
Global Warming Drives Shifts in the Suitable Habitats of Subalpine Shrublands in the Hengduan Mountains Region in China



Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Processing of Data
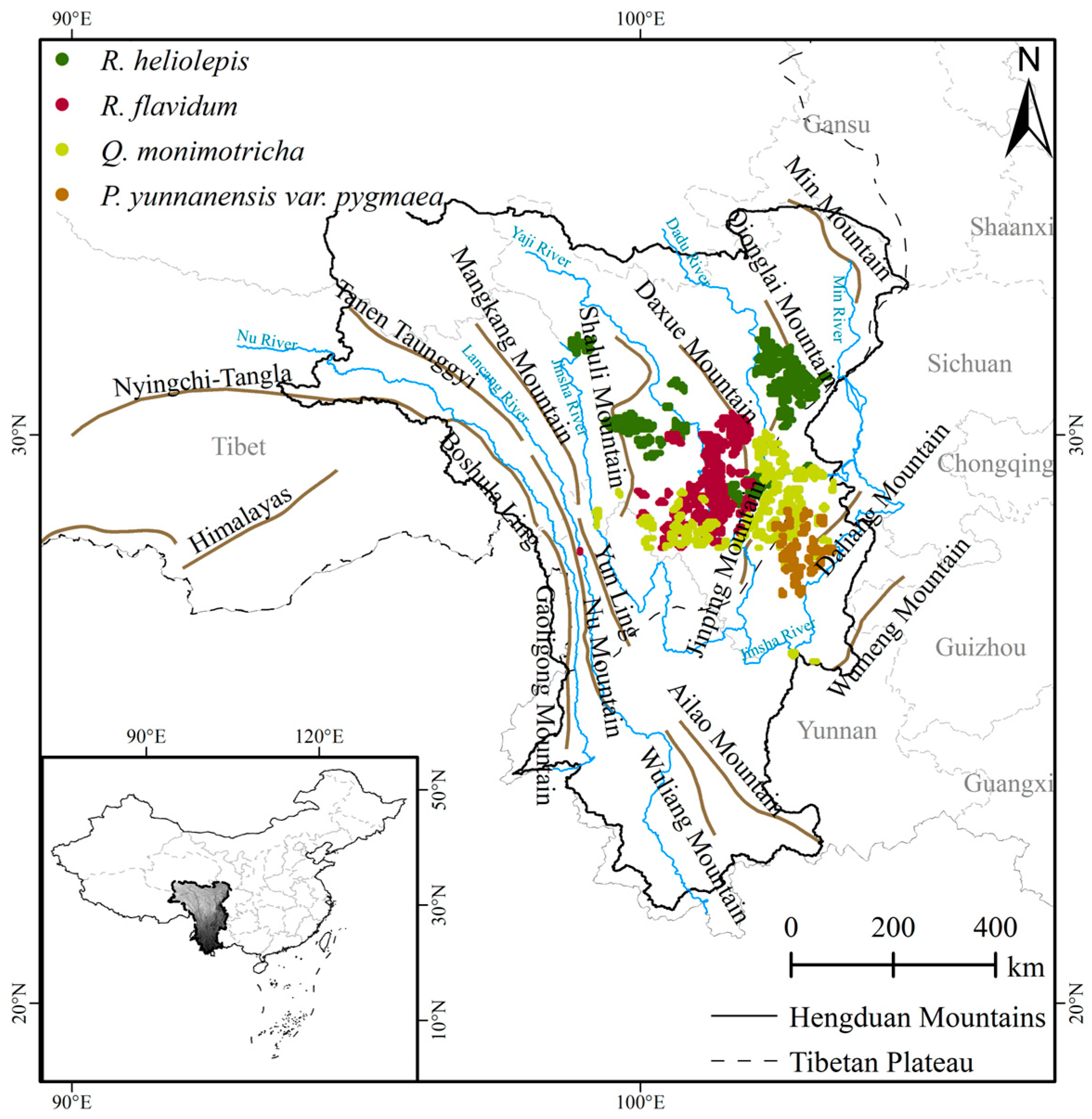
2.1.1. Vegetation Occurrence Data
2.1.2. Environmental Variable Data
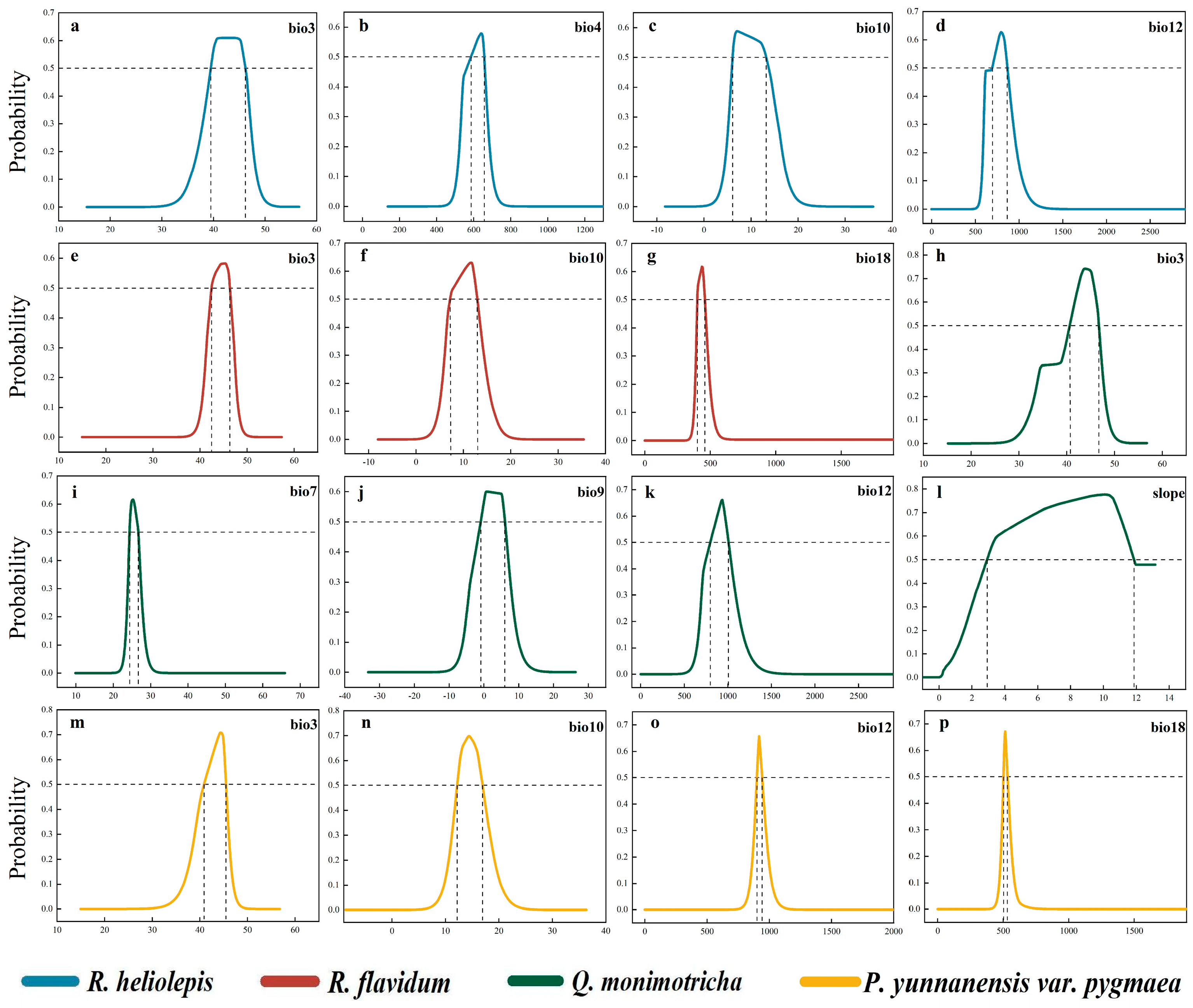
2.2. Classification of Suitable Habitats and Identification of Primary Drivers
2.3. Calculation of Contraction and Expansion, Centroid, and Elevation Shift in Suitable Habitats
3. Results
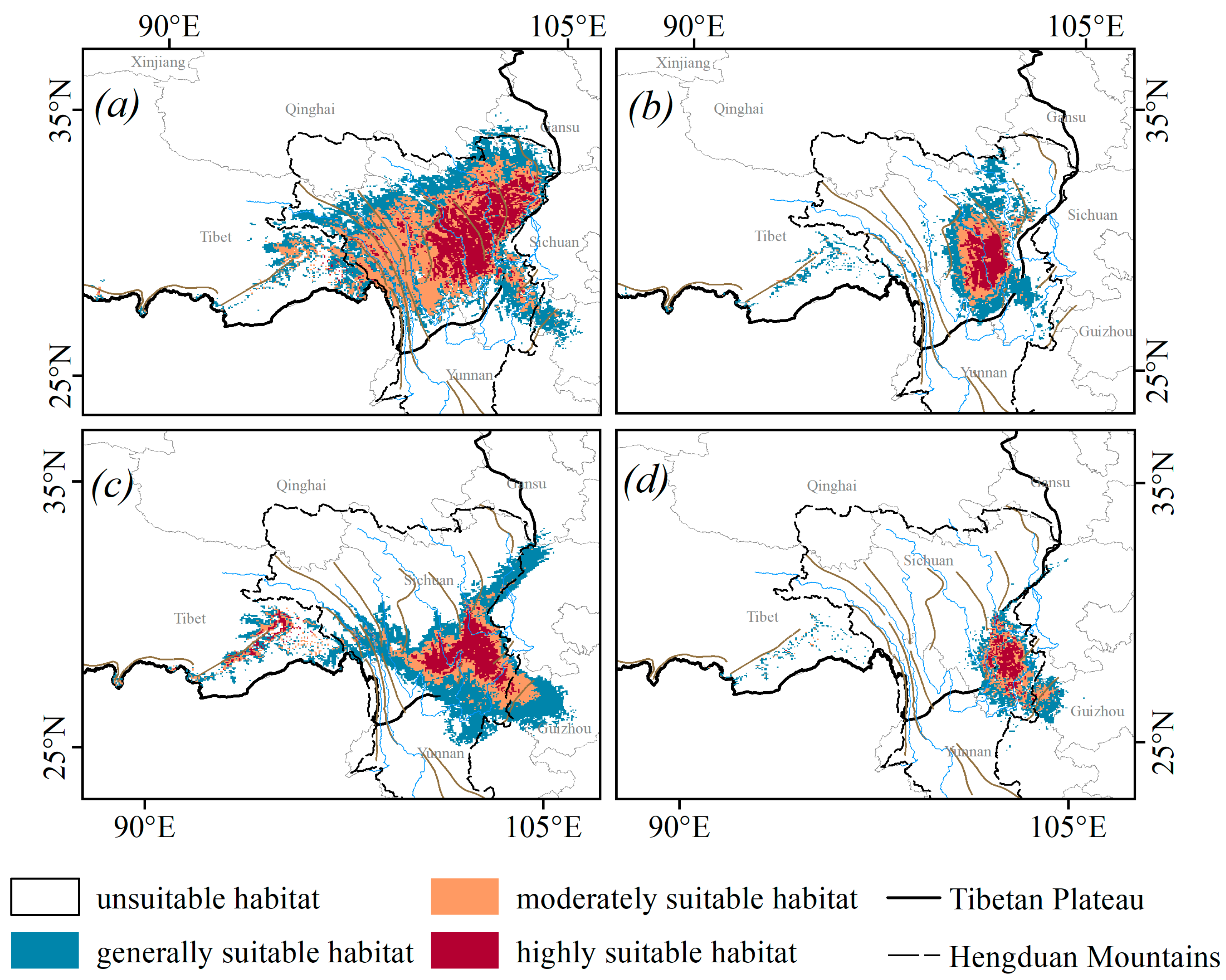
3.1. Suitable Habitat and Primary Drivers
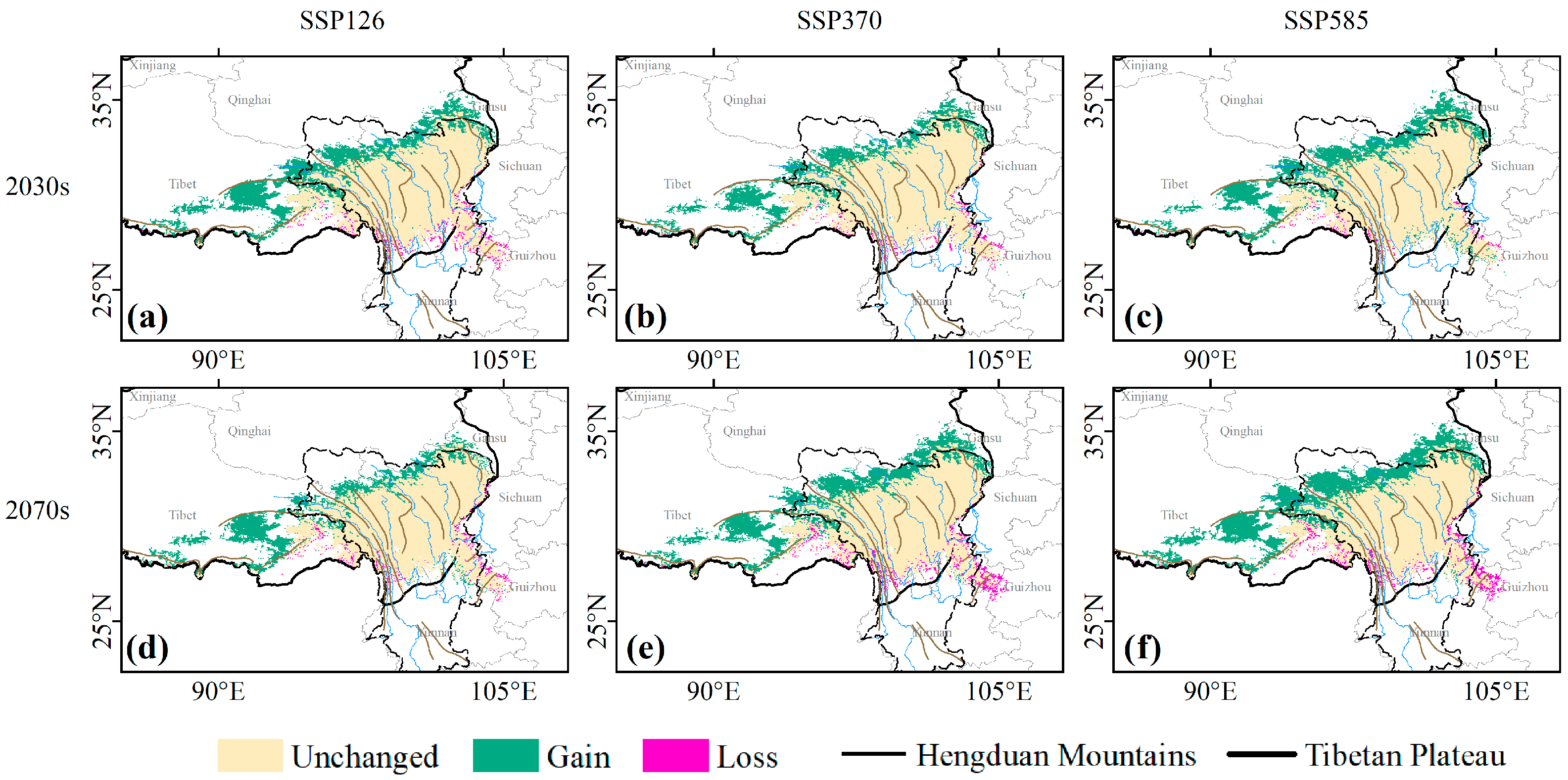
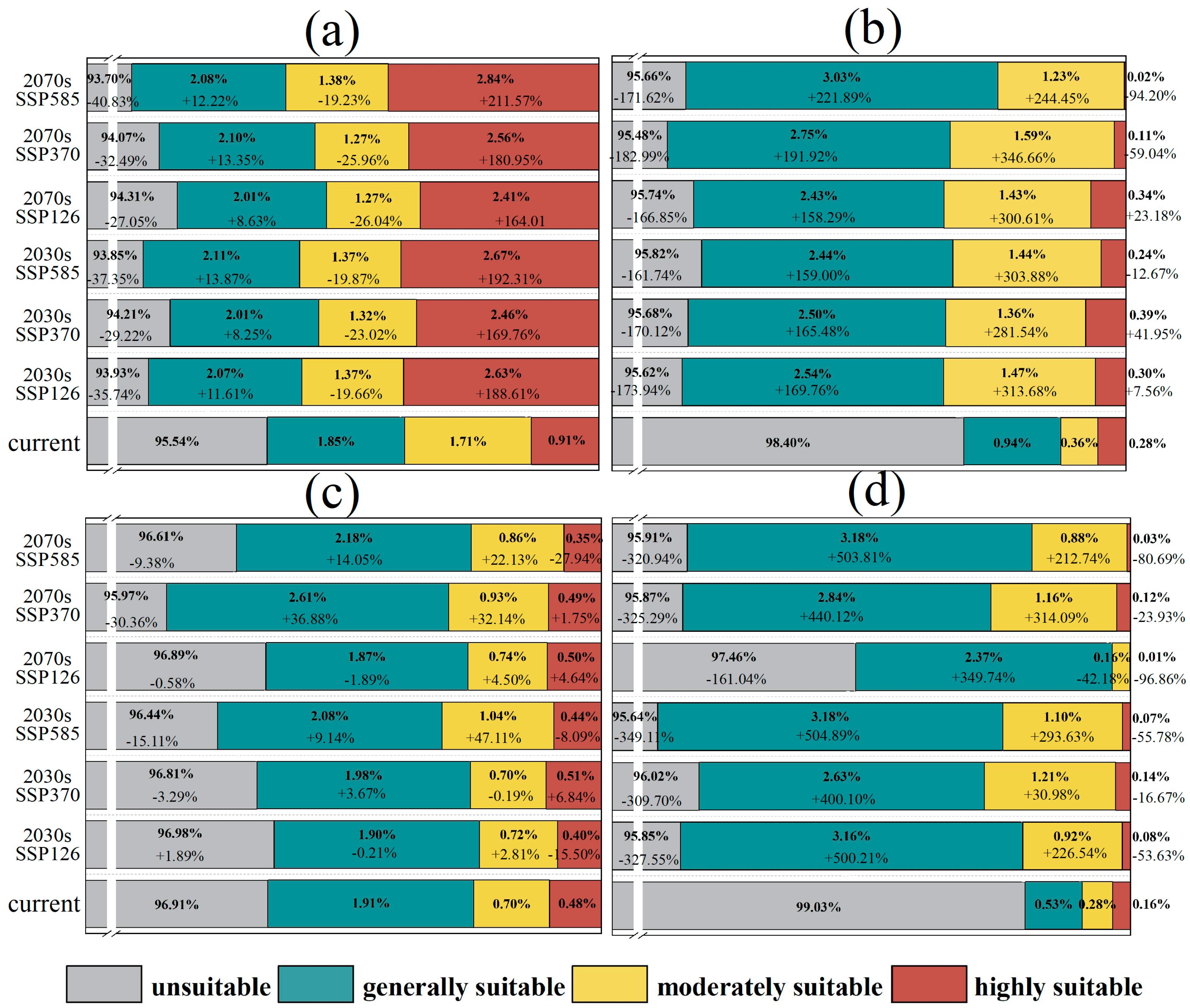
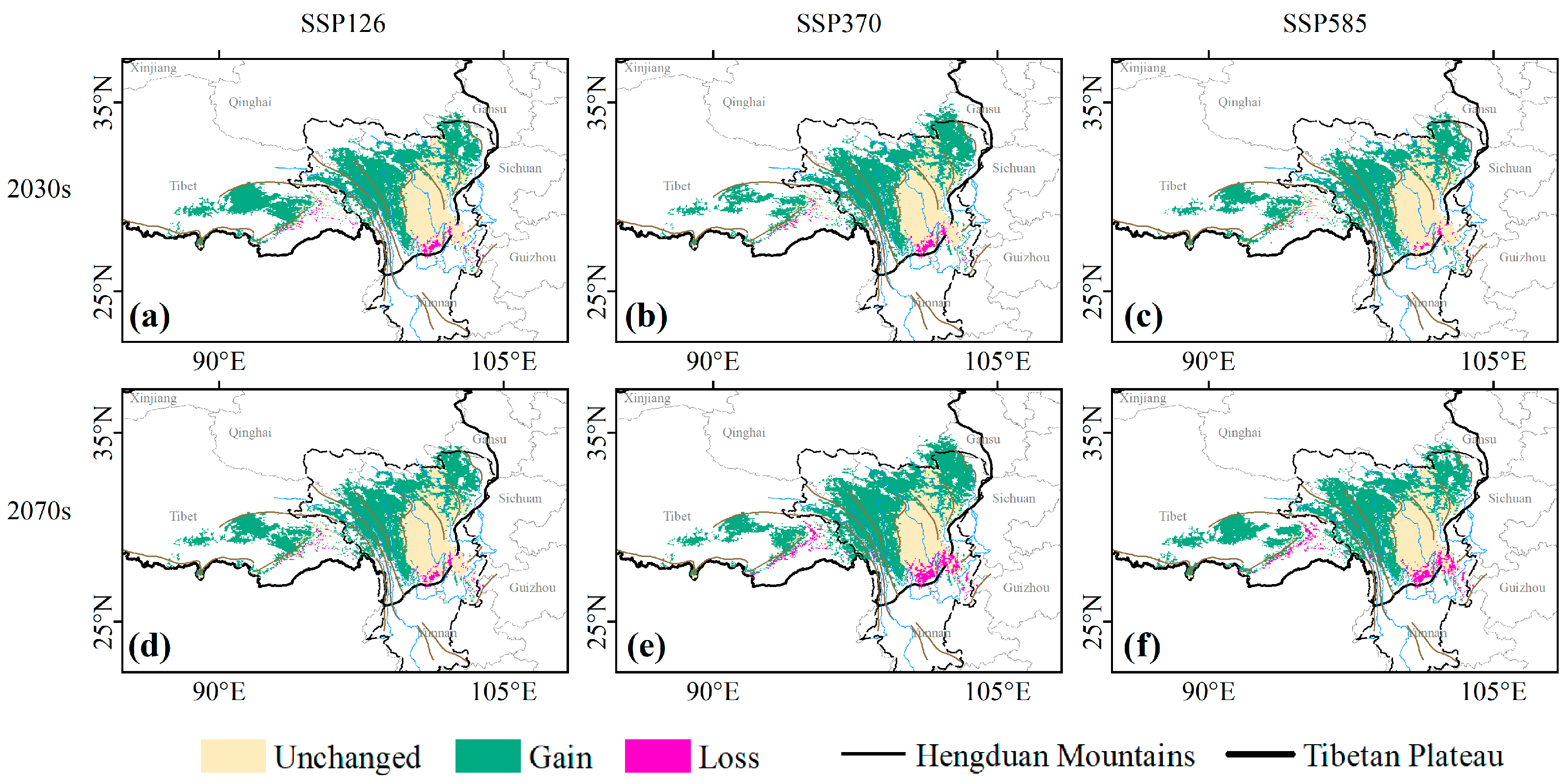
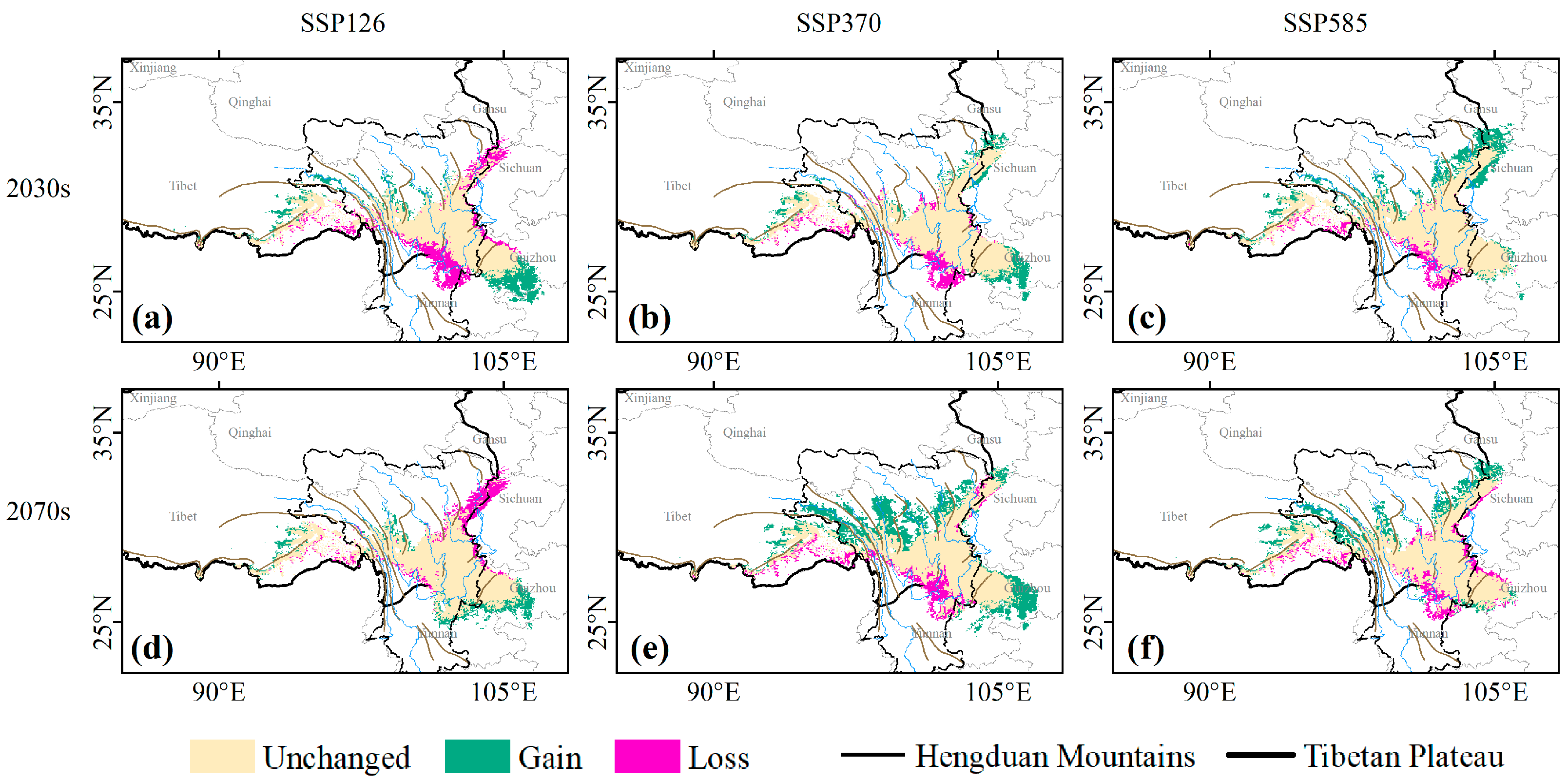
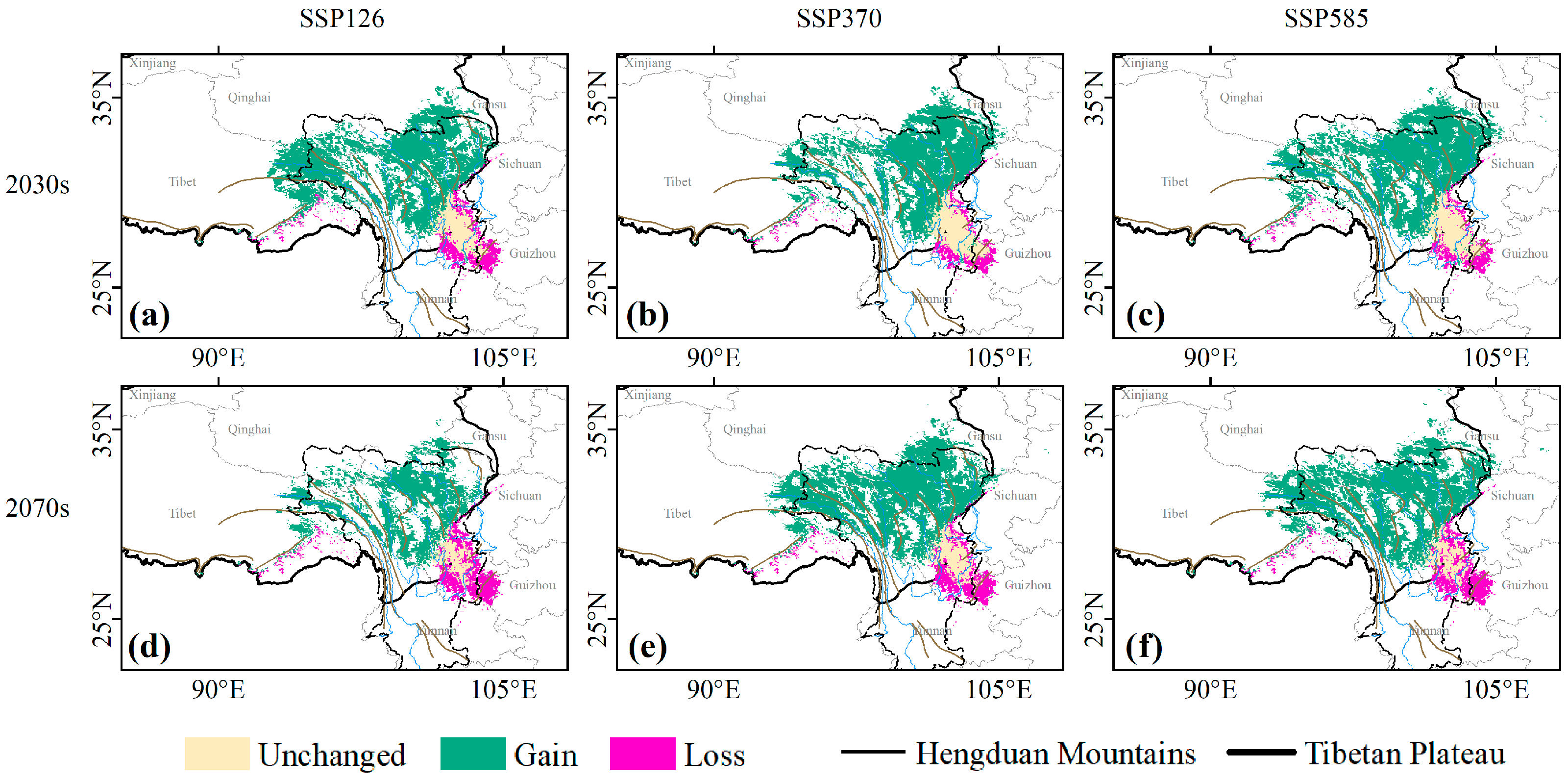
3.2. Contraction and Expansion of Suitable Habitats
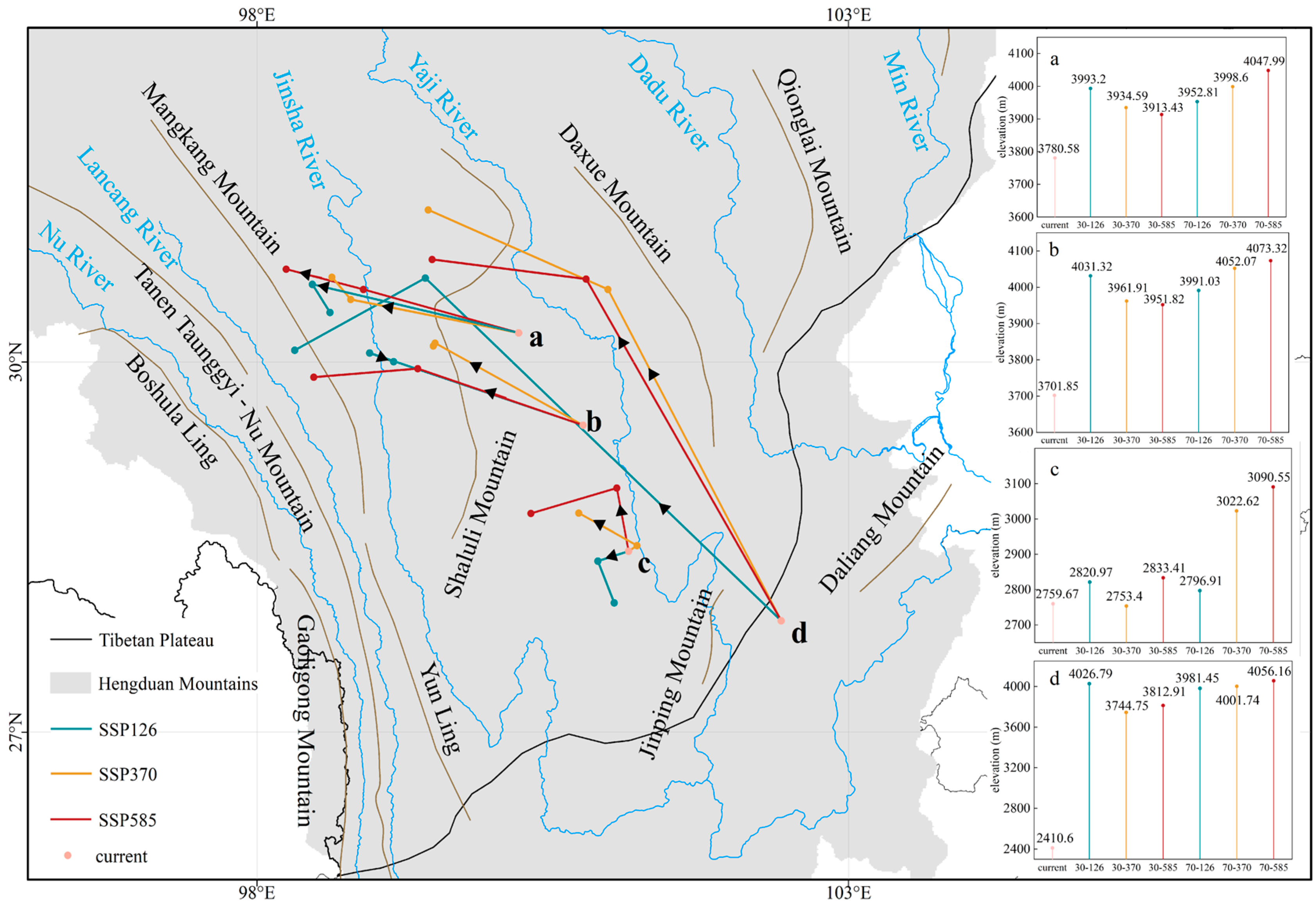
3.3. Centroid Migration and Elevation Shift of Suitable Habitats
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mori, A.S.; Isbell, F.; Cadotte, M.W. Assessing the Importance of Species and Their Assemblages for the Biodiversity-Ecosystem Multifunctionality Relationship. Ecology 2023, 104, e4104. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.G.; He, X.F.; Wang, S.W.; Yang, Y.; Sun, H. Cushion and Shrub Ecosystem Engineers Contribute Differently to Diversity and Functions in Alpine Ecosystems. J. Veg. Sci. 2019, 30, 362–374. [Google Scholar] [CrossRef]
