

Effects of a New Plant Growth Regulator on Callus Induction from Immature Embryo Explants of Korean Pine (Pinus koraiensis)

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Basal Medium
2.3. Plant Growth Regulators
2.4. Explant Inoculation and Induction of Embryogenic Callus
2.5. Data Analysis
3. Results
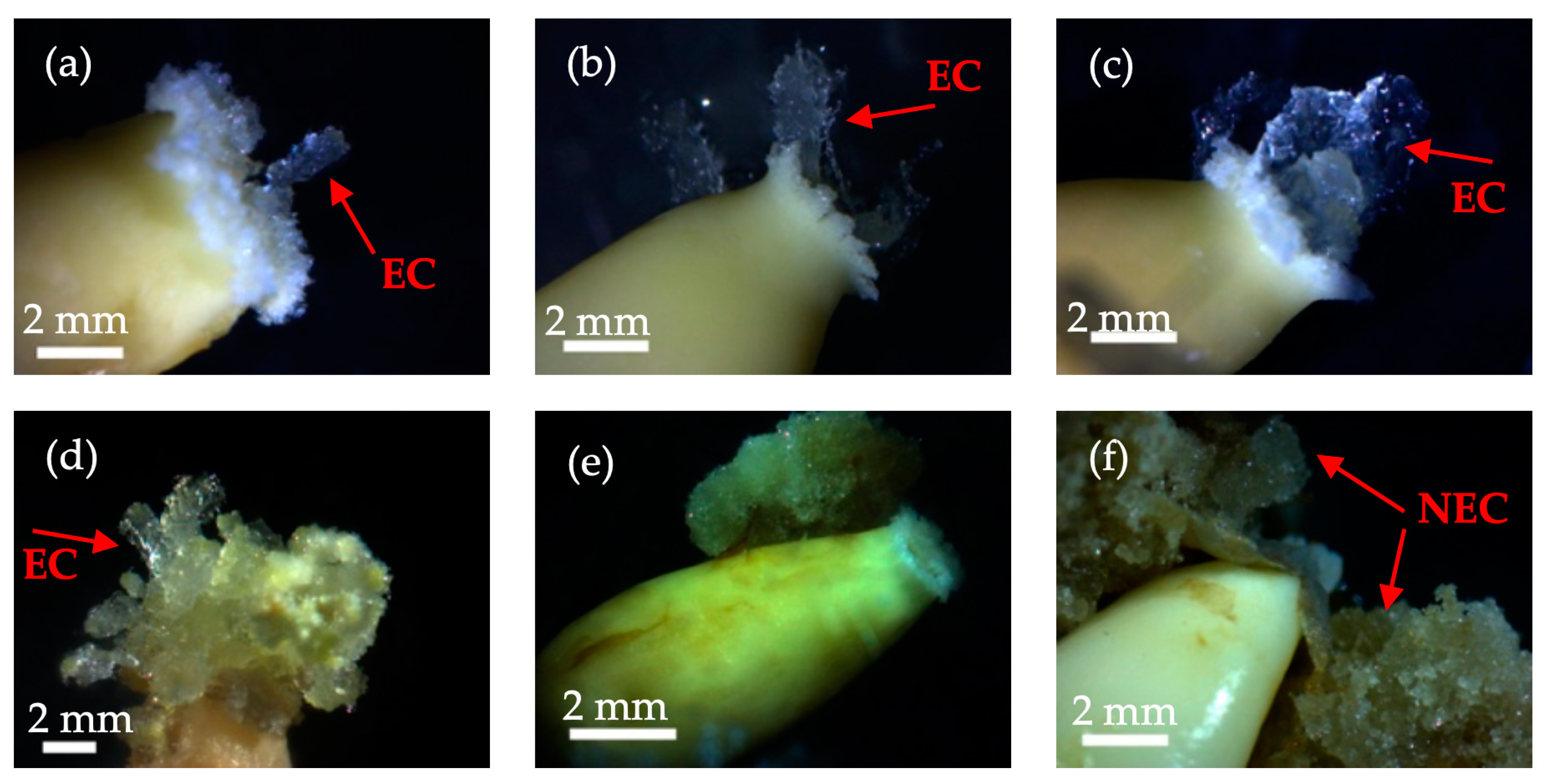
3.1. Induction and Development Process of Embryogenic Callus of Korean Pine
3.2. Impact of Brassinolide on Embryogenic Callus Induction of Different Genotypes of Korean Pine
3.3. Impact of Melatonin on Embryogenic Callus Induction of Different Genotypes of Korean Pine
3.4. Impact of Glutathione on Embryogenic Callus Induction of Different Genotypes of Korean Pine
3.5. Impact of Salicylic Acid on Embryogenic Callus Induction of Different Genotypes of Korean Pine
3.6. Impact of Ascorbic Acid on Embryogenic Callus Induction of Different Genotypes of Korean Pine
3.7. The Effects of Five Inducing Agents on the Induction of Callus Tissues from Different Genotypes of Korean Pine
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nørgaard, J.V.; Duran, V.; Johnsen, Ø.; Krogstrup, P.; Baldursson, S.; Arnold, S.V. Variations in Cryotolerance of Embryogenic Picea abies Cell Lines and the Association to Genetic, Morphological, and Physiological Factors. Can. J. For. Res. 1993, 23, 2560–2567. [Google Scholar] [CrossRef]
- Attree, S.M.; Fowke, L.C. Embryogeny of Gymnosperms: Advances in Synthetic Seed Technology of Conifers. Plant Cell Tissue Organ. Cult. 1993, 35, 1–35. [Google Scholar] [CrossRef]
- Bozhkov, P.V.; Ahn, I.S.; Park, Y.G. Two Alternative Pathways of Somatic Embryo Origin from Polyembryonic Mature Stored Seeds of Pinus koraiensis Sieb et Zucc. Can. J. Bot. 1997, 75, 509–512. [Google Scholar] [CrossRef]
- Peng, C.; Gao, F.; Wang, H.; Shen, H.; Yang, L. Optimization of Maturation Process for Somatic Embryo Production and Cryopreservation of Embryogenic Tissue in Pinus koraiensis. Plant Cell Tiss. Organ. Cult. 2021, 144, 185–194. [Google Scholar] [CrossRef]
- Wu, X.L.; Han, C.; Sun, Y.H.; Cao, S.; Hu, R.Y.; Xu, J.i.L.; Zheng, H.Q.; Li, Y. Optimization of induction conditions for embryogenic callus of somatic embryogenesis in Cunninghamia lanceolata. J. Beijing For. Univ. 2020, 42, 79–86. [Google Scholar] [CrossRef]
