
Different Influences of Soil and Climatic Factors on Shrubs and Herbaceous Plants in the Shrub-Encroached Grasslands of the Mongolian Plateau

 ,
,
Abstract
1. Introduction
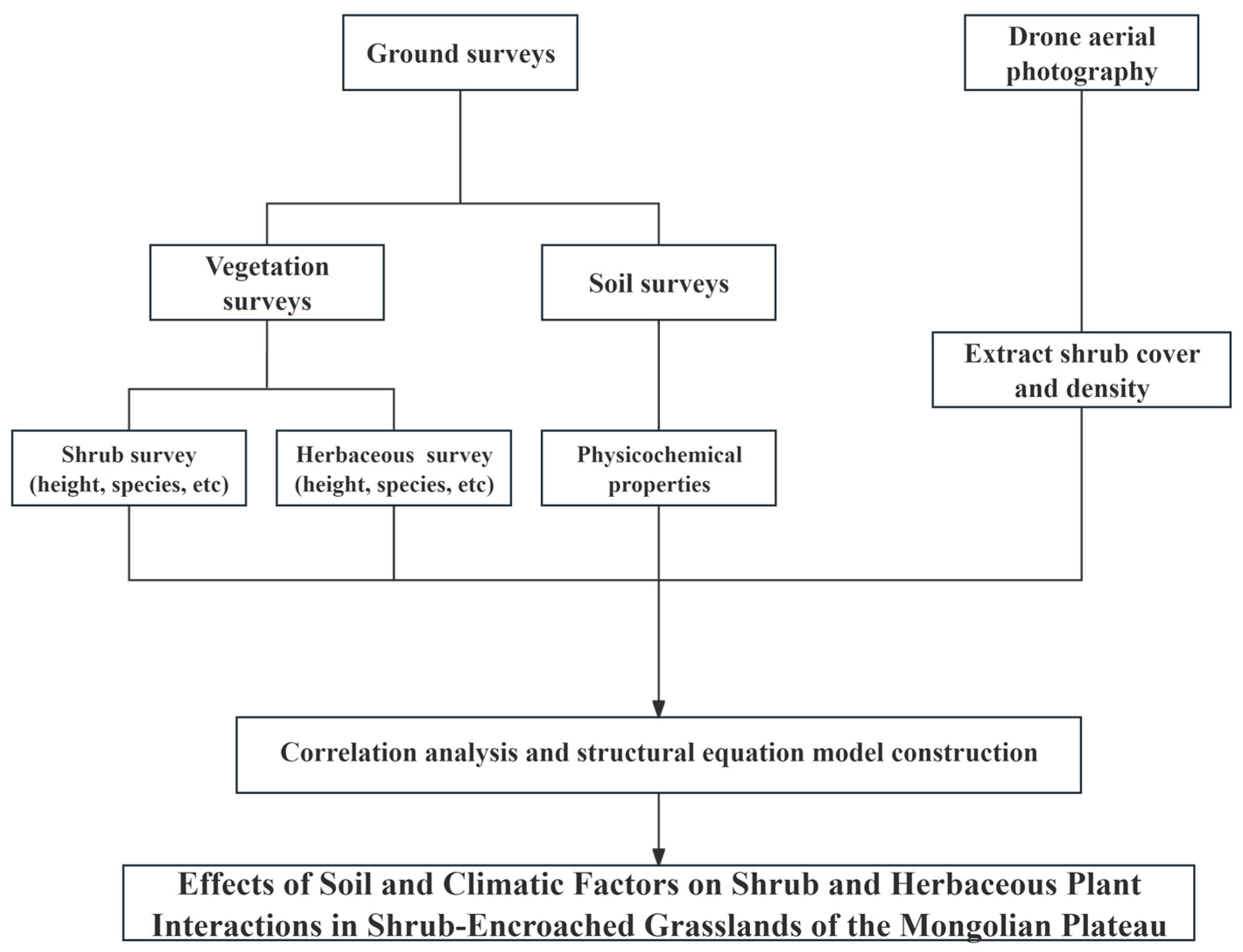
2. Materials and Methods
2.1. Overview of the Study Area
2.2. Plot Establishment and Survey
2.3. Soil Analysis
2.4. Climate Data Acquisition
2.5. Acquisition of Shrub Cover Data
2.6. Other Data Processing
3. Results
3.1. Basic Information on Caragana tibetica Communities and Soil Particle Size Characteristics
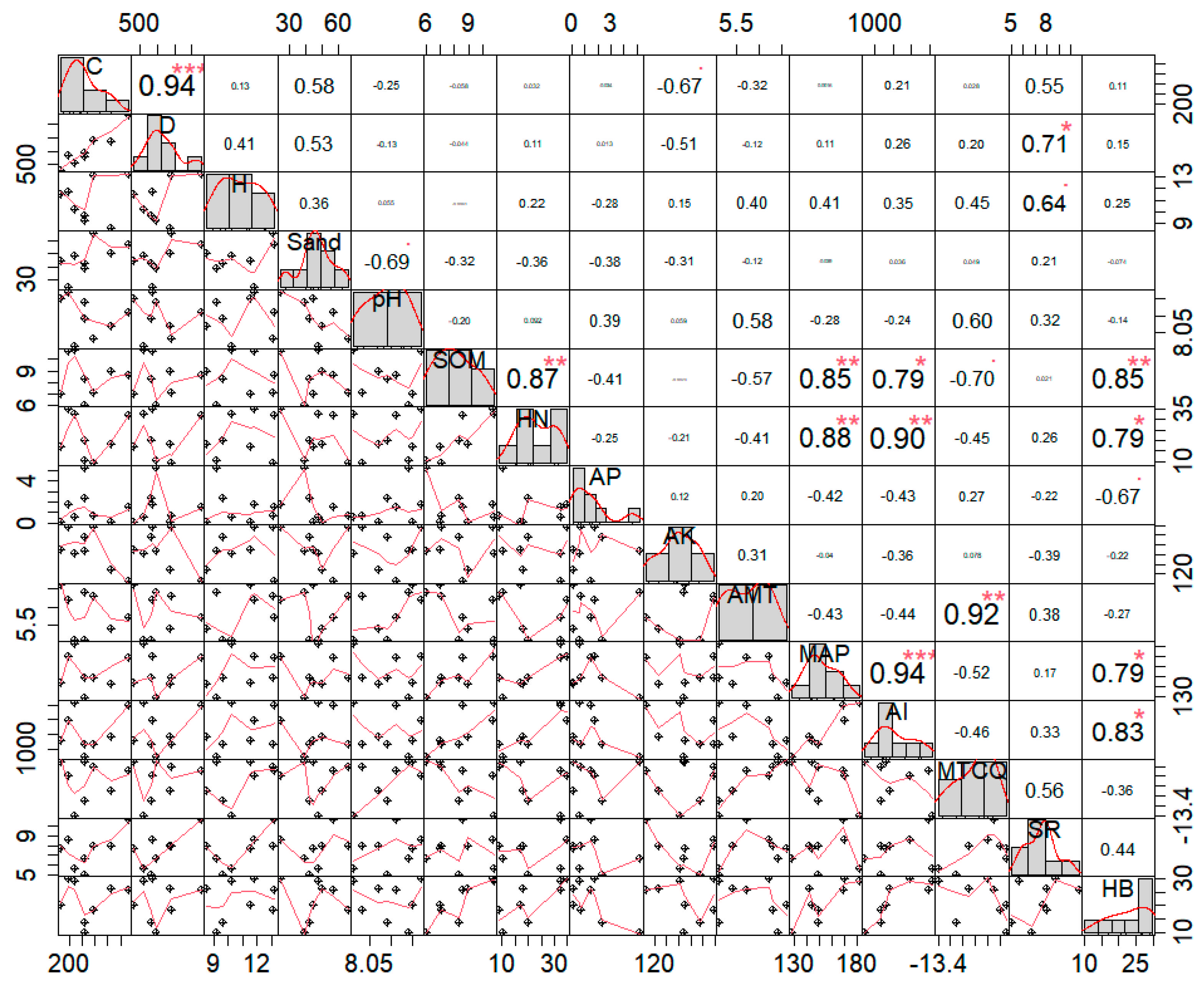
3.2. Correlation Analysis of Caragana Tibetica Plant Communities and Their Relationships with Environmental Factors
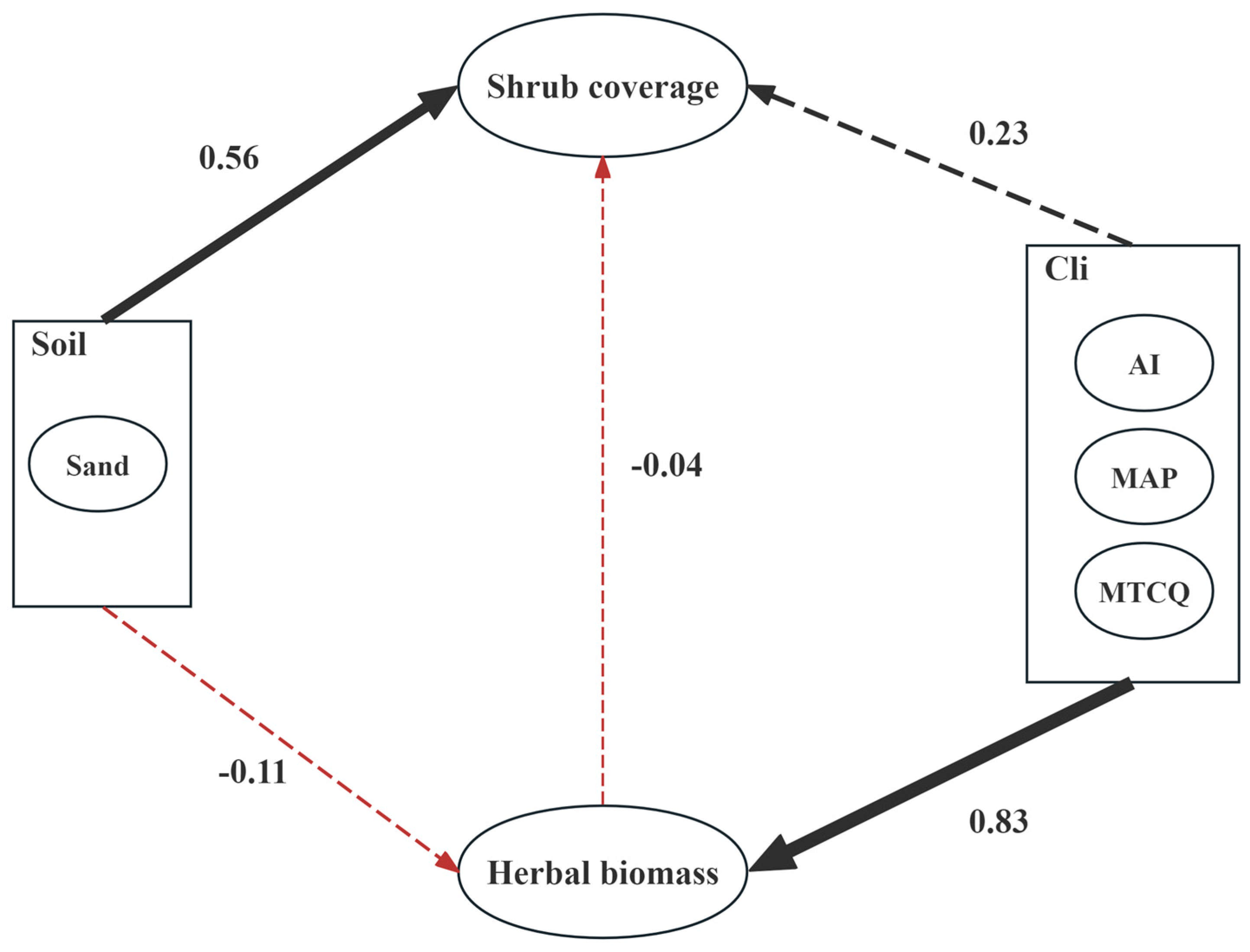
3.3. Analysis of Factors Influencing Caragana tibetica Communities
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, H.; Shen, H.; Zhou, L.; Zhu, Y.; Chen, L.; Hu, H.; Zhang, P.; Fang, J. Shrub encroachment increases soil carbon and nitrogen stocks in temperate grasslands in China. Land Degrad. Dev. 2019, 30, 756–767. [Google Scholar] [CrossRef]
- D’Odorico, P.; Okin, G.S.; Bestelmeyer, B.T. A synthetic review of feedbacks and drivers of shrub encroachment in arid grasslands. Ecohydrology 2012, 5, 520–530. [Google Scholar] [CrossRef]
- Sala Osvaldo, E.; Maestre Fernando, T. Grass–woodland transitions: Determinants and consequences for ecosystem functioning and provisioning of services. J. Ecol. 2014, 102, 1357–1362. [Google Scholar] [CrossRef]
- Van Auken, O.W. Shrub invasions of North American semiarid grasslands. Annu. Rev. Ecol. Syst. 2000, 31, 197–215. [Google Scholar] [CrossRef]
- Peng, H.Y.; Li, X.Y.; Li, G.Y.; Zhang, Z.H.; Zhang, S.Y.; Li, L.; Zhao, G.Q.; Jiang, Z.Y.; Ma, Y.J. Shrub encroachment with increasing anthropogenic disturbance in the semiarid Inner Mongolian grasslands of China. Catena 2013, 109, 39–48. [Google Scholar] [CrossRef]