- Luo, Y.H.; Cadotte, M.W.; Liu, J.; Burgess, K.S.; Tan, S.L.; Ye, L.; Zou, J.; Chen, Z.; Jiang, X.; Li, J.; et al. Multitrophic Diversity and Biotic Associations Influence Subalpine Forest Ecosystem Multifunctionality. Ecology 2022, 103, e3745. [Google Scholar] [CrossRef] [PubMed]
- Batllori, E.; Camarero, J.J.; Ninot, J.M.; Gutiérrez, E. Seedling Recruitment, Survival and Facilitation in Alpine Pinus Uncinata Tree Line Ecotones. Implications and Potential Responses to Climate Warming. Glob. Ecol. Biogeogr. 2009, 18, 460–472. [Google Scholar] [CrossRef]
- He, X.; Burgess, K.S.; Gao, L.-M.; Li, D.-Z. Distributional Responses to Climate Change for Alpine Species of Cyananthus and Primula Endemic to the Himalaya-Hengduan Mountains. Plant Divers. 2019, 41, 26–32. [Google Scholar] [CrossRef]
- Liu, J.Y.; Zou, H.X.; Bachelot, B.; Dong, T.; Zhu, Z.F.; Liao, Y.; Plenković-Moraj, A.; Wu, Y. Predicting the Responses of Subalpine Forest Landscape Dynamics to Climate Change on the Eastern Tibetan Plateau. Glob. Change Biol. 2021, 27, 4352–4366. [Google Scholar] [CrossRef]
- Liang, Q.L.; Xu, X.T.; Mao, K.S.; Wang, M.C.; Wang, K.; Xi, Z.X.; Liu, J.Q. Shifts in Plant Distributions in Response to Climate Warming in a Biodiversity Hotspot, the Hengduan Mountains. J. Biogeogr. 2018, 45, 1334–1344. [Google Scholar] [CrossRef]