- Han, D.Y.; Li, D.; Zhao, J.; Li, H.; Zhang, J.F. Factors Affecting Induction of Embryogenic Callus of Larix principis-rupprechtii. For. Res. 2013, 26, 454–458. [Google Scholar] [CrossRef]
- Dong, L.F.; Xiao, Y.; Shao, C.B. The Effect of Form and Concentration of Nitrogen, Phosphorus, Potassium on Inducing Embryos of Chinese Pine. J. Northwest For. Univ. 2006, 21, 64–66. [Google Scholar]
- Ali, A.; Zhang, J.; Zhou, M.; Chen, T.; Shah, L.; Rehman, S.U.; Hayat, S.; Shi, J.; Chen, J. Chitosan Oligosaccharides Stimulate the Efficacy of Somatic Embryogenesis in Different Genotypes of the Liriodendron Hybrid. Forests 2021, 12, 557. [Google Scholar] [CrossRef]
- Wei, Z.Y.; Li, J. Brassinosteroids Regulate Root Growth, Development, and Symbiosis. Mol. Plant 2016, 9, 86–100. [Google Scholar] [CrossRef]
- Nazir, S.; Jan, H.; Zaman, G.; Ahmed, N.; Drouet, S.; Hano, C.; Abbasi, B.H. Synergistic Effects of Salicylic Acid and Light Stress on Bioactive Metabolites in Basil Callus Cultures. Biocatal. Agric. Biotechnol. 2021, 37, 102176. [Google Scholar] [CrossRef]
- Gao, F.; Wang, R.; Shi, Y.; Shen, H.; Yang, L. Reactive Oxygen Metabolism in the Proliferation of Korean Pine Embryogenic Callus Cells Promoted by Exogenous GSH. Sci. Rep. 2023, 13, 2218. [Google Scholar] [CrossRef]
- Cheng, X.; Xie, T.Y.; Yang, L.; Shen, H.L. Effects of Ascorbic Acid on Physiological Characteristics during Somatic Embryogenesis of Fraxinus mandshurica. Int. J. Mol. Sci. 2022, 24, 289. [Google Scholar] [CrossRef]
- Litvay, J.D.; Verma, D.C.; Johnson, M.A. Influence of a Loblolly Pine (Pinus taeda L.). Culture Medium and Its Components on Growth and Somatic Embryogenesis of the Wild Carrot (Daucus carota L.). Plant Cell Rep. 1985, 4, 325–328. [Google Scholar] [CrossRef]
- Hargreaves, C.L.; Reeves, C.B.; Find, J.I.; Gough, K.; Josekutty, P.; Skudder, D.B.; van der Maas, S.A.; Sigley, M.R.; Menzies, M.I.; Low, C.B.; et al. Improving Initiation, Genotype Capture, and Family Representation in Somatic Embryogenesis of Pinus Radiata by a Combination of Zygotic Embryo Maturity, Media, and Explant Preparation. Can. J. For. Res. 2009, 39, 1566–1574. [Google Scholar] [CrossRef]
