conifers; Enright, N., Hill, R., Eds.; Melbourne University Press: Parkville, Australia, 1995; pp. 120–155. [Google Scholar]
- IUCN. The IUCN Red list of threatened species. In Red List of Threatened Species; 2023. Available online: https://www.iucnredlist.org/species/32975/2829141 (accessed on 13 March 2024).
- Balocchi, F.; Wingfield, M.; Ahumada, R.; Barnes, I. Pewenomyces kutranfy gen. nov. et sp. nov. causal agent of an important canker disease on Araucaria araucana in Chile. Plant Pathol. 2021, 70, 1243–1259. [Google Scholar] [CrossRef]
- Instituto Forestal (INFOR). INFOR and CMPC Sign Agreement for the Genetic Protection of the Araucaria. 21 June 2018. Available online: https://www.infor.cl/index.php/noticias/425-infor-y-cmpc-firman-convenio-para-el-resguardo-genetico-de-la-araucaria (accessed on 22 June 2018).
- Ministerio del Medio Ambiente. Araucaria araucana (Mol.) Koch, M.K. Species Classification: Species Background Sheet 2002, pp. 1–7. Available online: https://clasificacionespecies.mma.gob.cl/wp-content/uploads/2019/10/Araucaria_araucana_FINAL.pdf (accessed on 1 March 2024).
- Convention on Biological Diversity. 1992. Available online: https://www.cbd.int/ (accessed on 12 November 2023).
- Ruiz, E.; González, F.; Torres-Díaz, C.; Fuentes, G.; Mardones, M.; Stuessy, T.; Samuel, R.; Becerra, J.; Silva, M. Genetic diversity and differentiation within and among Chilean populations of Araucaria araucana (Araucariaceae) based on allozyme variability. Taxon 2007, 56, 1221–1228. [Google Scholar] [CrossRef]
- Goeten, D.; Farias-Soares, F.; Rogge-Renner, G.D.; Pereira, M.L.T.; Walters, C.; Silveira, V.; Catarina, C.S.; Guerra, M.P.; Steiner, N. Carbohydrate and dehydrin-like protein profiles during Araucaria angustifolia seed development provides insights toward ex situ conservation. Trees 2023, 37, 1–15. [Google Scholar] [CrossRef]
- Lineros, Y.; Balocchi, C.; Muñoz, X.; Sánchez, M.; Ríos, D. Cryopreservation of Pinus radiata embryogenic tissue: Effects of cryoprotective pretreatments on maturation ability. Plant Cell Tissue Organ Cult. 2018, 135, 357–366. [Google Scholar] [CrossRef]
- Stefenon, V.M.; Marques-Pinheiro, V.; Goeten, D.; Ree, J.F.; Steiner, N.; Guerra, M.P. Advances and constraints in somatic embryogenesis of Araucaria angustifolia, Acca sellowiana and Bactris gasipaes. Plant Cell Tissue Organ Cult. 2020, 143, 241–263. [Google Scholar] [CrossRef]
- Steiner, N.; Farias-Soares, F.; Schmidt, É.; Pereira, M.; Scheid, B.; Rogge-Renner, G.; Bouzon, Z.; Schmidt, D.; Maldonado, S.; Guerra, M.P. Toward establishing a morphological and ultrastructural characterization of proembryogenic masses and early somatic embryos of Araucaria angustifolia (Bert.) O. Kuntze. Protoplasma. 2016, 253, 487–501. [Google Scholar] [CrossRef]
- Corredoira, E.; Merkle, S.; Martínez, M.; Toribio, M.; Canhoto, J.; Correia, S.; Ballester, A.; Vieitez, A. Non-zygotic embryogenesis in hardwood species. Crit. Rev. Plant Sci. 2019, 38, 29–97. [Google Scholar] [CrossRef]
- Gaj, M. Factors influencing somatic embryogenesis induction and plant regeneration with particular reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 2004, 43, 27–47. [Google Scholar] [CrossRef]
- Fehér, A. Somatic embryogenesis: Stress-induced plant cell fate remodeling. biochemistry Biography. Bioenergy Act 2015, 1849, 385–402. [Google Scholar] [CrossRef]
- Fehér, A. The initiation phase of somatic embryogenesis: What we know and what we don’t. Act Biol. 2008, 52, 53–56. Available online: https://abs.bibl.u-szeged.hu/index.php/abs/article/view/2579 (accessed on 13 April 2025).
