Abstract
Grafting serves as a pervasive methodology in the propagation of jujube plants, yet the nuanced molecular mechanisms that dictate rootstock-scion interactions remain inadequately understood. We examined the transcriptomic landscapes of jujube heterograft combinations. Contrary to self-grafting conditions, early-stage heterografting yielded no discernible advantageous effects on scion biomass accretion. Interestingly, the rootstock’s biomass was significantly impacted by the scion, varying by species. The differentially expressed genes (DEGs) across graft combinations were mainly enriched for the vegetative growth of rootstocks, secondary metabolism, and resistance improvement of scions. Weighted gene co-expression network analysis (WGCNA) identified 27 hub genes which were negatively correlated with plant growth and biomass enlargement, serving as negative regulators, while the genes, L484_001734, ATHB-15, and BPC1, were involved in positive regulation. With biomass measurements, the transcriptomic data supported that an incomplete vascular recovery during early grafting led to nutrient accumulation at the graft junction, temporarily limiting plant growth while providing development resources for callus. In summary, our work has demonstrated that the intricate biological connections between the rootstock and scion guarantee the effective jujube grafting process by elucidating the molecular processes involved in the process.
1. Introduction
The jujube (Ziziphus jujuba Mill.), also called Chinese date, is a significant part of the Rhamnaceae family and one of the oldest fruit trees still in cultivation worldwide [1]. It is a Chinese product with a long history that dates back to the Neolithic Period, about 7000 years ago [2]. The fruit is not only an integral part of Chinese agriculture but also holds a revered position in Chinese folklore [3]. Beyond its historical significance, jujube has gained contemporary relevance for its nutrient-dense fruits that are superior in terms of carbohydrates, Vitamin C, cyclic nucleotides, and zinc content to many other fruits [4,5]. Grafting serves as one of the most historically rooted techniques, as evidenced by ancient Chinese manuscripts such as Qi Min Yao Shu, which dates back to the Northern Wei Dynasty, approximately 1400 years ago [6,7]. Grafting is traditionally used in jujube propagation. To conserve its desirable horticultural traits and expedite fruit production, jujube is commonly propagated asexually by grafting [8]. Despite the long history and widespread practice of grafting in jujube cultivation, there exists a significant knowledge gap in understanding the molecular mechanisms that govern the symbiosis between rootstock and scion. Given that grafting has been demonstrated to have several effects on the phenotypic of grafted plants, such as changes in growth, stress tolerance, fruit quality, and resistance to pests and diseases, the lack of knowledge is especially startling [9]. Grafting has become an important method for developing resistance to biotic and abiotic stresses and increasing the quality and potential yield in agricultural and horticultural production [7]. Grafting commonly influences the phenotype of the grafted plants including plant growth and development, tolerance to adversity and stress, changes in fruit quality, resistance to pests and pathogens, and other physiological disorders [10,11]. Related studies have shown that interactions between the rootstock and the scion can change physiological and morphological characteristics [12,13]. In recent years, the study of scion-rootstock metabolic interaction of grafted grapevines has shown that the heterografting affected rootstocks more than scions [14]. Numerous studies in relevant domains have unequivocally shown that the scion influences the rootstock’s growth and development [15]. The rootstock promotes the growth of the scion, changes the resistance of the scion, and alters the synthesis of secondary metabolites of the scion [16,17,18,19,20,21]. It has also been reported that genetic material exchange takes place between rootstocks and scions [22,23,24,25].
The present study aims to fill this knowledge gap by employing high-throughput transcriptomic data analysis. For the experiment, we selected Ziziphus spina-christi (L.) Willd from the Gwadar region near the Arabian sea in southwestern Pakistan and Ziziphus jujuba Mill. var. spinosa from Zanhuang County in north-central Hebei Province of China for heterografting (Figure 1). The two species were selected due to their robust and resistant characteristics, including tolerance to a range of environmental stressors such as heat, salt, and drought. Especially, both species have important ecological and economic value in greening barren and saline alkali land, as well as preventing and controlling desertification, and the fruits have high nutritional and medicinal value [26,27]. The choice of materials provides a unique opportunity to delve into the complex mechanisms governing rootstock-scion interactions, an area less explored in jujube [3,6,28,29,30,31,32,33].