- Cai, D.; Chang, J.; Zhu, H.; Luo, B. Tne Dual Effects of BR in Promoting Calluse Formation and Differentiation in Brassica napus. J. Huazhong Agric. 1989, 8, 10–15. [Google Scholar] [CrossRef]
- Chen, J.C.; Xu, M.D.; Zhao, Y.J. Effect of Epibrassinolide on Cell Differentiation of Arabidopsis thaliana. Acta Phytophysiol. Sin. 1996, 22, 399–403. [Google Scholar]
- He, Y.J.; Xu, R.J.; Huang, D.F. Effect of Epibrassinolide on Induction of Calli from Artemisia lactiflora Leaves. Chin. J. Appl. Environ. Biol. 1900, 3, 95–99. [Google Scholar]
- Lu, Z.; Huang, M.; Ge, D.P.; Yang, Y.H.; Cai, X.N.; Qin, P.; She, J.M. Effect of Brassinolide on Callus Growth and Regeneration in Spartina patens (Poaceae). Plant Cell Tissue Organ. Cult. 2003, 73, 87–89. [Google Scholar] [CrossRef]
- Tian, Y.; Zhao, N.; Wang, M.; Zhou, W.; Guo, J.; Han, C.; Zhou, C.; Wang, W.; Wu, S.; Tang, W.; et al. Integrated Regulation of Periclinal Cell Division by Transcriptional Module of BZR1-SHR in Arabidopsis Roots. New Phytol. 2022, 233, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H. Brassinolide Promotes Adventitious Shoot Regeneration from Cauliflower Hypocotyl Segments. Plant Cell Tissue Organ. Cult. 2002, 71, 111–116. [Google Scholar] [CrossRef]
- Yamagishi, Y.; Yoshimoto, J.; Uchiyama, H.; Nabeshima, E.; Nakaba, S.; Watanabe, U.; Funada, R. In Vitro Induction of Secondary Xylem-like Tracheary Elements in Calli of Hybrid Poplar (Populus sieboldii × P. grandidentata). Planta 2013, 237, 1179–1185. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.F.; Liu, W.C.; Wei, L.; Chen, J.N.; Zhang, C.N.; Qiu, J.B.; Ding, W.Y.; Zheng, Q.S. Melatonin Regulates Photosynthesis and Ascorbate-Glutathione Cycle in a Mangrove Kandelia obovata under Low Temperature Stress. Plant Physiol. J. 2019, 55, 1211–1221. [Google Scholar] [CrossRef]
- Zhou, R.; Cai, Y.; Lin, T.Y.; Chai, M.L. Effects of melatonin and epibrassinolide on the regeneration of long-term subcultured callus of Zoysia matrella (L.) Merr. under simulated drought stress. J. Zhejiang Univ. (Agric. Life Sci.) 2022, 48, 36–44. [Google Scholar] [CrossRef]
- Siamak, S.B.; Parisa, S. Strigolactone and Methyl Jasmonate-Induced Antioxidant Defense and the Composition Alterations of Different Active Compounds in Dracocephalum kotschyi Boiss Under Drought Stress. J. Plant Growth Regul. 2020, 40, 878–889. [Google Scholar] [CrossRef]
