
Construction and Comparison of Single-Tree Biomass Model for Dendrocalamus brandisii

 ,
,
Abstract
1. Introduction
2. Materials and Methods
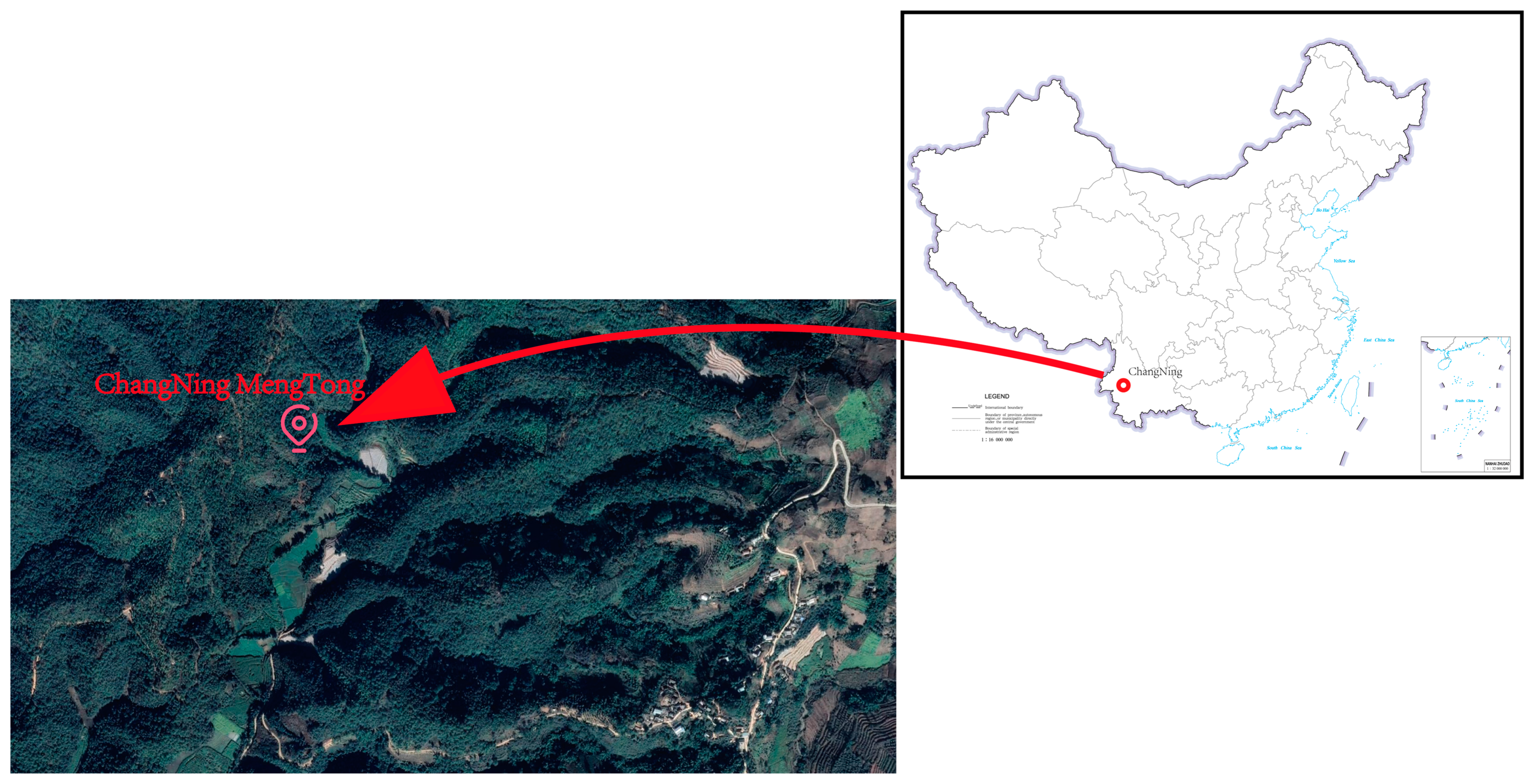
2.1. Study Area
2.2. Data Source
2.2.1. Selection and Sample Collection of Standard Bamboo
2.2.2. Sample Indicator Measurement
- MC represents the moisture content;
- Wdry represents the dry weight of the organ (in kg);
- Wfresh represents the fresh weight of the organ (in kg).