- Lin, D.; Xia, J.; Wan, S. Climate Warming and Biomass Accumulation of Terrestrial Plants: A Meta-Analysis. New Phytol. 2010, 188, 187–198. [Google Scholar]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; et al. Extinction Risk from Climate Change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Raxworthy, C.J.; Pearson, R.G.; Rabibisoa, N.; Rakotondrazafy, A.M.; Ramanamanjato, J.B.; Raselimanana, A.P.; Wu, S.; Nussbaum, R.A.; Stone, D.A. Extinction Vulnerability of Tropical Montane Endemism from Warming and Upslope Displacement: A Preliminary Appraisal for the Highest Massif in Madagascar. Glob. Change Biol. 2008, 14, 1703–1720. [Google Scholar] [CrossRef]
- Hill, R.A.; Granica, K.; Smith, G.M.; Schardt, M. Representation of an Alpine Treeline Ecotone in SPOT 5 HRG Data. Remote Sens. Environ. 2007, 110, 458–467. [Google Scholar] [CrossRef]
- De Toma, A.; Carboni, M.; Bazzichetto, M.; Malavasi, M.; Cutini, M. Dynamics of Dwarf Shrubs in Mediterranean High-Mountain Ecosystems. J. Veg. Sci. 2022, 33, e13143. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, B.; Huang, T.; Chen, H.; Yue, J.; Tian, Y. Responses of the Distribution Pattern of the Suitable Habitat of Juniperus Tibetica Komarov to Climate Change on the Qinghai-Tibet Plateau. Forests 2023, 14, 434. [Google Scholar] [CrossRef]