Figure 1.
Source of Grafting Materials. The grafting experiments were carried out using Ziziphus spina-christi (L.) Willd. from Gwadar port area in southwestern Pakistan near the Arabian Sea and Ziziphus jujuba Mill. var. Spinosa from Zanhuang County, Hebei Province, north-central China. (a) Jujube of Ziziphus jujuba Mill. var. spinosa. (b) Geographical location of the source of experimental materials, the green dot represents the location of the Gwadar port area in Pakistan, and the red dot represents the location of Hebei Province in China. (c) Jujube of Ziziphus jujuba Mill. var. Spinosa. (d) Fruit of Ziziphus spina-christi (L.) Willd. (e,f) Heterografting of Ziziphus spina-christi (L.) Willd. and Ziziphus jujuba Mill. var. spinosa. (g) Fruit of Ziziphus jujuba Mill. var. spinosa.
By employing heterograft combinations between the two species as our experimental model, we sought to identify the role of negative regulators in the post-grafting phase and understand the modulation of key metabolic pathways. Utilizing the Illumina platform for de novo sequencing, we constructed an extensive unigene library and performed comparative analysis to identify DEGs between the rootstocks and scions, thereby paving the way for a nuanced understanding of their interactive molecular dynamics. This transcriptomic-based approach aims to offer an invaluable roadmap for future research endeavors, particularly targeting the functional characterization of candidate genes pivotal for optimizing grafting techniques in woody perennial crops.
2. Materials and Methods
2.1. Experimentation with Planting Materials and Grafting Techniques
Seeds from two Ziziphus species, namely Ziziphus jujuba Mill. var. Spinosa and Ziziphus spina-christi (L.) Willd, were collected for the study. Seeds of Ziziphus jujuba Mill. var. Spinosa were obtained from a plantation in Zanhuang County, Shijiazhuang City, Hebei Province, China. Seeds of Ziziphus spina-christi (L.) Willd. were obtained from Gwadar Port, Gwadar City, Balochistan Province, Pakistan. Both sets of seeds originated from the same maternal tree. The study was performed from 2019 to 2022 at the Laboratory of Cultivation and Protection for Non Wood, College of Forestry, Central South University of Forestry and Technology in Changsha, China (Latitude 28°12′ N, Longitude 112°59′ E).
Seeds from both species were sown in seedling trays measuring 6 cm × 2.2 cm × 11 cm (Length × Width × Height), filled with a growing medium consisting of peat moss and perlite in a 3:1 ratio. These trays were then placed in an artificial growth chamber with a temperature range of 22–25 °C. Upon the unfurling of the first true leaf, seedlings were subjected to split grafting techniques.
Four different grafting combinations were investigated, denoted as follows (Table 1): Two heterografted combinations and two homografted combinations were constructed: (1) S/G: Ziziphus jujuba Mill. var. Spinosa (scion)/Ziziphus spina-christi (L.) Willd. (rootstock), (2) G/S: Ziziphus spina-christi (L.) Willd. (scion)/Ziziphus jujuba Mill. var. Spinosa (rootstock), (3) S/S: Ziziphus jujuba Mill. var. Spinosa (scion)/Ziziphus jujuba Mill. var. Spinosa (rootstock), and (4) G/G: Ziziphus spina-christi (L.) Willd. (scion)/Ziziphus spina-christi (L.) Willd. (rootstock). Thirty days post-grafting, seedlings that exhibited similar growth characteristics were randomly chosen as samples. Each grafting treatment was replicated three times with ten seedlings per replicate. Stems and leaves 1 cm above the graft junction (as the scion material after grafting) and rhizomes 1 cm below the junction (as the rootstock material after grafting) were sampled for three biological replicates at 30 days after grafting. Three grafted seedlings were chosen at random from each repetition after 30 days of grafting, washed with water, and then patted dry with kitchen paper. With homologous grafting as a control, the following comparative analysis was carried out.

Table 1.
Designs of heterografted combinations and homografted combinations.