- Urbina, I.; Grau, O.; Sardans, J.; Ninot, J.M.; Peñuelas, J. Encroachment of shrubs into subalpine grasslands in the Pyrenees changes the plant-soil stoichiometry spectrum. Plant Soil 2020, 448, 37–53. [Google Scholar] [CrossRef]
- Knapp, A.K.; Briggs, J.M.; Collins, S.L.; Archer, S.R.; BRET-HARTE, M.S.; Ewers, B.E.; Peters, D.P.; Young, D.R.; Shaver, G.R.; Pendall, E.; et al. Shrub encroachment in North American grasslands: Shifts in growth form dominance rapidly alters control of ecosystem carbon inputs. Glob. Change Biol. 2008, 14, 615–623. [Google Scholar] [CrossRef]
- Shackleton, C.M.; Scholes, R.J. Above ground woody community attributes, biomass and carbon stocks along a rainfall gradient in the savannas of the central lowveld, South Africa. S. Afr. J. Bot. 2010, 77, 184–192. [Google Scholar] [CrossRef]
- Cai, Y.; Yan, Y.; Xu, D.; Xu, X.; Wang, C.; Wang, X.; Chen, J.; Xin, X.; Eldridge, D.J. The fertile island effect collapses under extreme overgrazing: Evidence from a shrub-encroached grassland. Plant Soil 2020, 448, 201–212. [Google Scholar] [CrossRef]
- Bond William, J.; Midgley Jeremy, J. Ecology of sprouting in woody plants: The persistence niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Lou, S.; Hu, A.; Ren, J.; Hu, J.; Zhang, J.; Hou, F. Shrub Encroachment in Grasslands: Processes, Mechanisms, and Effects. Acta Prataculturae Sin. 2018, 27, 219–227. [Google Scholar]
- Miao, L.; Jiang, C.; Xue, B.; Liu, Q.; He, B.; Nath, R.; Cui, X. Vegetation dynamics and factor analysis in arid and semi-arid Inner Mongolia. Environ. Earth Sci. 2015, 73, 2343–2352. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Li, X.Y.; Yang, X.; Shi, Y.; Zhang, S.Y.; Jiang, Z.Y. Changes in soil properties following shrub encroachment in the semiarid Inner Mongolian grasslands of China. Soil Sci. Plant Nutr. 2020, 66, 369–378. [Google Scholar] [CrossRef]
- Shen, X.; Liu, Y.; Liu, B.; Zhang, J.; Wang, L.; Lu, X.; Jiang, M. Effect of shrub encroachment on land surface temperature in semi-arid areas of temperate regions of the Northern Hemisphere. Agric. For. Meteorol. 2022, 320, 108943. [Google Scholar] [CrossRef]