- Zhang, Y.; Ya, R.; Xu, W.R.; Wang, J.H.; Cui, Y.; Li, J.D. Role of melatonin in the induction of somatic embryogenesis from seeds of Vitis vinifera ‘Chardonnay’. J. Fruit. Sci. 2021, 38, 922–933. [Google Scholar] [CrossRef]
- do Nascimento Vieira, L.; Santa-Catarina, C.; de Freitas Fraga, H.P.; dos Santos, A.L.W.; Steinmacher, D.A.; Schlogl, P.S.; Silveira, V.; Steiner, N.; Floh, E.I.S.; Guerra, M.P. Glutathione Improves Early Somatic Embryogenesis in Araucaria angustifolia (Bert) O. Kuntze by Alteration in Nitric Oxide Emission. Plant Sci. 2012, 195, 80–87. [Google Scholar] [CrossRef]
- Shan, Y.S.; Dai, H.H.; He, X.; Xin, Z.Q.; Wu, N.B. Effects of Exogenous Methyl Jasmonate and Salicylic Acid on Physiological Characteristics and Secondary Metabolism of Atropa belladonna under NaCl Stress. Plant Physiol. J. 2019, 55, 1335–1346. [Google Scholar] [CrossRef]
- Shang, J.; Wu, Z.W.; Ma, Y.G. Phenylpropanoid Metabolism Pathway in Plants. Chin. J. Biochem. Mol. Biol. 2022, 38, 1467–1476. [Google Scholar] [CrossRef]
- Stasolla, C.; Yeung, E.C. Cellular Ascorbic Acid Regulates the Activity of Major Peroxidases in the Apical Poles of Germinating White Spruce (Picea glauca) Somatic Embryos. Plant Physiol. Biochem. 2007, 45, 188–198. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Exogenous Additives | Concentrations | ||||
|---|---|---|---|---|---|
| Brassinolide (mg·L−1) | 0.05 | 0.25 | 0.50 | 1.00 | 2.00 |
| Melatonin (mg·L−1) | 2.50 | 25 | 50 | 100 | 200 |
| Salicylic acid (mg·L−1) | 0.10 | 1.0 | 10 | 50 | 100 |
| Glutathione (g·L−1) | 0.10 | 0.30 | 0.60 | 0.90 | 1.80 |
| Ascorbic acid (g·L−1) | 0.10 | 0.20 | 0.40 | 1.0 | 1.50 |
| Culture Days | Genotype | Concentrations of Brassinolide/mg·L−1 | Control | ||||
|---|---|---|---|---|---|---|---|
| 0.05 | 0.25 | 0.50 | 1.00 | 2.00 | |||
| 10 | I | 0 b | 4.00 ± 0.36 ab | 0 b | 20.00 ± 4.9 a | 4.00 ± 0.36 ab | 12.00 ± 2.15 ab |
| II | 8.00 ± 0.88 a | 12.00 ± 2.15 a | 0 a | 8.00 ± 0.88 a | 12.00 ± 2.15 a | 4.00 ± 0.36 a | |
| III | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a | |
| 15 | I | 4.00 ± 0.36 b | 16.00 ± 2.68 ab | 0 b | 32.00 ± 5.72 a | 0 b | 12.00 ± 2.15 b |
| II | 0 b | 0 b | 0 b | 0 b | 0 b | 9.00 ± 1.12 a | |