2.2. Determination and Analysis of Phenotypic Indexes of Grafted Seedlings
Scanned images of the grafted combinations were employed for the calculation of biomass statistics. Tree growth was ascertained using Epson Scan (Expression 11000XL, Skeko Epson Corp., Tokyo, Japan) for scanning the samples. Subsequent analysis was conducted using WinRHIZO PRO 2013 (Regent Instruments, Quebec, QC, Canada) and electronic balances to measure scion weight, length, diameter, and projected area, as well as rootstock length, diameter, projected area, surface area, average diameter, and root volume.
2.3. Transcriptome Sequencing and Data Analysis
At 30 days after grafting, samples that were healthy and devoid of diseases and pests were selected, and approximately 3 g was prepared for transcriptome analyses (at least three biological repeats). Scions and rootstocks were harvested from three independent plants per grafting combination and immediately preserved in liquid nitrogen. Transcriptome sequencing of grafted samples was performed by Beijing Biomarker Biotech Co., Ltd. Total RNA was extracted and the transcriptome assembly library was constructed. The Illumina Hiseq system (Illumina NovaSeq 6000 platform, San Diego, CA, USA) was used to determine RNA integrity and quality, for library construction, and cluster processing and sequencing. Clean reads were assembled using Trinity software (v2.5.1) [34], DIAMOND(v2.0.4) [35], KOBAS (v3.0) [36], Interpro (v5.34-73.0) and HMMER software packages (v3.1b2) which were used for unigene annotation. Bowtie (v2.3.5.1) was used to compare the reads obtained by sequencing with the unigene library [37]. The expression level was assessed in conjunction with RSEM (v1.2.19), per the comparison findings [38]. The FPKM value was used to represent the expression abundance of the corresponding unigene. For analysis of the DEGs, DESeq2 was used to analyze the differential expression between the sample groups, and the differentially expressed gene set between the two conditions was obtained [39]. Utilizing the Benjamini-Hochberg approach, the significant p-value derived from the initial hypothesis test was corrected during the differential expression analysis procedure. Lastly, the essential metric for identifying differentially expressed genes was the corrected p value, or FDR (False Discovery Rate). FDR < 0.01 and the difference multiple FC (Fold Change) more than or equal to 2 were the screening criteria applied during the procedure. Based on the findings, a Venn diagram representing DEGs was created. Additionally, BMKCloud (www.biocloud.net) was utilized to classify and enrich the DEGs using the Kyoto Encyclopedia of Genes and Genomes (KEGG).
2.4. Gene Validation and Expression Analysis
Actin was chosen as an internal reference gene, and sixteen unigenes associated with Multi-Pathway Analysis of Differentially Expressed Genes in Plant Graft Combinations were chosen for qPCR analysis in order to confirm the correctness of the transcriptome data. Primers software version 5 was used to build specific primers (Supplementary Table S14). As directed by the manufacturer of the 2 × SYBR Green Master Mix Enzyme kit (Biotool, Houston, TX, USA), the qPCR reaction system was set up. The steps involved in PCR amplification were predenaturation at 95 °C for 5 min, 40 cycles of 95 °C for 15 s each, and annealing at 60 °C for 40 s. Dissolution curves were recorded with 0.5 °C increments every 5 s, ranging from 60 °C to 95 °C. Three repetitions of each response were made. The 2−ΔΔCq technique was used to determine the target genes’ relative expression levels. Using SPSS version 17.0 software (SPSS Inc., Chicago, IL, USA), graphs were generated using GraphPad Prism (version 8.0.0 for Windows, GraphPad Software, San Diego, CA USA, www.graphpad.com), while correlation analysis was carried out in accordance with the gene’s FKPM value and relative expression.
2.5. Screening of Key Structural Genes in Multiple Pathways
A heat map was created using BMKCloud (www.biocloud.net) based on the FPKM values of the DEGs in the pathways. Following that, the KEGG pathway map was used to create an expression map of the DEGs. Screening of the important genes was performed after a thorough analysis of the expression rules was conducted using the expression map.