- Du, Z.; Zheng, H.; Penuelas, J.; Sardans, J.; Deng, D.; Cai, X.; Gao, D.; Nie, S.; He, Y.; Lü, X.; et al. Shrub encroachment leads to accumulation of C, N, and P in grassland soils and alters C: N: P stoichiometry: A meta-analysis. Sci. Total Environ. 2024, 951, 175534. [Google Scholar] [CrossRef]
- Wen, Y.Y.; Zhu, J.; Wang, H.; Zhang, M.D.; Lu, S.B.; Zheng, S.X. Population characteristics of Caragana microphylla and the influencing soil factors in shrub-encroached grassland of Inner Mongolia, China. Ying Yong Sheng Tai Xue Bao = J. Appl. Ecol. 2024, 35, 1525–1533. [Google Scholar]
- Niu, F.; Pierce, N.A.; Okin, G.S.; Archer, S.R.; Fischella, M.R.; Nadoum, S. Sandblasting promotes shrub encroachment in arid grasslands. New Phytol. 2023, 240, 1817–1829. [Google Scholar] [CrossRef]
- Xie, L.; Li, Y.; Guo, H.; Wang, C.; Chen, Q.; He, P.; Ma, C. Sandy habitats play an important role in shrub encroachment in grasslands. Agronomy 2022, 12, 2858. [Google Scholar] [CrossRef]
- Allington, G.R.; Valone, T.J. Islands of fertility: A byproduct of grazing. Ecosystems 2014, 17, 127–141. [Google Scholar] [CrossRef]
- Good, S.P.; Caylor, K.K. Climatological determinants of woody cover in Africa. Proc. Natl. Acad. Sci. USA 2011, 108, 4902–4907. [Google Scholar] [CrossRef]
- Weber-Grullon, L.; Gherardi, L.; Rutherford, W.A.; Archer, S.R.; Sala, O.E. Woody-plant encroachment: Precipitation, herbivory and grass-competition interact to affect shrub recruitment. Ecol. Appl. 2022, 32, e2536. [Google Scholar] [CrossRef]
- Pei, Y.; Huang, L.; Shao, M.; Li, R.; Zhang, Y. Characteristics and Influencing Factors of Soil Water Recharge Under Different Groundwater Table Depths in the Mu Us Sandy Land. Trans. Chin. Soc. Agric. Eng. 2021, 37, 108–116. [Google Scholar]
- Zhang, Z.Y.; Gao, G.Q.; Xu, X.L.; Yu, M.; Qiang, T.Y. Shrubs proliferated within a six-year exclosure in a temperate grassland-spatiotemporal relationships between vegetation and soil variables. Sci. Cold Arid Reg. 2014, 6, 139–149. [Google Scholar]