| III | 24.00 ± 2.15 a | 20.00 ± 5.66 a | 4.00 ± 0.36 a | 20.00 ± 4 a | 12.00 ± 1.31 a | 4.00 ± 0.36 a | |
| 20 | I | 12.00 ± 2.15 ab | 16.00 ± 5.72 ab | 8.00 ± 0.88 ab | 28.00 ± 3.07 ab | 0 b | 28.00 ± 5.01 a |
| II | 20.00 ± 0 ab | 28.00 ± 6.38 ab | 8.00 ± 1.43 b | 36.00 ± 10.68 a | 12.00 ± 1.31 ab | 16.00 ± 1.43 ab | |
| III | 0 a | 0 a | 15.00 ± 3.35 a | 8.00 ± 1.43 a | 0 a | 5.00 ± 0.56 a | |
| 25 | I | 4.00 ± 0.36 b | 16.00 ± 5.72 ab | 8.00 ± 0.88 ab | 36.00 ± 7.89 a | 0 b | 36.00 ± 7.89 a |
| II | 20.00 ± 2.83 ab | 24.00 ± 6.26 a | 0 b | 12.00 ± 2.15 ab | 8.00 ± 0.88 ab | 20.00 ± 2.83 ab | |
| III | 0 b | 0 b | 25.00 ± 4.42 a | 8.00 ± 1.43 b | 0 b | 0 b | |
| 30 | I | 12.00 ± 1.31 ab | 16.00 ± 5.72 ab | 4.00 ± 0.36 b | 32.00 ± 3.51 a | 0 b | 8.00 ± 1.43 ab |
| II | 20.00 ± 2.83 a | 20.00 ± 4.00 a | 0 a | 12.00 ± 2.15 a | 12.00 ± 2.15 a | 20.00 ± 2.83 a | |
| III | 0 a | 0 a | 0 a | 20.00 ± 4.00 a | 4.00 ± 0.36 a | 0 a | |
| Culture Days | Genotype | Concentrations of Melatonin/mg·L−1 | Control | ||||
|---|---|---|---|---|---|---|---|
| 2.5 | 25 | 50 | 100 | 200 | |||
| 10 | I | 16.00 ± 2.68 a | 20.00 ± 0.01 a | 4.00 ± 0.36 a | 4.00 ± 0.36 a | 4.00 ± 0.36 a | 12.00 ± 2.15 a |
| II | 0 b | 12.00 ± 1.31 a | 0 b | 0 b | 0 b | 4.00 ± 0.36 b | |
| III | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a | |
| 15 | I | 12.00 ± 2.15 a | 16.00 ± 2.68 a | 4.00 ± 0.36 a | 0 a | 0 a | 12.00 ± 2.15 a |
| II | 0 b | 0 b | 0 b | 0 b | 0 b | 9.00 ± 1.12 a | |
| III | 8.00 ± 0.88 b | 12.00 ± 2.15 b | 28.00 ± 3.07 a | 0 b | 4.00 ± 0.36 b | 4.00 ± 0.36 b | |
| 20 | I | 12.00 ± 2.15 ab | 16.00 ± 2.68 ab | 4.00 ± 0.36 b | 0 b | 0 b | 28.00 ± 5.01 a |
| II | 8.00 ± 1.43 a | 12.00 ± 1.31 a | 16.00 ± 2.68 a | 0 a | 0 a | 16.00 ± 1.43 a | |
| III | 4.00 ± 0.36 a | 0 a | 0 a | 0 a | 0 a | 5.00 ± 0.56 a | |
| 25 | I | 20.00 ± 4.9 ab | 16.00 ± 2.68 ab | 4.00 ± 0.36 b | 0 b | 0 b | 36.00 ± 7.89 a |
| II | 8.00 ± 1.43 ab | 8.00 ± 0.88 ab | 8.00 ± 1.43 ab | 0 b | 0 b | 20.00 ± 2.83 a | |
| III | 0 a | 4.00 ± 0.36 a | 0 a | 0 a | 0 a | 0 a | |
| 30 | I | 4.00 ± 0.36 b | 28.00 ± 5.01 a | 0 b | 0 b | 0 b | 8.00 ± 1.43 b |
| II | 8.00 ± 1.43 ab | 8.00 ± 0.88 ab | 8.00 ± 0.88 ab | 0 b | 0 b | 20.00 ± 2.83 a | |
| III | 4.00 ± 0.36 a | 4.00 ± 0.36 a | 0 a | 0 a | 0 a | 0 a | |