2.6. Construction of Gene Co-Expression Networks via WGCNA
The Weighted Gene Co-expression Network Analysis (WGCNA) R package (v4.3.2) was employed to construct gene co-expression networks. The parameters for the analysis were set as follows: powerEstimate at 16, maxBlockSize at 25,000, minModuleSize at 30, and mergeCutHeight at 0.25. A module—phenotype correlation heatmap was created using the ggplot2 package in R (v4.3.2). Within the “green” module, hub genes were selected based on GS1 and datKME values that exceeded 0.85.
3. Results
3.1. Phenotypic Variability and Influences across Grafting Combinations
To offer a nuanced, comprehensive understanding of the phenotypic variations and interactions between scion and rootstock in jujube plants when subjected to different grafting combinations, homograft combinations were used as controls. This exploration is encapsulated in (Figure 2a–x and Supplementary Table S1), which serves as a robust data portal featuring various growth parameters and biomass attributes. Our integrated analysis divulges a complex relationship between scion and rootstock, substantiated by multiple measurements including diameter, length, projected area, and weight. Specifically, the diameter of the scion was markedly greater in heterografts (G/S_scion = 1.74 mm ± 0.24, S/G_scion = 1.34 mm ± 0.03) as opposed to homografts (G/G_scion = 1.44 mm ± 0.01, S/S_scion= 1.25 mm ± 0.01). Conversely, compared with homologous grafting, scion length, projected area, and weight did not show a significant advantage in heterografts across the grafting categories.
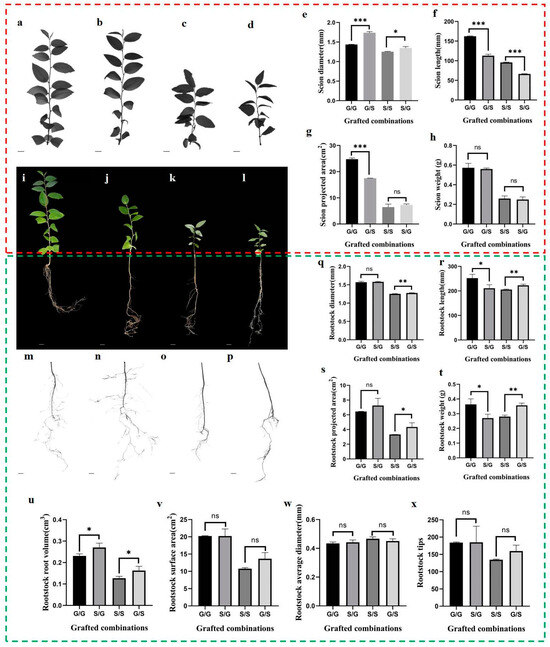
Figure 2.
Early effects of heterograft in comparison with homograft. (a–d) Scanning of G/G_scion, G/S_scion, S/S_scion, S/G_scion. Scale bars, 1 cm. (e–h) Effect of heterograft on scion diameter, length, projected area, and weight compared with homograft. (i–l) Growth of grafted seedlings, G/G, G/S, S/S, S/G. Scale bars, 1 cm. (m–p) Scanning of G/G_rootstock, S/G_rootstock, S/S_rootstock, G/S_rootstock. Scale bars, 1 cm. (q–x) Effect of heterograft on rootstock diameter, length, projected area, and weight compared with homograft. Statistical significance was determined by Student’s t-test and the data are shown as mean ± standard deviation (n = 3). (Significance codes: *** represents significant at a p < 0.001 threshold, ** represents significant at a p < 0.01 threshold, * represents significant at a p < 0.05 threshold, ns represents no significance).
As for the rootstock, we observed that the diameter, length, projected area, and weight of the rootstock in G/S heterografts (1.28 mm ± 0.01, 222.70 mm ± 3.82, 4.35 cm2 ± 0.46, 0.36 g ± 0.01) surpassed that in S/S homografts (1.25 mm ± 0.01, 205.60 mm ± 1.42, 3.33 cm2 ± 0.01, 0.28 g ± 0.01), and the root volume of the rootstock was markedly greater in heterografts (S/G_rootstock = 0.27 cm3 ± 0.02, G/S_rootstock = 0.16 cm3 ± 0.02) as opposed to homografts (G/G_rootstock= 0.23 cm3 ± 0.01, S/S_rootstock = 0.13 cm3 ± 0.01). However, additional parameters—including rootstock surface area, average diameter, root volume, and root tip number—did not show significant differences across the grafting categories.