2.3. Model Construction and Selection
2.3.1. Model Construction
2.3.2. The Criteria for Selecting Models
2.4. Analysis Tools
3. Results
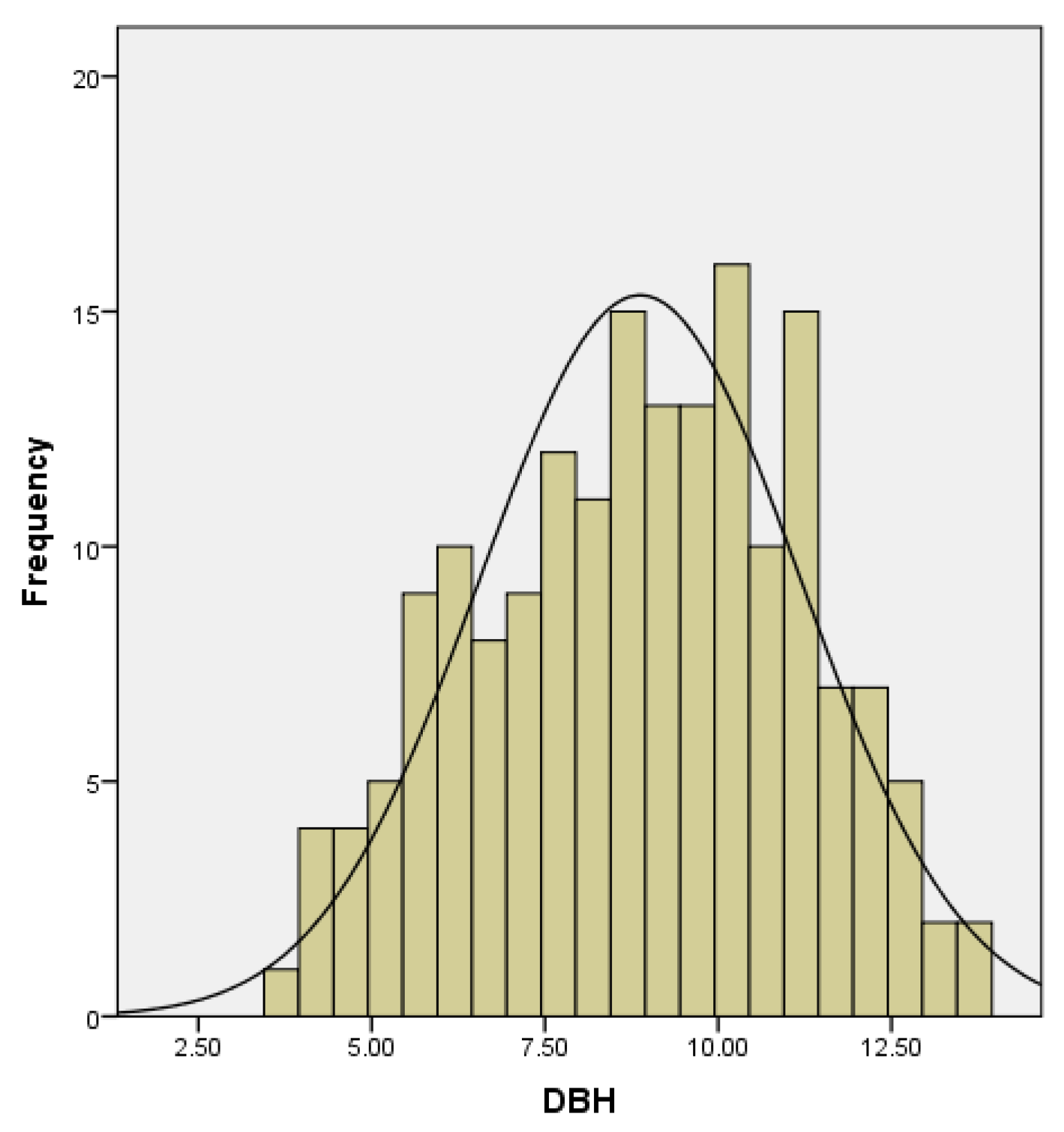
3.1. Data Collection
3.1.1. Analysis of the Water Content in the Organs of D. brandisii
3.1.2. Aboveground Biomass Proportion of Each Organ in D. brandisii
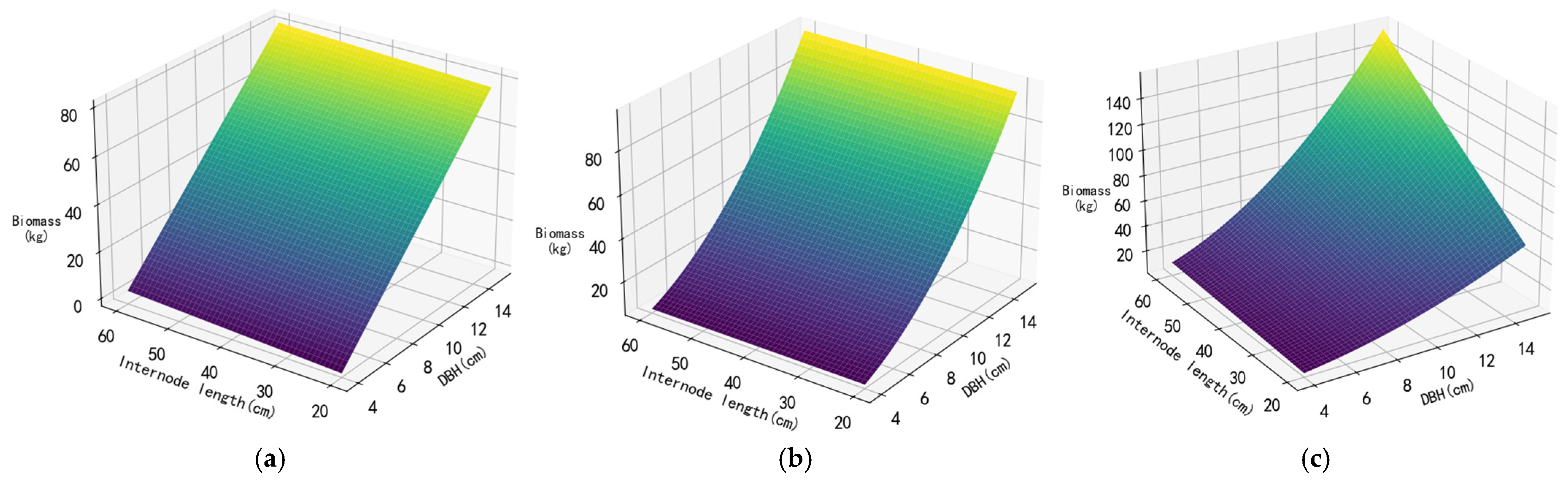
3.2. Construction of Biomass Regression Model
3.3. Selection of Biomass Models
3.3.1. Performance Metrics of the Model
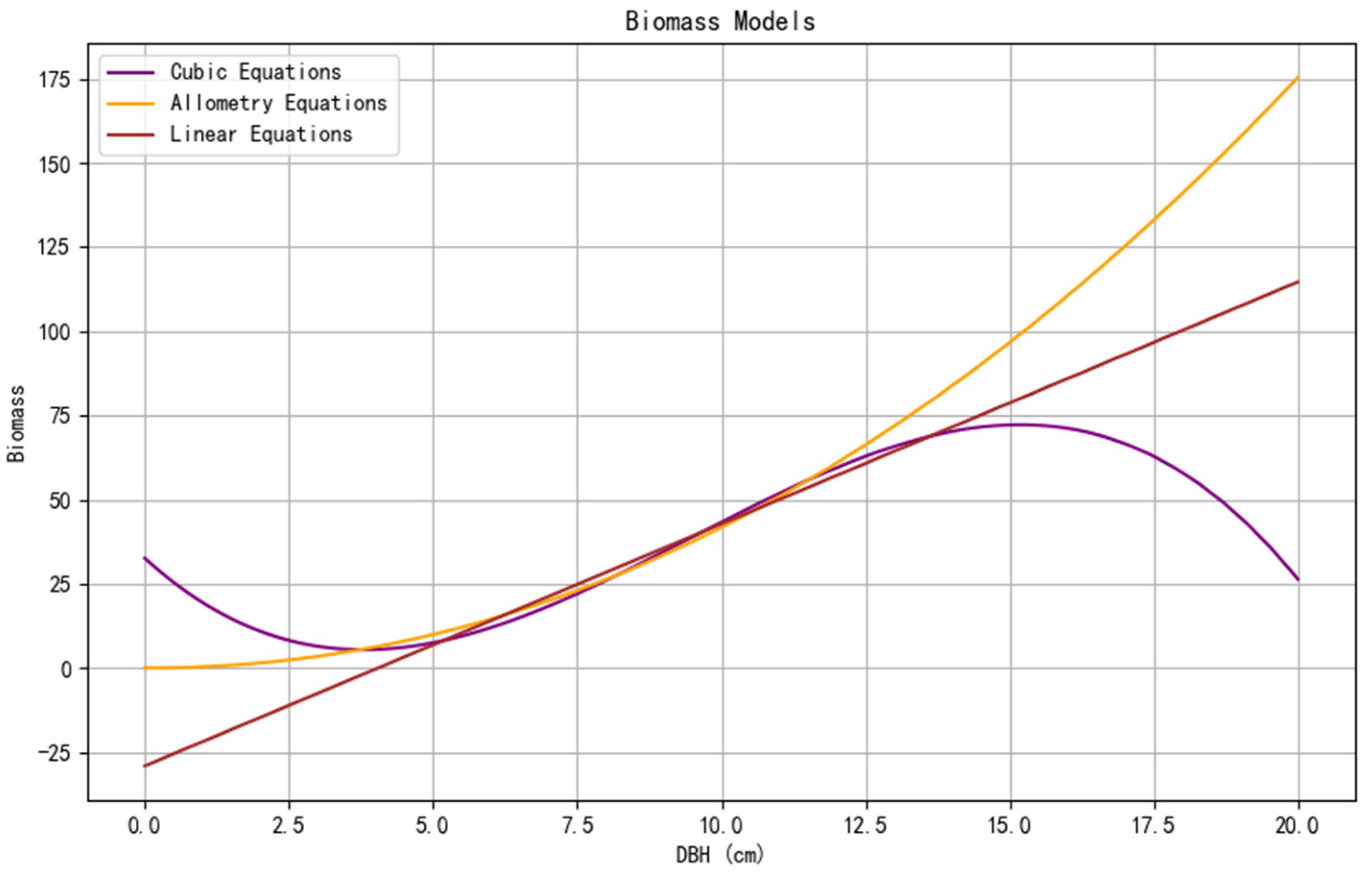
3.3.2. Fitting Results of Models
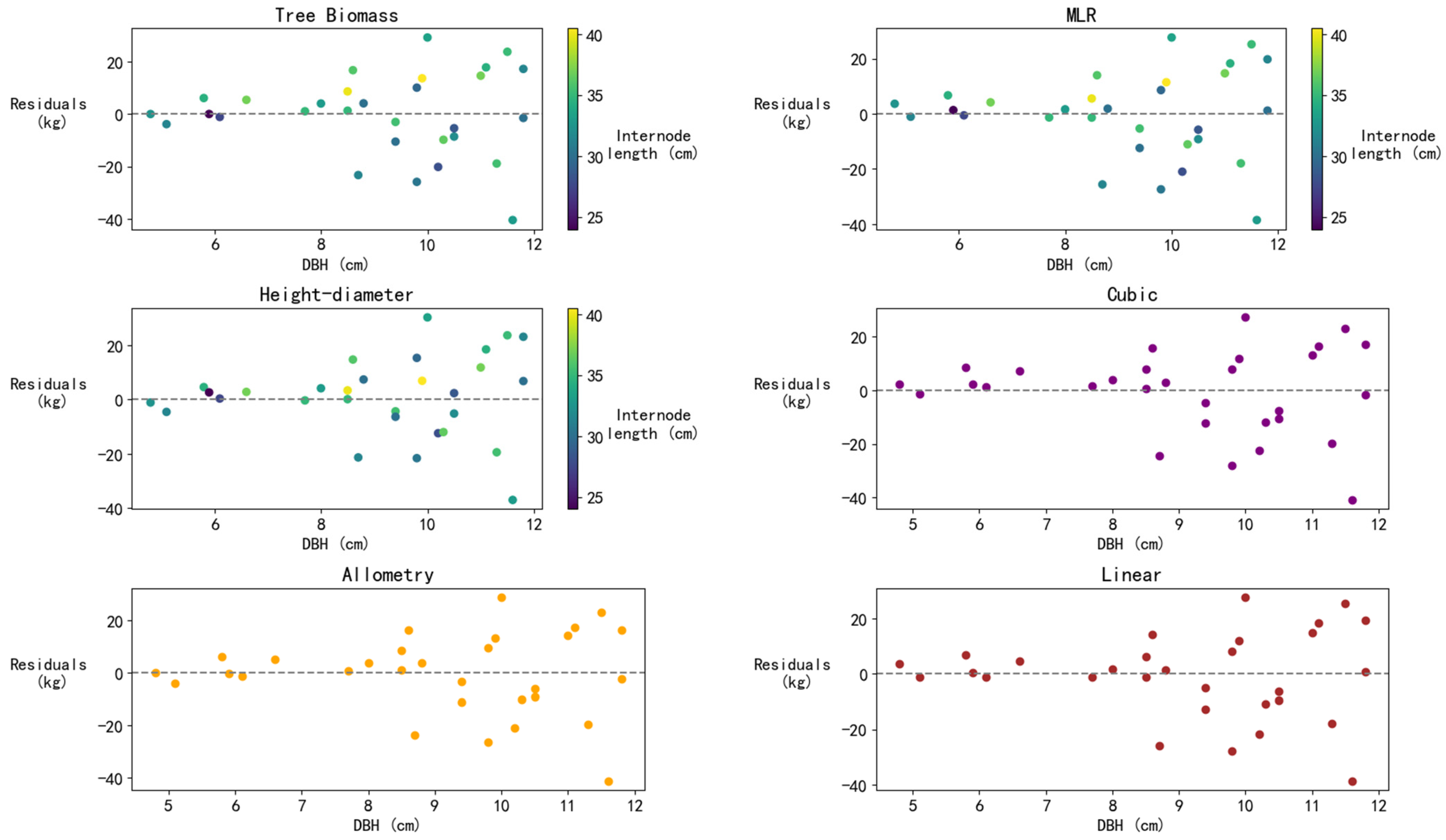
3.3.3. Residual Analysis of Models
4. Discussion
4.1. Data
4.2. Model Building Method
4.3. Comparison of Model Optimization Results
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zianis, D.; Muukkonen, P.; Mäkipää, R.; Mencuccini, M. Biomass and Stem Volume Equations for Tree Species in Europe; Silva Fennica Monogr; Tammer-Paino Oy: Tampere, Finland, 2005. [Google Scholar]
- Yang, H.; Guo, Z.; Chu, X.; Man, R.; Chen, J.; Liu, C.; Tao, J.; Jiang, Y. Comment on “Impacts of Species Richness on Productivity in a Large-Scale Subtropical Forest Experiment”. Science 2019, 363, eaav9117. [Google Scholar] [CrossRef] [PubMed]
- Návar, J. A Stand-Class Growth and Yield Model for Mexico’s Northern Temperate, Mixed and Multiaged Forests. Forests 2014, 5, 3048–3069. [Google Scholar] [CrossRef]
- Jyoti Nath, A.; Das, G.; Das, A.K. Above Ground Standing Biomass and Carbon Storage in Village Bamboos in North East India. Biomass Bioenergy 2009, 33, 1188–1196. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, L.; Li, F. Developing Two Additive Biomass Equations for Three Coniferous Plantation Species in Northeast China. Forests 2016, 7, 136. [Google Scholar] [CrossRef]
- Esteban, J.; McRoberts, R.E.; Fernández-Landa, A.; Tomé, J.L.; Nӕsset, E. Estimating Forest Volume and Biomass and Their Changes Using Random Forests and Remotely Sensed Data. Remote Sens. 2019, 11, 1944. [Google Scholar] [CrossRef]