| Culture Days | Genotype | Concentrations of Glutathione/g·L−1 | Control | ||||
|---|---|---|---|---|---|---|---|
| 0.10 | 0.30 | 0.60 | 0.90 | 1.80 | |||
| 10 | I | 0 a | 0 a | 8.00 ± 0.88 a | 0 a | 16.00 ± 3.51 a | 12.00 ± 2.15 a |
| II | 0 a | 0 a | 0 a | 0 a | 0 a | 4.00 ± 0.36 a | |
| III | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a | |
| 15 | I | 4.00 ± 0.36 a | 8.00 ± 0.88 a | 8.00 ± 0.88 a | 8.00 ± 0.88 a | 16.00 ± 3.51 a | 12.00 ± 2.15 a |
| II | 0 a | 0 a | 0 a | 4.00 ± 0.36 a | 0 a | 9.00 ± 1.12 a | |
| III | 4.00 ± 0.36 a | 0 a | 8.00 ± 0.88 a | 0 a | 4.00 ± 0.36 a | 4.00 ± 0.36 b | |
| 20 | I | 12.00 ± 2.15 ab | 0 b | 20.00 ± 4.9 ab | 4.00 ± 0.36 b | 12.00 ± 2.15 ab | 28.00 ± 5.01 a |
| II | 4.00 ± 0.36 b | 0 b | 0 b | 4.00 ± 0.36 b | 4.00 ± 0.36 b | 16.00 ± 1.43 a | |
| III | 8.00 ± 0.88 a | 0 a | 0 a | 12.00 ± 2.15 a | 0 a | 5.00 ± 0.56 a | |
| 25 | I | 24.00 ± 6.26 ab | 0 b | 16.00 ± 2.68 ab | 8.00 ± 0.88 b | 12.00 ± 2.15 ab | 36.00 ± 7.89 a |
| II | 0 b | 0 b | 4.00 ± 0.36 b | 4.00 ± 0.36 b | 8.00 ± 1.43 ab | 20.00 ± 2.83 a | |
| III | 8.00 ± 0.88 a | 0 a | 5.00 ± 0.56 a | 0 a | 0 a | 0 a | |
| 30 | I | 20.00 ± 4.9 a | 4.00 ± 0.36 a | 12.00 ± 1.31 a | 4.00 ± 0.36 a | 8.00 ± 0.88 a | 8.00 ± 1.43 b |
| II | 12.00 ± 2.15 a | 4.00 ± 0.36 a | 8.00 ± 1.43 a | 8.00 ± 0.88 a | 4.00 ± 0.36 a | 20.00 ± 2.83 a | |
| III | 4.00 ± 0.36 a | 0 a | 5.00 ± 0.56 a | 4.00 ± 0.36 a | 0 a | 0 a | |
| Culture Days | Genotype | Concentrations of Salicylic Acid/mg·L−1 | Control | ||||
|---|---|---|---|---|---|---|---|
| 0.10 | 1.0 | 10 | 50 | 100 | |||
| 10 | I | 0 b | 4.00 ± 0.36 ab | 0 b | 0 b | 0 b | 12.00 ± 2.15 a |
| II | 4.00 ± 0.36 a | 0 a | 0 a | 0 a | 0 a | 4.00 ± 0.36 b | |
| III | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a | |
| 15 | I | 0 b | 0 b | 0 b | 0 b | 0 b | 12.00 ± 2.15 a |
| II | 0 b | 0 b | 0 b | 0 b | 0 b | 9.00 ± 1.12 a | |
| III | 0 a | 0 a | 0 a | 0 a | 0 a | 4.00 ± 0.36 b | |
| 20 | I | 8.00 ± 0.88 b | 8.00 ± 0.88 b | 8.00 ± 0.88 b | 4.00 ± 0.36 b | 0 b | 28.00 ± 5.01 a |
| II | 0 b | 0 b | 0 b | 0 b | 0 b | 16.00 ± 1.43 a | |
| III | 0 a | 5.00 ± 0.56 a | 0 a | 0 a | 0 a | 5.00 ± 0.56 a | |
| 25 | I | 8.00 ± 0.88 b | 4.00 ± 0.36 b | 0 b | 0 b | 0 b | 36.00 ± 7.89 a |