- Yu, H.; Miao, S.; Xie, G.; Guo, X.; Chen, Z.; Favre, A. Contrasting Floristic Diversity of the Hengduan Mountains, the Himalayas and the Qinghai-Tibet Plateau Sensu Stricto in China. Front. Ecol. Evol. 2020, 8, 136. [Google Scholar] [CrossRef]
- Kuang, X.; Jiao, J.J. Review on Climate Change on the Tibetan Plateau during the Last Half Century. J. Geophys. Res. Atmos. 2016, 121, 3979–4007. [Google Scholar] [CrossRef]
- Fang, O.; Zhang, Q.-B. Tree Resilience to Drought Increases in the Tibetan Plateau. Glob. Change Biol. 2019, 25, 245–253. [Google Scholar] [CrossRef]
- Huang, Y.-J.; Zhu, H.; Su, T.; Spicer, R.A.; Hu, J.-J.; Jia, L.-B.; Zhou, Z.-K. Rise of Herbaceous Diversity at the Southeastern Margin of the Tibetan Plateau: First Insight from Fossils. J. Syst. Evol. 2022, 60, 1109–1123. [Google Scholar] [CrossRef]
- Kramer, A.; Herzschuh, U.; Mischke, S.; Zhang, C. Holocene Treeline Shifts and Monsoon Variability in the Hengduan Mountains (Southeastern Tibetan Plateau), Implications from Palynological Investigations. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 286, 23–41. [Google Scholar] [CrossRef]
- Chen, J.G.; Yang, Y.; Wang, S.W.; Sun, H.; Schöb, C. Shrub Facilitation Promotes Selective Tree Establishment beyond the Climatic Treeline. Sci. Total Environ. 2020, 708, 134618. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Sillero, N. What Does Ecological Modelling Model? A Proposed Classification of Ecological Niche Models Based on Their Underlying Methods. Ecol. Model. 2011, 222, 1343–1346. [Google Scholar] [CrossRef]