- Millan, M.; Bonnet, A.; Dauzat, J.; Vezy, R. Advancing Fine Branch Biomass Estimation with Lidar and Structural Models. Ann. Bot. 2024, 134, 455–466. [Google Scholar] [CrossRef]
- Lin, Y.; Jaakkola, A.; Hyyppä, J.; Kaartinen, H. From TLS to VLS: Biomass Estimation at Individual Tree Level. Remote Sens. 2010, 2, 1864–1879. [Google Scholar] [CrossRef]
- Brown, S.; Gillespie, A.J.R.; Lugo, A.E. Biomass Estimation Methods for Tropical Forests with Applications to Forest Inventory Data. For. Sci. 1989, 35, 881–902. [Google Scholar] [CrossRef]
- Jenkins, J.C. Comprehensive Database of Diameter-Based Biomass Regressions for North American Tree Species; United States Department of Agriculture, Forest Service, Northeastern Research Station: Amherst, MA, USA, 2004. [Google Scholar]
- Seelan, S.K.; Laguette, S.; Casady, G.M.; Seielstad, G.A. Remote Sensing Applications for Precision Agriculture: A Learning Community Approach. Remote Sens. Environ. 2003, 88, 157–169. [Google Scholar] [CrossRef]
- Wang, S.; Lin, S.; Li, W.; Ding, Y. Variations in Fiber Morphology and Chemical Components of Dendrocalamus giganteus. For. Prod. J. 2016, 66, 319–325. [Google Scholar] [CrossRef]
- Wu, Y.; Cao, X.; Zhang, X.; Wei, J.; Xiong, Z.; You, J.; Gan, S.; Chen, D.; Zhang, H. The Influence Analysis of Bamboo Stand Structure on the Bamboo Shoot Yield of Pleioblastus macutatus. J. Sichuan For. Sci. Technol. 2019, 40, 55–59. [Google Scholar]
- Yen, T.; Lee, J. Comparing Aboveground Carbon Sequestration between Moso Bamboo (Phyllostachys heterocycla) and China Fir (Cunninghamia lanceolata) Forests Based on the Allometric Model. For. Ecol. Manag. 2011, 261, 995–1002. [Google Scholar] [CrossRef]
- Eggleston, H.S.; Buendia, L.; Miwa, K.; Ngara, T.; Tanabe, K. (Eds.) 2006 IPCC Guidelines for National Greenhouse Gas Inventories; Institute for Global Environmental Strategies (IGES): Hayama, Japan, 2006. [Google Scholar]