- Su, Y.H.; Tang, L.; Zhao, X.; Zhang, X. Plant cell totipotency: Insights into cellular reprogramming. J. Integr. Plant Biol. 2021, 63, 228–240. [Google Scholar] [CrossRef] [PubMed]
- George, E.F.; Hall, M.; Klerk, G. Plant Growth Regulators I: Introduction; Auxins, their analogues and Inhibitors. In Plant Propagation by Tissue Culture; George, E.F., Hall, M., Klerk, G., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 175–204. [Google Scholar] [CrossRef]
- Wójcik, A.M.; Wójcikowska, B.; Gaj, M. Current Perspectives on the Auxin-Mediated Genetic Network that Controls the Induction of Somatic Embryogenesis in Plants. Int. J. Mol. Sci. 2020, 21, 1333. [Google Scholar] [CrossRef]
- Merkle, S.A. Strategies to address the limitations of somatic embryogenesis in hardwood trees. Plant Tissue Cult. Biotechnol. 1995, 31, 6. [Google Scholar]
- Ballester, A.; Corredoira, E.; Vieitez, A. Limitations of somatic embryogenesis in hardwood trees. In Vegetative Propagation of Forest Trees; Park, Y., Bonga, J., Moon, H., Eds.; National Institute of Forest Sciences: Seoul, Republic of Korea, 2016; pp. 56–74. Available online: http://hdl.handle.net/10261/218291 (accessed on 13 April 2025).
- Goeten, D.; Rogge-Renner, G.; Schmidt, E.; Bouzon, Z.; Farias-Soares, F.; Guerra, M.P.; Steiner, N. Updating embryonic ontogenesis in Araucaria angustifolia: From Burlingame (1915) to the present. Protoplasma. 2020, 257, 931–948. [Google Scholar] [CrossRef]
- Stefenon, V.; Steiner, N.; Guerra, M.P.; Nodari, R. Integrating approaches towards the conservation of forest genetic resources: A case study of Araucaria angustifolia. Biodivers. Conserv. 2009, 18, 2433–2448. [Google Scholar] [CrossRef]
- Astarita, L.; Guerra, M.P.; Silveira, V.; Santos, A.W.; Nodari, R. Somatic embryogenesis in Araucaria angustifolia (Bert) O. Kuntze. In Somatic Embryogenesis in Woody Plants; Jain, S., Gupta, P., Newton, R., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; Volume 6, pp. 457–478. [Google Scholar] [CrossRef]
- Silveira, V.; Steiner, N.; dos Santos, A.; Nodari, R.; Guerra, M.P. Biotechnology tolls in Araucaria angustifolia conservation and improvement: Inductive factors affecting Somatic embryogenesis. Crop Breed. Appl. Biotechnol. 2002, 2, 463–470. [Google Scholar] [CrossRef]
- Ma, X.; Bucalo, K.; Determann, R.O.; Cruse-Sanders, J.; Pullman, G. Somatic embryogenesis, plant regeneration, and cryopreservation for Torreya taxifolia, a highly endangered coniferous species. Vitr. Cell Dev. Biol. Plant 2012, 48, 324–334. [Google Scholar] [CrossRef]
- Khuri, S.; Shmoury, M.; Baalbaki, R.; Maunder, M.; Talhouk, S. Conservation of the Cedrus libani populations in Lebanon: History, current status and experimental application of somatic embryogenesis. Biodivers. Conserv. 2000, 9, 1261–1273. [Google Scholar] [CrossRef]
- von Arnold, S.; Egertsdotter, U.; Ekberg, I.; Gupta, P.; Mo, H.; Nörgaard, J. Somatic embryogenesis in Norway spruce (Picea abies). In Somatic Embryogenesis in Woody Plants; Springer: Dordrecht, The Netherlands, 1995; Volume 3, pp. 17–36. [Google Scholar]
- Gupta, P.K.; Pullman, G. Method for Reproducing Coniferous Plants by Somatic Embryogenesis Using Adsorbent Materials in the Development Stage Media. U.S. Patent No. 5034326, 23 July 1991. [Google Scholar]
- Gupta, P.K.; Pullman, G.; Timmis, R.; Kreitinger, M.; Carlson, W.; Grob, J.; Welty, E. Forestry in the 21st century. The biotechnology of somatic embryogenesis. Biotechnology 1993, 11, 454–459. [Google Scholar] [CrossRef]
- Pullman, G.; Webb, D. An embryo staging system for comparison of zygotic and somatic embryo development. In TAPPI R&D Division Biological Sciences Symposium; Springer: Minneapolis, MN, USA, 1994; pp. 31–34. [Google Scholar]
- Pullman, G. Embryogenic tissue initiation in loblolly pine (Pinus taeda L.). Step Wise Protoc. Somat. Embryog. Important Woody Plants 2018, 1, 13–31. [Google Scholar] [CrossRef]