These observations suggest that while the rootstock does influence the growth of the scion, particularly in diameter, it does not uniformly affect other attributes. Yet, the G scion uniformly promotes the growth of the S rootstock in terms of diameter, length, projection area, root volume, and weight. Thus, our findings underscore the multi-dimensional impact of grafting combinations on the developmental trajectory of both the scion and rootstock in jujube plants.
3.2. Transcriptomic Analysis of Grafted Jujube Combinations
To study the genetic factors influencing grafted jujube development, we analyzed the transcriptomes of Ziziphus jujuba Mill. var. spinosa and Ziziphus spina-christi (L.) Willd across various graft combinations, using self-grafted plants as controls. Over 30 days, the G/S heterograft rootstock exhibited significant growth advantages in weight and length compared to the S/S homograft control. Notably, the effect of the Ziziphus spina-christi (L.) Willd scion was more pronounced than the Ziziphus jujuba Mill. var. spinosa scion. For RNA-seq analysis, a cDNA library was constructed from RNA isolated from the different rootstock and scion combinations. Library sequencing was performed on the Illumina platform, generating 24 post-QC samples totaling 202.76 Gb of clean data. De novo assembly produced 41,574 unigenes with an N50 length of 2218 bp. Over 70% of unigenes were annotated across eight major databases (Supplementary Table S2), primarily aligning to Ziziphus jujuba in Nr (Supplementary Figure S1a). Approximately 23,502 annotated unigenes were categorized into three groups based on their match in the GO database: molecular function, biological process (4918, 19.8%), and cellular component (6530, 26.3%). The largest GO terms were “binding”, “catalytic activity”, and “structural molecular activity” for molecular function; “cell”, “cell part”, and “organelle” for cellular component; and “metabolic process”, “cellular process”, and “single-organism process” for biological process (Supplementary Figure S1b). Approximately 23502 unigenes were categorized into 31 GO terms and 137 KEGG pathways (Supplementary Tables S3 and S4), encompassing a comprehensive range of ontologies and pathways.
3.3. Comprehensive Differential Expression Analysis Illuminates Scion-Rootstock Interactions in Grafted Jujube Plants
In an endeavor to understand the genes potentially influencing scion-rootstock dynamics, a differential expression analysis was simultaneously carried out across multiple graft combinations and tissues. Intriguingly, the minimal number of DEGs was observed between G/G_scion vs. G/S_scion and S/S_scion vs. S/G_scion. This stands in stark contrast to G/G_rootstock vs. S/G_rootstock and S/S_rootstock vs. G/S_rootstock, which manifested 5159 and 1204 DEGs, respectively—figures which are significantly higher than that for grafted scions. Cumulatively, the analysis yielded approximately 6363 DEGs (Supplementary Figure S2a), and we analyzed the DEGs of the G/G_scion vs. G/S_scion, S/S_scion vs. S/G_scion, G/G_rootstock vs. S/G_rootstock and S/S_rootstock vs. G/S_rootstock using Venn diagrams (Supplementary Figure S2b). We found that 507 DEGs were shared in G/G_rootstock vs. S/G_rootstock and S/S_rootstock vs. G/S_rootstock (Supplementary Figure S2c), and 24 DEGs were shared in G/G_scion vs. G/S_scion and S/S_scion vs. S/G_scion (Supplementary Figure S2d). To substantiate the integrity of our RNA-Seq findings, we employed quantitative RT-PCR (RT-qPCR) validation on a random subset of 16 genes, featuring diverse expression levels. RNA from the scions and rootstocks of S/G, G/S, S/S, and G/G grafts served as the templates for this verification process. The RT-qPCR outcomes were largely in alignment with our RNA-Seq data, thereby validating the robustness of our transcriptomic analysis (Supplementary Figure S3).