- Yen, T.; Sun, P.; Li, L. Predicting Aboveground Biomass and Carbon Storage for Ma Bamboo (Dendrocalamus latiflorus Munro) Plantations. Forests 2023, 14, 854. [Google Scholar] [CrossRef]
- Yen, T.; Ji, Y.; Lee, J. Estimating Biomass Production and Carbon Storage for a Fast-Growing Makino Bamboo (Phyllostachys makinoi) Plant Based on the Diameter Distribution Model. For. Ecol. Manag. 2010, 260, 339–344. [Google Scholar] [CrossRef]
- Huang, D.; Huang, D.; Wei, L.; Li, L.; Xu, Z. Culm Form Structure and Biomass of Bambusa chungii. World Bamboo Ratt. 2020, 18, 28–32. [Google Scholar]
- Hou, Y.; Yao, W.; Yang, G.; Cui, Y.; Zhou, H.; Zhang, X. Study on the Culm Structure and Allocation of Aboveground Biomass of Dendrocalamus tsiangii. J. Bamboo Res. 2023, 42, 51–57. [Google Scholar]
- Peng, Y.; Fan, S.; Su, W.; Xia, P. Distribution Pattern of Above-Ground Biomass and Culm Form Characteristics of Bambusa blumeana. J. Sichuan Agric. Univ. 2013, 31, 296–301. [Google Scholar]
- Zheng, Y.; Xu, Z.; Jia, D.; Li, Z. Study on Allocation of Aboveground Biomass and Optimal Model of Dendrocalamus latiflorus Shoot-Producing. J. Bamboo Res. 2023, 42, 23–29. [Google Scholar]
- Lin, D.; Zhao, H.; Li, Z.; Huang, C.; Xu, W. The Distribution Characteristics and Regression Model of Aboveground Biomass of Pleioblastus amarus in Nanling Mountain, Gangdong. Chin. J. Ecol. 2024, 43, 3205–3210. [Google Scholar]
- Kaushal, R.; Islam, S.; Tewari, S.; Tomar, J.M.S.; Thapliyal, S.; Madhu, M.; Trinh, T.L.; Singh, T.; Singh, A.; Durai, J. An Allometric Model-Based Approach for Estimating Biomass in Seven Indian Bamboo Species in Western Himalayan Foothills, India. Sci. Rep. 2022, 12, 7527. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Zhou, H.; Yan, Y.; Hou, Y.; Yang, Y.; Xie, T. Culm Structure and Above-Ground Biomass Allocation of Dendrocalamus farinosus and Bambusa rigida. Guizhou For. Sci. Technol. 2023, 51, 39–43. [Google Scholar]