- Gupta, P.K.; Durzan, D. Shoot multiplication from mature trees of Douglas-fir (Pseudotsuga menziesii) and sugar pine (Pinus-lambertiana). Plant Cell Rep. 1985, 4, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Becwar, M.; Blush, T.; Brown, D.; Chesick, E. Multiple paternal genotypes in embryogenic tissue derived from individual immature loblolly pine seeds. Plant Cell Tissue Organ Cult. 1991, 26, 37–44. [Google Scholar] [CrossRef]
- Lelu, M.; Bastien, C.; Drugeault, A.; Gouez, M.; Klimaszewska, K. Somatic embryogenesis and plantlet development in Pinus sylvestris and Pinus pinaster on medium with and without growth regulators. Physiol. Plant. 1999, 105, 719–728. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Smith, D. Maturation of somatic embryos of Pinus strobus is promoted by a high concentration of gellan gum. Physiol. Plant. 1997, 100, 949–957. [Google Scholar] [CrossRef]
- Astarita, L.; Guerra, M.P. Early somatic embryogenesis in Araucaria angustifolia: Induction and maintenance of embryonal-suspensor mass cultures. Braz. J. Plant Physiol. 1998, 10, 113–118. [Google Scholar]
- Haberlandt, G. Culturversuche mit isolierten Pflanzenzellen. In Plant Tissue Culture; Laimer, M., Rücker, W., Eds.; Springer: Vienna, Austria, 2003; Volume 111, pp. 69–91. [Google Scholar] [CrossRef]
- Huihui, G.; Li, Z.; Haixia, G.; Xiwang, C.; Yupeng, F.; Tongtong, L.; Xiushan, Q.; Tongdi, Y.; Aiyun, C.; Fengjuan, S.; et al. Single-cell transcriptome atlas reveals somatic cell embryogenic differentiation features during regeneration. Plant Physiol. 2024, 195, 1414–1431. [Google Scholar] [CrossRef]
- Park, Y.; Pond, S.; Bonga, J. Initiation of somatic embryogenesis in white spruce (Picea glauca): Genetic control, culture treatment effects, and implications for tree breeding. Theor. Appl. Genet. 1993, 86, 427–436. [Google Scholar] [CrossRef]
- Dong, J.; Dunstan, D. Growth parameters, protein and DNA synthesis of an embryogenic suspension culture of white spruce (Picea glauca). J. Plant Physiol. 1994, 144, 201–208. [Google Scholar] [CrossRef]
- Dong, J.; Dunstan, D. Cloning and characterization of six embryogenesis-associated cDNAs from somatic embryos of Picea glauca and their comparative expression during zygotic embryogenesis. Plant Mol. Biol. 1999, 39, 859–864. [Google Scholar] [CrossRef]
- dos Santos, A.; Silveira, V.; Steiner, N.; Vidor, M.; Guerra, M.P. Somatic embryogenesis in Parana pine (Araucaria angustifolia (Bert.) O. Kuntze). Braz. Arch. Biol. Technol. 2002, 45, 97–106. [Google Scholar] [CrossRef]
- Filonova, L.; Bozkov, P.; von Arnold, S. Developmental pathway of somatic embryogenesis in Picea abies as revealed by time-lapse tracking. J. Exp. Bot. 2000, 51, 249–264. [Google Scholar] [CrossRef] [PubMed]
- Farias-Soares, F.; Steiner, N.; Schmidt, E.; Pereira, M.; Rogge-Renner, G.; Bouzon, Z.; Floh, E.; Guerra, M.P. The transition of proembryogenic masses to somatic embryos in Araucaria angustifolia (Bertol.) Kuntze is related to the endogenous contents of IAA, ABA and polyamines. Acta Physiol. Plant. 2014, 36, 1853–1865. [Google Scholar] [CrossRef]
- Singh, H. Embryology of Gimnosperms. Encyclopedia of Plant Anatomy; Gebrüder Borntraeger: Berlin, Germany, 1978. [Google Scholar]
- Attree, S.; Fowke, L. Embryogeny of gymnosperms: Advances in synthetic seed technology of conifers. Plant Cell Tissue Organ Cult. 1993, 35, 1–35. [Google Scholar] [CrossRef]
- Bozhkov, P.V.; Von Arnold, S. Polyethylene glycol promotes maturation but inhibits further development of Picea abies somatic embryos. Physiol. Plant. 1998, 104, 211–224. [Google Scholar] [CrossRef]
- Hristoforoglu, K.; Schmidt, J.; Bolhar-Nordenkampf, H. Development and germination of Abies alba somatic embryos. Plant Cell Tissue Organ Cult. 1995, 40, 277–284. [Google Scholar] [CrossRef]