- Liu, X.; Zhuang, Q.; Lai, L.; Zhou, J.; Sun, Q.; Yi, S.; Liu, B.; Zheng, Y. Soil water use sources and patterns in shrub encroachment in semiarid grasslands of Inner Mongolia. Agric. For. Meteorol. 2021, 308, 108579. [Google Scholar] [CrossRef]
- World Reference Base for Soil Resources. World Reference Base for Soil Resources 2022: International Soil Classification and Correlation System; FAO: Rome, Italy, 2022. [Google Scholar]
- Song, Y.; Guo, Z.; Lu, Y.; Yan, D.; Liao, Z.; Liu, H.; Cui, Y. Pixel-level spatiotemporal analyses of vegetation fractional coverage variation and its influential factors in a desert steppe: A case study in Inner Mongolia, China. Water 2017, 9, 478. [Google Scholar] [CrossRef]
- Li, C.; Hao, X.; Zhao, M.; Han, G.; Willms, W.D. Influence of historic sheep grazing on vegetation and soil properties of a Desert Steppe in Inner Mongolia. Agric. Ecosyst. Environ. 2008, 128, 109–116. [Google Scholar] [CrossRef]
- Guo, K.; Liu, C.C.; Xie, Z.Q.; Li, Y.; Lu, Z.J.; Ma, K.P. China Vegetation Classification: Concept, approach and applications. Phytocoenologia 2018, 48, 113–120. [Google Scholar] [CrossRef]
- Yang, J.H.; Wang, C.L.; Dai, H.L. Soil Agrochemical Analysis and Environmental Monitoring; Chine Land Press: Beijing, China, 2008. [Google Scholar]
- Huang, C. Soil Science; China Agriculture Press: Beijing, China, 2000; pp. 71–75. [Google Scholar]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. The vegan package. Community Ecol. Package 2007, 10, 719. [Google Scholar]
- Harrell, F.E., Jr.; Harrell, M.F.E., Jr. Package ‘hmisc’. CRAN 2019, 2019, 235–236. [Google Scholar]
- Rosseel, Y.; Oberski, D.; Byrnes, J.; Vanbrabant, L.; Savalei, V.; Merkle, E.; Hallquist, M.; Rhemtulla, M.; Katsikatsou, M.; Barendse, M.; et al. Package ‘lavaan’. CRAN 2017, 17, 2017. [Google Scholar]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Oxford University Press: London, UK, 1934. [Google Scholar]