- Yuen, J.; Fung, T.; Ziegler, A.D. Carbon Stocks in Bamboo Ecosystems Worldwide: Estimates and Uncertainties. For. Ecol. Manag. 2017, 393, 113–138. [Google Scholar] [CrossRef]
- Kong, F.; Zhong, Q.; Yu, L.; Kuang, X.; Huang, S.; Lu, Y.; Li, Y.; Gao, F. Regression Model of Biomass of Phyllstachys edulis at Different Age Classes in Ruijin, Jiangxi Province. South China For. Sci. 2022, 50, 39–42. [Google Scholar]
- Shi, Z.; Hui, C.; Zhang, J.; Wu, C.; Qin, Y. Research on Chemical Composition and Fiber Morphology of Dendrocalamus brandisii. Biomass Chem. Eng. 2009, 43, 21–24. [Google Scholar]
- Huang, D.; Li, L.; Huang, Y.; Lin, J.; Lu, Z.; Wei, C. Investigation on Germplasm Resources of Shoot-Producing Bamboo Species in Guangxi and Their Utilization. World Bamboo Ratt. 2024, 22, 48–53. [Google Scholar]
- Ma, S.; Ouyang, W.; Liu, X.; Han, M.; Yang, Y.; Guo, A. Physicochemical Properties of Dietary Fiber from Dendrocalamus brandisii Leaves. Food Res. Dev. 2021, 42, 1–7. [Google Scholar]
- Hui, C.; He, H.; Liu, W.; Shi, Z.; Sun, S. Promoting the Development of Dendrocalamus brandisii Industry for Rural Vitalization. World Bamboo Ratt. 2022, 20, 66–69. [Google Scholar]
- Tan, H.; He, B.; Tan, R.; Zhou, T. Technical Approaches for Building High-Yield and High-Efficiency Bamboo Forest Models for Sweet Bamboo. For. Sci. Technol. 2000, 32–33. [Google Scholar]
- Jianwei, L.; Wanling, Q.; Chaomao, H.; Qian, C.; Yumeng, W.; Shuhong, Z.; Xiuting, Z.; Weiyi, L. Geographical Variation in Quality of Dendrocalamus brandisii Bamboo Shoots and Its Relationship with Site Conditions. Sci. Silvae Sin. 2024, 60, 75–83. [Google Scholar]
- Yang, F.; Tang, M. Relationship between Site and Structure and Its Influence on Biomass in Phyllostachys edulis Forest. J. Zhejiang A&F Univ. 2020, 37, 823–832. [Google Scholar]
- Ji, X.; Luo, Q.; Ding, Y.; Wang, Y.; Zhao, J.; Wang, S. A Study on the Aboveground Biomass Model of Dendrocalamus brandisii. J. Bamboo Res. 2015, 34, 49–53. [Google Scholar]