- Miller, J. Species Distribution Modeling. Geogr. Compass 2010, 4, 490–509. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting Species Distribution: Offering More than Simple Habitat Models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Archibald, C.L.; Summers, D.M.; Graham, E.M.; Bryan, B.A. Habitat Suitability Maps for Australian Flora and Fauna under CMIP6 Climate Scenarios. GigaScience 2024, 13, giae002. [Google Scholar] [CrossRef]
- Förderer, E.-M.; Rödder, D.; Langer, M.R. Global Diversity Patterns of Larger Benthic Foraminifera under Future Climate Change. Glob. Change Biol. 2023, 29, 969–981. [Google Scholar] [CrossRef]
- Dai, J.; Roberts, D.A.; Stow, D.A.; An, L.; Hall, S.J.; Yabiku, S.T.; Kyriakidis, P.C. Mapping Understory Invasive Plant Species with Field and Remotely Sensed Data in Chitwan, Nepal. Remote Sens. Environ. 2020, 250, 112037. [Google Scholar] [CrossRef]
- Luo, J.; Ma, Y.; Liu, Y.; Zhu, D.; Guo, X. Predicting Polygonum Capitatum Distribution in China across Climate Scenarios Using MaxEnt Modeling. Sci. Rep. 2024, 14, 20020. [Google Scholar] [CrossRef]