- Eldridge, D.J.; Bowker, M.A.; Maestre, F.T.; Roger, E.; Reynolds, J.F.; Whitford, W.G. Impacts of shrub encroachment on ecosystem structure and functioning: Towards a global synthesis. Ecol. Lett. 2011, 14, 709–722. [Google Scholar] [CrossRef]
- Meng, B.; Jin, Z.; Chen, J.; Kuang, Y.; Yi, S.; Lv, Y.; Li, J. Critical signals for grassland desertification prediction in the transition zone between desert and typical steppe in InnerMongolia, China. Ecol. Indic. 2025, 170, 113065. [Google Scholar] [CrossRef]
- Yu, L.; Guo, T.; Sun, Z.; Ma, Y.; Li, Z.; Zhao, Y.; Wang, H. Seed Germination and Threshold Characteristics of Two Dominant Plant Species During the Transition from Desert Steppe to Shrubland. Acta Ecol. Sin. 2021, 41, 4160–4169. [Google Scholar]
- Shi, L.; Zhang, Z.J.; Zhang, C.Y.; Zhang, J.Z. Effects of sand burial on survival, growth, gas exchange and biomass allocation of Ulmus pumila seedlings in the Hunshandak Sandland, China. Ann. Bot. 2004, 94, 553–560. [Google Scholar] [CrossRef]
- Chengyi, Z.; Feihai, Y.; Ming, D. Effects of sand burial on the survival, growth, and biomass allocation in semi-shrub Hedysarum laeve seedlings. Acta Bot. Sin. 2002, 44, 337–343. [Google Scholar]
- Campanella, M.; Bisigato, A.J. Conspecific leaf litter and root competition inhibits shrub emergence in the Patagonian steppe. Plant Ecol. 2019, 220, 985–993. [Google Scholar] [CrossRef]
- Schenk, H.J.; Jackson, R.B. Rooting Depths, Lateral Root Spreads and Below-Ground/Above-Ground Allometries of Plants in Water-Limited Ecosystems. J. Ecol. 2002, 90, 480–494. [Google Scholar] [CrossRef]
- Zhu, Y.; Shen, H.; Akinyemi, D.S.; Zhang, P.; Feng, Y.; Zhao, M.; Kang, J.; Zhao, X.; Hu, H.; Fang, J. Increased precipitation attenuates shrub encroachment by facilitating herbaceous growth in a Mongolian grassland. Funct. Ecol. 2022, 36, 2356–2366. [Google Scholar] [CrossRef]