| II | 4.00 ± 0.36 b | 0 b | 0 b | 0 b | 0 b | 20.00 ± 2.83 a | |
| III | 0 a | 4.00 ± 0.36 a | 0 a | 0 a | 0 a | 0 a | |
| 30 | I | 0 a | 0 a | 0 a | 0 a | 0 a | 8.00 ± 1.43 a |
| II | 4.00 ± 0.36 b | 0 b | 0 b | 0 b | 0 b | 20.00 ± 2.83 a | |
| III | 0 b | 0 b | 0 b | 0 b | 0 b | 0 b | |
| Culture Days | Genotype | Concentrations of Ascorbic Acid/g·L−1 | Control | ||||
|---|---|---|---|---|---|---|---|
| 0.1 | 0.20 | 0.40 | 1.00 | 1.50 | |||
| 10 | I | 8.00 ± 0.88 a | 0 a | 0 a | 0 a | 0 a | 12.00 ± 2.15 a |
| II | 0 a | 0 a | 0 a | 0 a | 0 a | 4.00 ± 0.36 b | |
| III | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a | |
| 15 | I | 8.00 ± 0.88 a | 4.00 ± 0.36 a | 8.00 ± 0.88 a | 0 a | 0 a | 12.00 ± 2.15 a |
| II | 8.00 ± 1.43 a | 8.00 ± 0.88 a | 0 a | 0 a | 0 a | 9.00 ± 1.12 a | |
| III | 4.00 ± 0.36 a | 4.00 ± 0.36 a | 0 a | 0 a | 0 a | 4.00 ± 0.36 b | |
| 20 | I | 0 b | 4.00 ± 0.36 b | 4.00 ± 0.36 b | 4.00 ± 0.36 b | 4.00 ± 0.36 b | 28.00 ± 5.01 a |
| II | 0 b | 8.00 ± 0.88 b | 0 b | 0 b | 0 b | 16.00 ± 1.43 a | |
| III | 0 a | 4.00 ± 0.36 a | 4.00 ± 0.36 a | 0 a | 0 a | 5.00 ± 0.56 a | |
| 25 | I | 4.00 ± 0.36 b | 0 b | 0 b | 4.00 ± 0.36 b | 12.00 ± 1.31 b | 36.00 ± 7.89 a |
| II | 0 b | 12.00 ± 2.15 ab | 4.00 ± 0.36 b | 0 b | 0 b | 20.00 ± 2.83 a | |
| III | 4.00 ± 0.36 a | 0 a | 4.00 ± 0.36 a | 0 a | 0 aa | 0 a | |
| 30 | I | 8.00 ± 1.43 a | 4.00 ± 0.36 a | 12.00 ± 2.15 a | 0 a | 0 a | 8.00 ± 1.43 b |
| II | 4.00 ± 0.36 b | 8.00 ± 1.43 ab | 0 b | 0 b | 0 b | 20.00 ± 2.83 a | |
| III | 4.00 ± 0.36 a | 0 a | 0 a | 0 a | 0 a | 0 a | |
| Exogenous Additives | Concentrations | Genotype I Induction Rate/% | Genotype Ⅱ Induction Rate/% | Genotype Ⅲ Induction Rate/% |
|---|---|---|---|---|
| AsA (g·L-1) | 0.00 | 19.20 ± 3.92 a | 13.80 ± 1.75 a | 1.80 ± 0.11 a |
| 0.10 | 5.60 ± 0.61 b | 2.40 ± 0.21 c | 2.40 ± 0.16 a | |
| 0.20 | 2.40 ± 0.16 b | 7.20 ± 0.92 b | 1.60 ± 0.09 a | |
| 0.40 | 4.80 ± 0.5 b | 0.80 ± 0.03 c | 1.60 ± 0.09 a | |
| 1.00 | 1.60 ± 0.09 b | 0 c | 0 a | |
| 1.50 | 3.20 ± 0.24 b | 0 c | 0 a | |
| BL (mg·L-1) | 0.00 | 19.20 ± 3.92 a | 13.80 ± 1.75 a | 1.80 ± 0.11 a |
| 0.05 | 6.40 ± 0.71 cd | 13.60 ± 1.71 a | 4.80 ± 0.5 ab | |
| 0.25 | 13.60 ± 3.66 bc | 16.80 ± 3.45 a | 4.00 ± 0.57 ab | |