- Yang, Q.-S.; Chen, W.-Y.; Xia, K.; Zhou, Z.-K. Climatic Envelope of Evergreen Sclerophyllous Oaks and Their Present Distribution in the Eastern Himalaya and Hengduan Mountains. J. Syst. Evol. 2009, 47, 183–190. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, S.; Luo, T.; Zheng, W.; Yang, W.; Li, J.; Wang, Y.; Wang, S. Distribution Pattern of Pinus yunnanensis and P. yunnanensis var. pygmaea and Related Key Ecological Factors. J. Northeast For. Univ. 2021, 49, 8–14. [Google Scholar] [CrossRef]
- Hou, X.Y. Vegetation Atlas of China; Scientific Press: Beijing, China, 2001. [Google Scholar]
- Warren, D.L.; Matzke, N.J.; Cardillo, M.; Baumgartner, J.B.; Beaumont, L.J.; Turelli, M.; Glor, R.E.; Huron, N.A.; Simões, M.; Iglesias, T.L.; et al. ENMTools 1.0: An R Package for Comparative Ecological Biogeography. Ecography 2021, 44, 504–511. [Google Scholar] [CrossRef]
- Xu, L.; Fan, Y.; Zheng, J.; Guan, J.; Lin, J.; Wu, J.; Liu, L.; Wu, R.; Liu, Y. Impacts of Climate Change and Human Activity on the Potential Distribution of Aconitum Leucostomum in China. Sci. Total Environ. 2024, 912, 168829. [Google Scholar] [CrossRef] [PubMed]
- WorldClim. Available online: https://www.worldclim.org/ (accessed on 10 June 2024).
- Harmonized World Soil Database v1.2. Available online: https://www.fao.org/soils-portal/soil-survey/soil-maps-and-databases/harmonized-world-soil-database-v12/en/ (accessed on 13 June 2024).
- Last of the Wild Project, Version 3 (LWP-3): 1993 Human Footprint, 2018 Release. Available online: https://www.earthdata.nasa.gov/data/catalog/ (accessed on 15 June 2024).