- Wu, H.; Zhao, G.; Li, X.-Y.; Wang, Y.; He, B.; Jiang, Z.; Zhang, S.; Sun, W. Identifying water sources used by alpine riparian plants in a restoration zone on the Qinghai-Tibet Plateau: Evidence from stable isotopes. Sci. Total Environ. 2019, 697, 134092. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Li, X.Y.; Jiang, Z.Y.; Wang, Y.M. Study on the Relationship Between Shrub Encroachment and Soil Properties in Typical Grasslands of Inner Mongolia. Acta Prataculturae Sin. 2017, 26, 224–230. [Google Scholar]
- Hou, H.; Yan, H.; Bai, X.; Zhang, Y.; Guo, Y.; Zhou, J.; Gao, S. A Synthetic Review of Feedbacks and Drivers of Shrub–Grass Interaction in the Process of Grassland Shrub Encroachment. Plants 2025, 14, 605. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Elorza, M.; Dana, E.D.; González, A.; Sobrino, E. Changes in the high-mountain vegetation of the central Iberian Peninsula as a probable sign of global warming. Ann. Bot. 2003, 92, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Dong, L.; Li, Z.; Zhang, J.; Li, Z.; Miao, B.; Jia, C.; Liang, C.; Wang, L.; Li, F.Y. Phylogenetic structure and formation mechanism of shrub communities in arid and semiarid areas of the Mongolian Plateau. Ecol. Evol. 2019, 9, 13320–13331. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Longitude | Latitude | Shrub Cover (%) | Density (plant/hm2) | Average Height (cm) | Number of Herbaceous Species | Herbaceous Biomass/(g/m2) |
|---|---|---|---|---|---|---|---|
| CT-1 | 108.2646 | 41.7069 | 10.24 ± 1.15 a | 2292.00 ± 129.46 a | 13.22 ± 4.02 a | 13 a | 17.83 ± 7.09 a |
| CT-2 | 109.2642 | 42.2174 | 1.89 ± 0.28 b | 854.00 ± 46.51 bc | 11.78 ± 1.87 b | 8 ab | 24.92 ± 4.36 ab |
| CT-3 | 108.8783 | 42.0381 | 2.71 ± 0.32 b | 583.33 ± 59.77 bc | 10.22 ± 2.35 bc | 9 bc | 28.04 ± 2.07 bc |
| CT-4 | 108.1888 | 42.0834 | 5.75 ± 0.53 b | 1410.67 ± 241.42 ab | 13.11 ± 2.10 a | 10 abc | 14.66 ± 5.03 abc |
| CT-5 | 107.9456 | 1.8527 | 4.65 ± 2.71 b | 1355.00 ± 727.65 ab | 8.56 ± 1.64 c | 8 abc | 13.02 ± 1.02 abc |
| CT-6 | 107.6377 | 41.8570 | 3.30 ± 0.78 b | 815.67 ± 194.02 bc | 9.67 ± 1.76 ab | 11 c | 13.02 ± 1.02 c |