- Zhang, X.; Zhang, F. A Study on the “Hollow Design” of Chinese Bamboo Product. In Proceedings of the 2008 9th International Conference on Computer-Aided Industrial Design and Conceptual Design, Kunming, China, 22–25 November 2008; pp. 729–733. [Google Scholar]
- Casas, L.F.; Aldana, A.M.; Henao-Diaz, F.; Villanueva, B.; Stevenson, P.R. Specific Gravity of Woody Tissue from Lowland Neotropical Plants: Differences Among Forest Types. Ecology 2017, 98, 1474. [Google Scholar] [CrossRef] [PubMed]
- Ghale, K.; Yadav, S.K.; Thapa, N.; Sah, J.N. Potentiality of Bamboos to Combat Climate Change Impacts and Biodiversity Conservation in the Annapurna Conservation Area, Nepal. J. For. Nat. Resour. Manag. 2020, 2, 79–91. [Google Scholar] [CrossRef]
- Ordóñez-Prado, C.; Tamarit-Urias, J.C.; Nava-Nava, A.; Rodríguez-Acosta, M. Additive Equations System to Estimate Aboveground Biomass by Structural Component and Total of Three Giant Bamboo Species in Mexico. CERNE 2024, 30, e-103267. [Google Scholar] [CrossRef]
- Li, Z.; Guan, F.; Zhou, X.; Liu, L.; Fu, D.; Zhang, X.; Li, M. Effect of Fertilization on Soil Fertility and Individual Stand Biomass in Strip Cut Moso Bamboo (Phyllostachys edulis) Forests. Forests 2024, 15, 252. [Google Scholar] [CrossRef]
- Xayalath, S.; Hirota, I.; Tomita, S.; Nakagawa, M. Allometric Equations for Estimating the Aboveground Biomass of Bamboos in Northern Laos. J. For. Res. 2019, 24, 115–119. [Google Scholar] [CrossRef]
- Etherington, T.R. Mahalanobis Distances for Ecological Niche Modelling and Outlier Detection: Implications of Sample Size, Error, and Bias for Selecting and Parameterising a Multivariate Location and Scatter Method. PeerJ 2021, 9, e11436. [Google Scholar] [CrossRef] [PubMed]
- Osewe, E.O.; Dutcă, I. The Effects of Combining the Variables in Allometric Biomass Models on Biomass Estimates over Large Forest Areas: A European Beech Case Study. Forests 2021, 12, 1428. [Google Scholar] [CrossRef]