- Li, X.; Huang, F.; Murphy, J.; Gbur, E. Polyethylene glycol and maltose enhance somatic embryo maturation in loblolly pine (Pinus taeda L.). In Vitro Cell. Dev. Biol.-Plant 1998, 34, 22–26. [Google Scholar] [CrossRef]
- Salajova, T.; Salaj, J.; Kormutak, A. Initiation of embryogenic tissues and plantlet regeneration from somatic embryos of Pinus nigra Arn. Plant Sci. 1999, 145, 33–40. [Google Scholar] [CrossRef]
- Li, X.; Huang, F.; Gbur, E. Polyethylene glycol promoted development of somatic embryos in loblolly pine (Pinus taeda L.). In Vitro Cell. Dev. Biol. -Plant 1997, 33, 184–189. [Google Scholar] [CrossRef]
- von Arnold, S.; Bozhkov, P.; Clapham, D.; Dyachok, J.; Filonova, L.; Högberg, K.; Ingouff, M.; Wiweger, M. Propagation of Norway spruce via somatic embryogenesis. Plant Cell Tissue Organ Cult. 2005, 81, 323–329. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. America and Chile Geographical Location | Plant Population Provenance | Zygotic Embryo Development Stage (Months) | ||
|---|---|---|---|---|
| Pro-Embryo (November) | Early (December) | Late (January) | ||
 | Malalcahuello 38°26’ S –  71 °31’ W 71 °31’ W |  |  |  |
Trongol 37°37’ S – 37°37’ S – 73°07’ W | 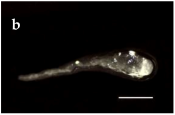 |  | 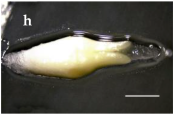 | |
Villa Las  Araucarias Araucarias38°29’ S – 73°14’ W |  |  | 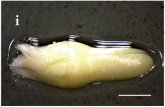 Scale: 1.0 mm | |
| Plant Population Provenance | Pro- Embryo (%) | Early (%) | Late (%) | Pro- Embryo (%) | Early (%) | Late (%) | Pro- Embryo (%) | Early (%) | Late (%) |
|---|---|---|---|---|---|---|---|---|---|
| IC | MIP | MIN | |||||||
| Malalca huello | 6 Bb * | 3 Cb | 0 Cc | 0 Cc | 39 Aa | 0 Cc | 0 Cc | 3 Cb | 0 Cc |
| Trongol | 0 Cc | 17 Bb | 0 Cc | 0 Cc | 22 Ba | 0 Cc | 0 Cc | 19 Ba | 0 Cc |
| Villa Las Araucarias | 0 Cc | 11 Bb | 0 Cc | 0 Cc | 75 Aa | 0 Cc | 0 Cc | 17 Bb | 0 Cc |
| Morpho-Cytological Response of PEMs in Pre-Maturation and Maturation | ||
|---|---|---|
| Step 1 (PGR-free-42 days) | Step 2 (ABA-21 days) | PEMs Cytology (ABA-21 days) |
 | 34 μM ABA | |
 | 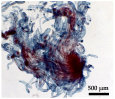 | |
 | 68 μM ABA | |
 |  | |
 | 95 μM ABA | |
 |  | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riffo-Agurto, D.; Steiner, N.; Cartes, P.; Quiroga, P.; Espejo, J.; San Martin, E.; Lasserre, J.-P.; Martínez-Montero, M.E.; Hernández de la Torre, M.; Ríos-Leal, D.; et al. Induction of Somatic Embryogenesis in Araucaria araucana (Molina) K. Koch: Considerations for Ex Situ Conservation of Ancient Tree in Chile. Forests 2025, 16, 732. https://doi.org/10.3390/f16050732
Riffo-Agurto D, Steiner N, Cartes P, Quiroga P, Espejo J, San Martin E, Lasserre J-P, Martínez-Montero ME, Hernández de la Torre M, Ríos-Leal D, et al. Induction of Somatic Embryogenesis in Araucaria araucana (Molina) K. Koch: Considerations for Ex Situ Conservation of Ancient Tree in Chile. Forests. 2025; 16(5):732. https://doi.org/10.3390/f16050732
Chicago/Turabian StyleRiffo-Agurto, Daniela, Neusa Steiner, Priscila Cartes, Pamela Quiroga, Jaime Espejo, Ester San Martin, Jean-Pierre Lasserre, Marcos Edel Martínez-Montero, Martha Hernández de la Torre, Darcy Ríos-Leal, and et al. 2025. "Induction of Somatic Embryogenesis in Araucaria araucana (Molina) K. Koch: Considerations for Ex Situ Conservation of Ancient Tree in Chile" Forests 16, no. 5: 732. https://doi.org/10.3390/f16050732
APA StyleRiffo-Agurto, D., Steiner, N., Cartes, P., Quiroga, P., Espejo, J., San Martin, E., Lasserre, J.-P., Martínez-Montero, M. E., Hernández de la Torre, M., Ríos-Leal, D., Ipinza, R., Sandoval, S., & Sánchez-Olate, M. (2025). Induction of Somatic Embryogenesis in Araucaria araucana (Molina) K. Koch: Considerations for Ex Situ Conservation of Ancient Tree in Chile. Forests, 16(5), 732. https://doi.org/10.3390/f16050732