- Wu, T.; Lu, Y.; Fang, Y.; Xin, X.; Li, L.; Li, W.; Jie, W.; Zhang, J.; Liu, Y.; Zhang, L.; et al. The Beijing Climate Center Climate System Model (BCC-CSM): The Main Progress from CMIP5 to CMIP6. Geosci. Model Dev. 2019, 12, 1573–1600. [Google Scholar] [CrossRef]
- Huang, B.; Chen, S.; Xu, L.; Jiang, H.; Chen, X.; He, H.; Chen, T. Predicting the Potential Geographical Distribution of Zingiber Striolatum Diels (Zingiberaceae), a Medicine Food Homology Plant in China. Sci. Rep. 2024, 14, 22206. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, J.; Ren, G.; Zhao, K.; Wang, X. Global Potential Distribution Prediction of Xanthium Italicum Based on Maxent Model. Sci. Rep. 2021, 11, 16545. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Tao, J. Predicting the Potential Distribution of Paeonia Veitchii (Paeoniaceae) in China by Incorporating Climate Change into a Maxent Model. Forests 2019, 10, 190. [Google Scholar] [CrossRef]
- Yang, X.-Q.; Kushwaha, S.P.S.; Saran, S.; Xu, J.; Roy, P.S. Maxent Modeling for Predicting the Potential Distribution of Medicinal Plant, Justicia Adhatoda L. in Lesser Himalayan Foothills. Ecol. Eng. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Zhu, C.; Qu, Y.; Wang, H. Predicting the Potential Distribution of an Invasive Species, Erigeron Canadensis L., in China with a Maximum Entropy Model. Glob. Ecol. Conserv. 2020, 21, e00822. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, B.; Meng, Y.; Jia, Y.; Xu, Q.; Pan, Y. The Influence of Climate Change on the Distribution of Hibiscus Mutabilis in China: MaxEnt Model-Based Prediction. Plants 2024, 13, 1744. [Google Scholar] [CrossRef]
- Li, J.; Fan, G.; He, Y. Predicting the Current and Future Distribution of Three Coptis Herbs in China under Climate Change Conditions, Using the MaxEnt Model and Chemical Analysis. Sci. Total Environ. 2020, 698, 134141. [Google Scholar] [CrossRef]
- He, Y.; Ma, J.; Chen, G. Potential Geographical Distribution and Its Multi-Factor Analysis of Pinus Massoniana in China Based on the Maxent Model. Ecol. Indic. 2023, 154, 110790. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Wani, I.A.; Khan, S.; Verma, S.; Al-Misned, F.A.; Shafik, H.M.; El-Serehy, H.A. Predicting Habitat Suitability and Niche Dynamics of Dactylorhiza Hatagirea and Rheum Webbianum in the Himalaya under Projected Climate Change. Sci. Rep. 2022, 12, 13205. [Google Scholar] [CrossRef]
- Wei, L.; Wang, G.; Xie, C.; Gao, Z.; Huang, Q.; Jim, C.Y. Predicting Suitable Habitat for the Endangered Tree Ormosia Microphylla in China. Sci. Rep. 2024, 14, 10330. [Google Scholar] [CrossRef]
- Hosseini, N.; Ghorbanpour, M.; Mostafavi, H. Habitat Potential Modelling and the Effect of Climate Change on the Current and Future Distribution of Three Thymus Species in Iran Using MaxEnt. Sci. Rep. 2024, 14, 3641. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, C.; Gong, G.; Chen, Y.; Ma, S.; Wu, Y.; Wang, H.; Li, Y.; Duan, H. Biodiversity Conservation and Management of Lake Wetlands Based on the Spatiotemporal Evolution Patterns of Crane Habitats. J. Environ. Manage. 2024, 353, 120257. [Google Scholar] [CrossRef]
- Yu, F.; Groen, T.A.; Wang, T.; Skidmore, A.K.; Huang, J.; Ma, K. Climatic Niche Breadth Can Explain Variation in Geographical Range Size of Alpine and Subalpine Plants. Int. J. Geogr. Inf. Sci. 2017, 31, 190–212. [Google Scholar]
- Li, K.-J.; Liu, X.-F.; Zhang, J.-H.; Zhou, X.-L.; Yang, L.; Shen, S.-K. Complexity Responses of Rhododendron Species to Climate Change in China Reveal Their Urgent Need for Protection. For. Ecosyst. 2023, 10, 100124. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Y.; Yan, Y.; Wen, Y.; Zhang, D. Geographic Distribution and Impacts of Climate Change on the Suitable Habitats of Two Alpine Rhododendron in Southwest China. Glob. Ecol. Conserv. 2024, 54, e03176. [Google Scholar] [CrossRef]
- He, Z.; Du, J.; Zhao, W.; Yang, J.; Chen, L.; Zhu, X.; Chang, X.; Liu, H. Assessing Temperature Sensitivity of Subalpine Shrub Phenology in Semi-Arid Mountain Regions of China. Agric. For. Meteorol. 2015, 213, 42–52. [Google Scholar] [CrossRef]
- Sun, M.R.; Sun, P.S.; Liu, N.; Zhang, L.; Yu, Z.; Feng, Q.; Smettem, K.; Liu, S. Alternating Dominant Effects of Temperature and Precipitation along Elevational Gradient on the Alpine and Subalpine Vegetation Activities in Southwestern China. For. Ecol. Manag. 2024, 554, 121668. [Google Scholar] [CrossRef]
- Bunting, E.L.; Munson, S.M.; Villarreal, M.L. Climate Legacy and Lag Effects on Dryland Plant Communities in the Southwestern U.S. Ecol. Indic. 2017, 74, 216–229. [Google Scholar] [CrossRef]
- Mirhashemi, H.; Heydari, M.; Ahmadi, K.; Karami, O.; Kavgaci, A.; Matsui, T.; Heung, B. Species Distribution Models of Brant’s Oak (Quercus Brantii Lindl.): The Impact of Spatial Database on Predicting the Impacts of Climate Change. Ecol. Eng. 2023, 194, 107038. [Google Scholar] [CrossRef]
- Ouyang, X.; Lin, H.; Bai, S.; Chen, J.; Chen, A. Simulation the Potential Distribution of Dendrolimus Houi and Its Hosts, Pinus Yunnanensis and Cryptomeria Fortunei, under Climate Change in China. Front. Plant Sci. 2022, 13, 1054710. [Google Scholar] [CrossRef]
- Yu, F.; Wang, T.; Groen, T.A.; Skidmore, A.K.; Yang, X.; Ma, K.; Wu, Z. Climate and Land Use Changes Will Degrade the Distribution of Rhododendrons in China. Sci. Total Environ. 2019, 659, 515–528. [Google Scholar] [CrossRef]
- Luo, W.; Sun, C.; Yang, S.; Chen, W.; Sun, Y.; Li, Z.; Liu, J.; Tao, W.; Tao, J. Contrasting Range Changes and Drivers of Four Forest Foundation Species under Future Climate Change in China. Sci. Total Environ. 2024, 942, 173784. [Google Scholar] [CrossRef]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional Vegetation Die-off in Response to Global-Change-Type Drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef]
- Ranjitkar, S.; Kindt, R.; Sujakhu, N.M.; Hart, R.; Guo, W.; Yang, X.; Shrestha, K.K.; Xu, J.; Luedeling, E. Separation of the Bioclimatic Spaces of Himalayan Tree Rhododendron Species Predicted by Ensemble Suitability Models. Glob. Ecol. Conserv. 2014, 1, 2–12. [Google Scholar] [CrossRef]
- Xun, Q.L.; An, S.Z.; Lu, M.Z. Climate Change and Topographic Differences Influence Grassland Vegetation Greening across Environmental Gradients. Front. Environ. Sci. 2024, 11, 1324742. [Google Scholar] [CrossRef]
- Hassan, S.S.; Goheer, M.A.; Farah, H.; Nadeem, M.; Muazzam, A.; Munir, J.A.; Fatima, S. Monitoring the Effects of Climate Change and Topography on Vegetation Health in Tharparkar, Pakistan. J. Water Clim. Change 2024, 15, 3472–3486. [Google Scholar] [CrossRef]
- Elliott, G.P.; Kipfmueller, K.F. Multi-Scale Influences of Slope Aspect and Spatial Pattern on Ecotonal Dynamics at Upper Treeline in the Southern Rocky Mountains, U.S.A. Arct. Antarct. Alp. Res. 2010, 42, 45–56. [Google Scholar] [CrossRef]
- Fu, B.; Wang, J.; Chen, L.; Qiu, Y. The Effects of Land Use on Soil Moisture Variation in the Danangou Catchment of the Loess Plateau, China. CATENA 2003, 54, 197–213. [Google Scholar] [CrossRef]