- Zeng, W.; Tang, S. Goodness Evaluation and Precision Analysis of Tree Biomass Equations. Sci. Silvae Sin. 2011, 47, 106–113. [Google Scholar]
- ArauÂjo, T.M.; Higuchi, N.; Carvalho, J.A., Jr. Comparison of Formulae for Biomass Content Determination in a Tropical Rain Forest Site in the State of ParaÂ, Brazil. For. Ecol. Manag. 1999, 117, 52. [Google Scholar]
- Nocedal, J.; Wright, S.J. Quasi-Newton Methods. In Numerical Optimization; Springer: New York, NY, USA, 2006; pp. 135–163. [Google Scholar]
- Chunquan, X.; Qihu, X.; Liping, L.; Xiao, H.; Yong, L.; Han, Z.; Lei, C.; Yuancai, L. Biomass Models with Breast Height Diameter and Age for Main Nativetree Species in Guangdong Province. Sci. Silvae Sin. 2019, 55, 97–108. [Google Scholar]
- De Viliers, P. The Theory and Application of Regression Analysis and the Least-Sqaures Principle. Sci. Mil. South Afr. J. Mil. Stud. 2012, 7. [Google Scholar] [CrossRef]
- Hongwei, J.; Jielai, X. Partial Least Square and Its Application. J. Fourth Mil. Med. Univ. 2003, 24, 280–283. [Google Scholar]
- Berahas, A.S.; Jahani, M.; Richtárik, P.; Takáč, M. Quasi-Newton Methods for Machine Learning: Forget the Past, Just Sample. Optim. Methods Softw. 2022, 37, 1668–1704. [Google Scholar] [CrossRef]
- Wei, H.; Wei, X. Research on Surveying and Mapping Data Processing Based on Nonlinear Mathematical Models and Deep Learning Optimization. Scalable Comput. Pract. Exp. 2024, 25, 2260–2265. [Google Scholar] [CrossRef]
- Adediran, E.; Ameen, S. Machine Learning and Optimization Techniques for Solving Inverse Kinematics in a 7-DOF Robotic Arm. arXiv 2024, arXiv:2406.13064. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Name | Model Formula |
|---|---|
| Tree Biomass Model | |
| Relative Growth Model | |
| MLR | |
| Height–Diameter | |
| Schumacher–Hall | |
| Quadratic | |
| Cubic | |
| Compound | |
| Allometric | |
| Linear |
| Model Name | Model Formula |