In the comparison between G/G_rootstock and S/G_rootstock as well as between S/S_rootstock and G/S_rootstock, the majority of DEGs in these heterograft combinations were predominantly involved in pathways related to “Carbon Metabolism”, “Photosynthesis-Antenna Proteins”, “Pentose and Glucuronate Interconversions”, and “Phenylpropanoid Biosynthesis”. Similarly, in the comparative analyses of S/S_scion vs. S/G_scion and G/G_scion vs. G/S_scion, the DEGs were primarily mapped to pathways including “Phenylpropanoid Biosynthesis”, “Flavonoid Biosynthesis”, “Plant-Pathogen Interaction”, “MAPK Signaling Pathway”, “Zeatin Biosynthesis”, “Ubiquinone and Other Terpenoid-Quinone Biosynthesis”, “Cyanoamino Acid Metabolism”, “Cutin, Suberine, and Wax Biosynthesis”, as well as “Stilbenoid, Diarylheptanoid, and Gingerol Biosynthesis” (Supplementary Figure S4).
3.4. Multi-Pathway Analysis of Differentially Expressed Genes in Plant Graft Combinations
In this comprehensive study on graft combinations, multiple pathways integral to plant metabolism and stress resistance were examined, revealing a complex tapestry of DEGs. The pathways of “Carbon Metabolism” and “Pentose and Glucuronate Interconversion” were particularly salient in rootstocks, with 44 and 51 DEGs identified, respectively (Figure 3a,b, Supplementary Tables S5 and S6). These DEGs were notably upregulated in the G/S rootstock but demonstrated variable expression in the S/G scion. The genes encoding proteins such as HK, GPI, FBP, PFK, ALDO, among others, were predominantly upregulated in the G/S rootstock, while a subset (FPK, gapN, gpmA, ENO, etc.) was downregulated in the S/G scion. Such disparities underscore the necessity of further research to understand the role of these pathways in carbohydrate metabolism and their sensitivity to grafting combinations.
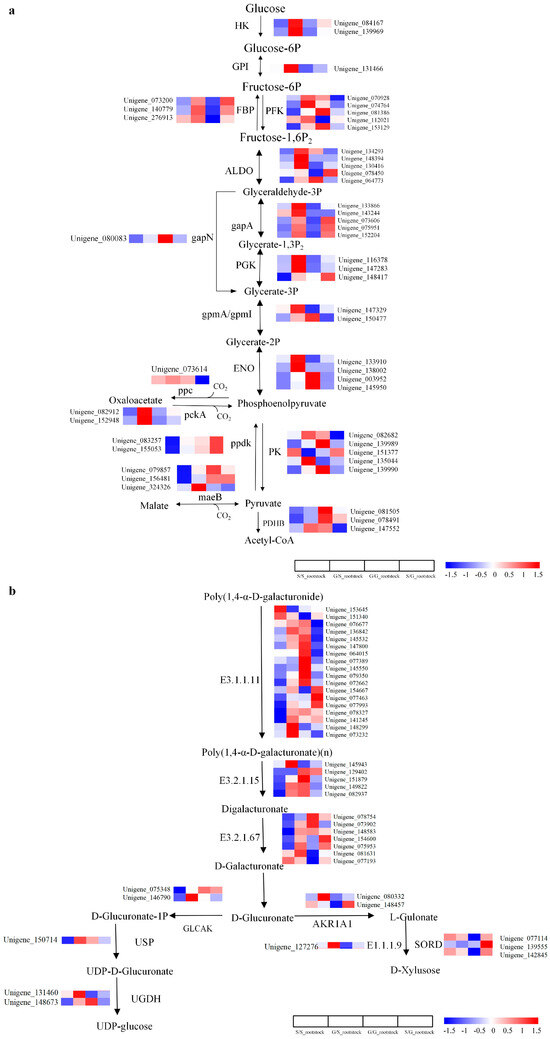
Figure 3.
Analysis of DEGs in the pathways of carbon metabolism and pentose and glucuronate interconversion in the rootstock. (a) DEGs in the pathway of carbon metabolism. (b) DEGs in the pathway of pentose and glucuronate interconversion. The log2-transformed FPKM (fragments per kilobase of exon per million mapped reads) values represented by color scale. High expression represented by red color, and low expression represented by blue color.