- Xiong, X.T.; Li, C.H.; Chen, J.H. Topographic regulatory role of vegetation response to climate change. Acta Geogr. Sin. 2023, 78, 2256–2270. [Google Scholar] [CrossRef]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and Its Drivers. Nat. Clim. Change 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Fang, T.; Shao, Y.; Oswald, T.; Lineaweaver, W.C.; Zhang, F. Effect of Sildenafil on Peripheral Nerve Regeneration. Ann. Plast. Surg. 2013, 70, 62–65. [Google Scholar] [CrossRef]
- De Toma, A.; Malavasi, M.; Marzialetti, F.; Cutini, M. Unveiling Spatial Patterns and Trajectories of Shrub Dynamics in Mediterranean Alpine Ecosystems. Plant Ecol. 2025, 226, 149–160. [Google Scholar] [CrossRef]
- Pérez-García, N.; Font, X.; Ferré, A.; Carreras, J. Drastic Reduction in the Potential Habitats for Alpine and Subalpine Vegetation in the Pyrenees Due to Twenty-First-Century Climate Change. Reg. Environ. Change 2013, 13, 1157–1169. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Forbes, B.C.; Wilmking, M.; Hallinger, M.; Lantz, T.; Blok, D.; Tape, K.D.; Macias-Fauria, M.; Sass-Klaassen, U.; Lévesque, E.; et al. Shrub Expansion in Tundra Ecosystems: Dynamics, Impacts and Research Priorities. Environ. Res. Lett. 2011, 6, 045509. [Google Scholar] [CrossRef]
- Hallinger, M.; Manthey, M.; Wilmking, M. Establishing a Missing Link: Warm Summers and Winter Snow Cover Promote Shrub Expansion into Alpine Tundra in Scandinavia. New Phytol. 2010, 186, 890–899. [Google Scholar] [CrossRef]
- Albrecht, E.C.; Dobbert, S.; Pape, R.; Löffler, J. Complex Environmental Control of Growth in a Dominant Mediterranean-Alpine Shrub Species. J. Ecol. 2024, 112, 1516–1532. [Google Scholar] [CrossRef]
- Feng, J.; Wang, B.; Xian, M.; Zhou, S.; Huang, C.; Cui, X. Prediction of Future Potential Distributions of Pinus Yunnanensis Varieties under Climate Change. Front. For. Glob. Change 2023, 6, 1308416. [Google Scholar] [CrossRef]
- Bertrand, R.; Lenoir, J.; Piedallu, C.; Riofrío-Dillon, G.; de Ruffray, P.; Vidal, C.; Pierrat, J.-C.; Gégout, J.-C. Changes in Plant Community Composition Lag behind Climate Warming in Lowland Forests. Nature 2011, 479, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chang, H.; Liu, T.; Zhang, C. The Potential Geographical Distribution of Haloxylon across Central Asia under Climate Change in the 21st Century. Agric. For. Meteorol. 2019, 275, 243–254. [Google Scholar] [CrossRef]
- Lamprecht, A.; Semenchuk, P.R.; Steinbauer, K.; Winkler, M.; Pauli, H. Climate Change Leads to Accelerated Transformation of High-Elevation Vegetation in the Central Alps. New Phytol. 2018, 220, 447–459. [Google Scholar] [CrossRef]
- Walther, G.-R.; Beißner, S.; Burga, C.A. Trends in the Upward Shift of Alpine Plants. J. Veg. Sci. 2005, 16, 541–548. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent Plant Diversity Changes on Europe’s Mountain Summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef]
- Lenoir, J.; Svenning, J.-C. Climate-Related Range Shifts—A Global Multidimensional Synthesis and New Research Directions. Ecography 2015, 38, 15–28. [Google Scholar] [CrossRef]
- Crimmins, S.M.; Dobrowski, S.Z.; Greenberg, J.A.; Abatzoglou, J.T.; Mynsberge, A.R. Changes in Climatic Water Balance Drive Downhill Shifts in Plant Species’ Optimum Elevations. Science 2011, 331, 324–327. [Google Scholar] [CrossRef]
- Zheng, X.Y.; Babst, F.; Camarero, J.J.; Li, X.X.; Lu, X.M.; Gao, S.; Sigdel, S.R.; Wang, Y.; Zhu, H.; Liang, E. Density-Dependent Species Interactions Modulate Alpine Treeline Shifts. Ecol. Lett. 2024, 27, e14403. [Google Scholar] [CrossRef]
- Pepin, N.; Bradley, R.S.; Diaz, H.F.; Baraer, M.; Caceres, E.B.; Forsythe, N.; Fowler, H.; Greenwood, G.; Hashmi, M.Z.; Liu, X.D.; et al. Elevation-Dependent Warming in Mountain Regions of the World. Nat. Clim. Change 2015, 5, 424–430. [Google Scholar] [CrossRef]
- Rumpf, S.B.; Hülber, K.; Wessely, J.; Willner, W.; Moser, D.; Gattringer, A.; Klonner, G.; Zimmermann, N.E.; Dullinger, S. Extinction Debts and Colonization Credits of Non-Forest Plants in the European Alps. Nat. Commun. 2019, 10, 4293. [Google Scholar] [CrossRef]
- Neilson, R.P.; Pitelka, L.F.; Solomon, A.M.; Nathan, R.; Midgley, G.F.; Fragoso, J.M.V.; Lischke, H.; Thompson, K. Forecasting Regional to Global Plant Migration in Response to Climate Change. BioScience 2005, 55, 749–759. [Google Scholar] [CrossRef]
- Hampe, A. Plants on the Move: The Role of Seed Dispersal and Initial Population Establishment for Climate-Driven Range Expansions. Acta Oecologica 2011, 37, 666–673. [Google Scholar] [CrossRef]
- Williams, M.I.; Dumroese, R.K. Preparing for Climate Change: Forestry and Assisted Migration. J. For. 2013, 111, 287–297. [Google Scholar] [CrossRef]
- Vitt, P.; Havens, K.; Kramer, A.T.; Sollenberger, D.; Yates, E. Assisted Migration of Plants: Changes in Latitudes, Changes in Attitudes. Biol. Conserv. 2010, 143, 18–27. [Google Scholar] [CrossRef]
- Lazarus, E.D.; McGill, B.J. Pushing the Pace of Tree Species Migration. PLoS ONE 2014, 9, e105380. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Description | Vegetation Type |
|---|---|---|
| bio2 (°C) | Mean diurnal range | △ |
| bio3 (%) | Isothermality | △▲○◆ |
| bio4 | Temperature seasonality | △▲○ |
| bio7 (°C) | Temperature annual range | ○◆ |
| bio9 (°C) | Mean temperature of driest quarter | ▲○ |
| bio10 (°C) | Mean temperature of warmest quarter | △▲◆ |
| bio12 (mm) | Annual precipitation | ▲○◆ |
| bio14 (mm) | Precipitation of driest month | △▲ |
| bio15 | Precipitation seasonality | ○◆ |
| bio18 (mm) | Precipitation of warmest quarter | △◆ |
| bio19 (mm) | Precipitation of coldest quarter | ○◆ |
| Awc (mm/m) | Available water storage capacity | △▲ |
| t-pH (−log (H+)) | Topsoil pH (H2O) | △◆ |
| t-texture | Topsoil texture | △▲◆ |
| Aspect (°) | Aspect | ◆ |
| Slope (°) | Slope | △▲○◆ |
| Hf | Human footprint | △▲○◆ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Yu, Y.; Ji, X.; Wang, Z.; Liu, Z. Global Warming Drives Shifts in the Suitable Habitats of Subalpine Shrublands in the Hengduan Mountains Region in China. Forests 2025, 16, 624. https://doi.org/10.3390/f16040624
Zhang H, Yu Y, Ji X, Wang Z, Liu Z. Global Warming Drives Shifts in the Suitable Habitats of Subalpine Shrublands in the Hengduan Mountains Region in China. Forests. 2025; 16(4):624. https://doi.org/10.3390/f16040624
Chicago/Turabian StyleZhang, Huayong, Yunyan Yu, Xiande Ji, Zhongyu Wang, and Zhao Liu. 2025. "Global Warming Drives Shifts in the Suitable Habitats of Subalpine Shrublands in the Hengduan Mountains Region in China" Forests 16, no. 4: 624. https://doi.org/10.3390/f16040624
APA StyleZhang, H., Yu, Y., Ji, X., Wang, Z., & Liu, Z. (2025). Global Warming Drives Shifts in the Suitable Habitats of Subalpine Shrublands in the Hengduan Mountains Region in China. Forests, 16(4), 624. https://doi.org/10.3390/f16040624