| CT-7 | 108.4936 | 42.1973 | 1.53 ± 1.13 b | 308.00 ± 73.43 c | 11.56 ± 2.11 b | 9 abc | 19.67 ± 0.92 bc |
| CT-8 | 107.7494 | 42.1065 | 2.38 ± 1.43 b | 944.33 ± 513.92 bc | 9.22 ± 1.93 ab | 6 bc | 10.40 ± 0.57 c |
| Family | Genera | Species | Life Form | Growth Form |
|---|---|---|---|---|
| Poaceae | Cleistogenes | Cleistogenes songorica (Roshev.) Ohwi | H | PH |
| Cleistogenes squarrosa (Trin.) Keng | H | PH | ||
| Tragus | Tragus mongolorum Ohwi | T | AH | |
| Stipa | Stipa tianschanica var. klemenzii (Roshev.) Norl. | H | PH | |
| Stipa breviflora Griseb. | H | PH | ||
| Agropyron | Agropyron cristatum (L.) Gaertn. | H | PH | |
| Agropyron desertorum (Fisch.) Schult. | H | PH | ||
| Setaria | Setaria viridis (L.) P. Beauv. | T | AH | |
| Aristida | Aristida adscensionis L. | T | AH | |
| Enneapogon | Enneapogon desvauxii P. Beauv. | T | AH | |
| Eragrostis | Eragrostis pilosa (L.) P. Beauv. | T | AH | |
| Fabaceae | Astragalus | Astragalus galactites Pall. | H | PH |
| Caragana | Caragana microphylla Lam. | P | S | |
| Caragana tibetica Kom. | P | S | ||
| Asteraceae | Artemisia | Artemisia frigida Willd. | H | PH |
| Aster | Aster altaicus Willd. | H | AH | |
| Echinops | Echinops gmelinii Turcz. | T | AH | |
| Amaranthaceae | Kochia | Bassia prostrata (L.) Beck | H | SS |
| Zygophyllaceae | Tribulus | Tribulus terrestris L. | T | AH |
| Nitrariaceae | Peganum | Peganum nigellastrum Bunge | P | PH |
| Liliaceae | Allium | Allium tenuissimum L. | H | PH |
| Allium mongolicum Regel | Cr | PH | ||
| Allium polyrhizum Turcz. ex Regel | H | PH | ||
| Allium bidentatum Fisch. ex Prokh. | H | PH | ||
| Asparagus | Asparagus dauricus Link | H | PH | |
| Asparagus gobicus Ivanova ex Grubov | H | PH | ||
| Convolvulaceae | Convolvulus | Convolvulus ammannii Desr. | H | PH |
| Polygalaceae | Polygala | Polygala tenuifolia Willd. | C | PH |
| Iridaceae | Iris | Iris bungei Maxim. | H | PH |
| Rutaceae | Haplophyllum | Haplophyllum dauricum (L.) G. Don | T | PH |
| Asclepiadaceae | Cynanchum | Cynanchum thesiodes L. | H | PH |