| 0.50 | 4.00 ± 0.33 cd | 1.60 ± 0.13 b | 8.80 ± 1.39 ab | |
| 1.00 | 29.60 ± 5.16 a | 13.60 ± 2.8 a | 11.20 ± 1.95 a | |
| 2.00 | 0.80 ± 0.03 d | 8.80 ± 1.15 ab | 2.40 ± 0.16 b | |
| GSH (g·L-1) | 0.00 | 19.20 ± 3.92 a | 13.80 ± 1.75 a | 1.80 ± 0.11 ab |
| 0.10 | 12.00 ± 2.3 ab | 3.20 ± 0.30 b | 4.80 ± 0.42 a | |
| 0.30 | 2.40 ± 0.16 c | 0.80 ± 0.03 b | 0 b | |
| 0.60 | 12.80 ± 1.94 ab | 2.40 ± 0.21 b | 3.60 ± 0.31 ab | |
| 0.90 | 4.80 ± 0.42 bc | 4.00 ± 0.33 b | 3.20 ± 0.30 ab | |
| 1.80 | 12.80 ± 2.2 ab | 3.20 ± 0.30 b | 0.80 ± 0.03 ab | |
| Mel (mg·L-1) | 0.00 | 19.20 ± 3.92 a | 13.80 ± 1.75 a | 1.80 ± 0.11 ab |
| 2.50 | 12.80 ± 2.2 a | 4.80 ± 0.64 bc | 3.20 ± 0.24 ab | |
| 25.00 | 19.20 ± 2.82 a | 8.00 ± 0.80 b | 4.00 ± 0.40 ab | |
| 50.00 | 3.20 ± 0.24 b | 6.40 ± 0.80 b | 5.60 ± 0.69 a | |
| 100.00 | 0.80 ± 0.03 b | 0 c | 0 b | |
| 200.00 | 0.80 ± 0.03 b | 0 c | 0.80 ± 0.03 ab | |
| SA (mg·L-1) | 0.00 | 19.20 ± 3.92 a | 13.80 ± 1.75 a | 1.80 ± 0.11 a |
| 0.10 | 3.20 ± 0.24 b | 2.40 ± 0.16 b | 0 a | |
| 1.00 | 3.20 ± 0.24 b | 0 b | 1.80 ± 0.11 a | |
| 10.00 | 1.60 ± 0.09 b | 0 b | 0 a | |
| 50.00 | 0.80 ± 0.03 b | 0 b | 0 a | |
| 100.00 | 0 b | 0 b | 0 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nie, S.; Wang, Y.; Yan, Y.; Liu, S.; Guo, W.; Yang, L.; Shen, H. Effects of a New Plant Growth Regulator on Callus Induction from Immature Embryo Explants of Korean Pine (Pinus koraiensis). Forests 2023, 14, 2413. https://doi.org/10.3390/f14122413
Nie S, Wang Y, Yan Y, Liu S, Guo W, Yang L, Shen H. Effects of a New Plant Growth Regulator on Callus Induction from Immature Embryo Explants of Korean Pine (Pinus koraiensis). Forests. 2023; 14(12):2413. https://doi.org/10.3390/f14122413
Chicago/Turabian StyleNie, Shuai, Yue Wang, Yong Yan, Shanshan Liu, Wenhui Guo, Ling Yang, and Hailong Shen. 2023. "Effects of a New Plant Growth Regulator on Callus Induction from Immature Embryo Explants of Korean Pine (Pinus koraiensis)" Forests 14, no. 12: 2413. https://doi.org/10.3390/f14122413
APA StyleNie, S., Wang, Y., Yan, Y., Liu, S., Guo, W., Yang, L., & Shen, H. (2023). Effects of a New Plant Growth Regulator on Callus Induction from Immature Embryo Explants of Korean Pine (Pinus koraiensis). Forests, 14(12), 2413. https://doi.org/10.3390/f14122413