|---|---|
| Tree Biomass Model | |
| Relative Growth Model | |
| MLR | |
| Height–Diameter | |
| Schumacher–Hall | |
| Quadratic | |
| Cubic | |
| Compound | |
| Allometric | |
| Linear |
| Model Name | Model Formula |
|---|---|
| Tree Biomass Model | |
| Relative Growth Model | |
| MLR | |
| Height–Diameter | |
| Schumacher–Hall | |
| Quadratic | |
| Cubic | |
| Compound | |
| Allometric | |
| Linear |
| Model Name | R2 | SEE | TRE | MSE | MPE | MPSE | TRE * | MSE * |
|---|---|---|---|---|---|---|---|---|
| Tree Biomass Model | 0.745 | 10.6 | −0.71 | −3.16 | 6.29 | 3.92 | 5.35 | 0.34 |
| MLR | 0.737 | 10.7 | 1.76 × 10−6 | 2.79 | 6.32 | 54.93 | 4.01 | −1.30 |
| Height–Diameter | 0.692 | 11.5 | 5.21 × 10−6 | −3.11 | 6.80 | 26.17 | 8.71 | −3.11 |
| Cubic | 0.753 | 10.4 | −5.19 × 10−5 | 0.24 | 6.09 | 24.06 | 3.87 | −0.44 |
| Allometric | 0.745 | 10.5 | −0.71 | −3.13 | 6.19 | 23.97 | 5.32 | 0.30 |
| Linear | 0.736 | 10.7 | −2.93 | 0.72 | 6.30 | 50.15 | 3.84 | −1.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Zeng, W.; Guo, L.; Xu, Z.; Fan, S.; Cai, C.; Hui, C.; Liu, W. Construction and Comparison of Single-Tree Biomass Model for Dendrocalamus brandisii. Forests 2025, 16, 301. https://doi.org/10.3390/f16020301
Wang Z, Zeng W, Guo L, Xu Z, Fan S, Cai C, Hui C, Liu W. Construction and Comparison of Single-Tree Biomass Model for Dendrocalamus brandisii. Forests. 2025; 16(2):301. https://doi.org/10.3390/f16020301
Chicago/Turabian StyleWang, Zuming, Weisheng Zeng, Lei Guo, Zhihong Xu, Shaohui Fan, Chunjun Cai, Chaomao Hui, and Weiyi Liu. 2025. "Construction and Comparison of Single-Tree Biomass Model for Dendrocalamus brandisii" Forests 16, no. 2: 301. https://doi.org/10.3390/f16020301
APA StyleWang, Z., Zeng, W., Guo, L., Xu, Z., Fan, S., Cai, C., Hui, C., & Liu, W. (2025). Construction and Comparison of Single-Tree Biomass Model for Dendrocalamus brandisii. Forests, 16(2), 301. https://doi.org/10.3390/f16020301