| Caryophyllaceae | Arenaria | Eremogone juncea (M. Bieb.) Fenzl | T | AH |
| Chenopodiaceae | Krascheninnikovia | Krascheninnikovia ceratoide Gueldenst. | C | PH |
| Corispermum | Corispermum mongolicum Iljin | T | AH | |
| Salsola | Salsola collina Pall. | T | AH |
| Chi-Square-to-Degrees of Freedom Ratio (χ2/df) | Root Mean Square Error (RMSEA) | Standardized Root Mean Square Residual (SRMR) | Normed Fit Index (NFI) | Comparative Fit Index (CFI) | |
|---|---|---|---|---|---|
| Standard value | <0.05 | <0.08 | >0.9 | >0.9 | |
| Actual value | 4.364 | 0.000 | 0.061 | 1.185 | 1.000 |
| Factors | Direct Effects | Indirect Effects | Total Effects |
|---|---|---|---|
| Climate Factors | 0.23 | −0.03 | 0.20 |
| Soil Factors | 0.56 | 0.004 | 0.564 |
| Herbaceous Biomass | −0.04 | 0 | −0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Dong, L.; Li, J.; Lu, S.; Yi, L.; Li, H.; Chai, S.; Wang, J. Different Influences of Soil and Climatic Factors on Shrubs and Herbaceous Plants in the Shrub-Encroached Grasslands of the Mongolian Plateau. Forests 2025, 16, 696. https://doi.org/10.3390/f16040696
Liu Y, Dong L, Li J, Lu S, Yi L, Li H, Chai S, Wang J. Different Influences of Soil and Climatic Factors on Shrubs and Herbaceous Plants in the Shrub-Encroached Grasslands of the Mongolian Plateau. Forests. 2025; 16(4):696. https://doi.org/10.3390/f16040696
Chicago/Turabian StyleLiu, Yue, Lei Dong, Jinrong Li, Shuaizhi Lu, Liqing Yi, Huimin Li, Shaoqi Chai, and Jian Wang. 2025. "Different Influences of Soil and Climatic Factors on Shrubs and Herbaceous Plants in the Shrub-Encroached Grasslands of the Mongolian Plateau" Forests 16, no. 4: 696. https://doi.org/10.3390/f16040696
APA StyleLiu, Y., Dong, L., Li, J., Lu, S., Yi, L., Li, H., Chai, S., & Wang, J. (2025). Different Influences of Soil and Climatic Factors on Shrubs and Herbaceous Plants in the Shrub-Encroached Grasslands of the Mongolian Plateau. Forests, 16(4), 696. https://doi.org/10.3390/f16040696