Focusing on pathways central to plant defense, namely “Phenylpropanoid Biosynthesis” and “Flavonoid Biosynthesis”, we observed significant activity in both the scions and rootstocks of heterografts. In the Phenylpropanoid Biosynthesis pathway, 77 DEGs were identified, with genes encoding proteins such as 4CL, CCR, and CAD being significantly upregulated in both heterograft scions and rootstocks, compared to homografts. Additionally, proteins like PAL, bglx, CYP73A, and others demonstrated nuanced expression patterns across different graft combinations, suggesting a highly regulated biosynthetic activity of phenylpropanoids. Similarly, 26 DEGs were detected in the Flavonoid Biosynthesis pathway. Genes encoding proteins like CYP73A, LAR, and ANR were uniformly upregulated in heterograft scions, whereas a more complex expression pattern emerged for proteins like CYP98A, HCT, and F3H depending on the graft combinations (Figure 4a,b, Supplementary Figure S5 and Tables S7 and S8). The results not only signify the potential role of these pathways in bolstering the plant’s defensive mechanisms but also underline the significance of graft combinations in modulating these critical biological activities.
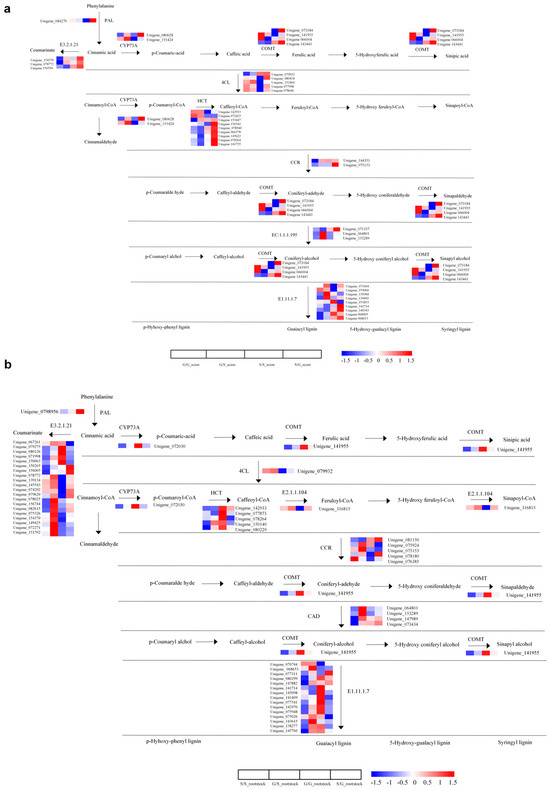
Figure 4.
Analysis of DEGs in the pathways of phenylpropanoid biosynthesis. (a) DEGs in the phenylpropanoid biosynthesis pathway of scions. (b) DEGs in the phenylpropanoid biosynthesis pathway of rootstocks. The log2-transformed FPKM (fragments per kilobase of exon per million mapped reads) values represented by color scale. High expression represented by red color, and low expression represented by blue color.
In terms of secondary metabolism, genes implicated in ubiquinone and other terpenoid-quinone biosynthesis, cyanoamino acid metabolism, stilbenoid, diarylheptanoid, and gingerol biosynthesis, and cution, suberine, and wax biosynthesis were predominantly upregulated in G/S and S/G scions as compared to homografts. Specifically, a marked upregulation was noted in S/G scions, indicating a graft-specific modulation of the biosynthetic pathways (Supplementary Figure S6a–d, Table S9). Additionally, genes associated with plant-pathogen interaction and MAPK signaling pathways were significantly altered. For instance, 61 DEGs were identified in plant-pathogen interaction and 33 DEGs in the MAPK signaling pathway (Supplementary Figures S7 and S8 and Tables S10 and S11). Noteworthy was the up-regulation of pathogenesis-related proteins and transcription factors like BAK1 and WRKY22 in both heterograft scions. Similarly, genes encoding proteins related to stress and defense responses, including calcium signaling and reactive oxygen species scavenging, exhibited differential expression profiles. Particularly, proteins such as CALM, MPK8, and RBOH were implicated in maintaining reactive oxygen species homeostasis, while PP2C and MAPKKK17_18 were involved in stress adaptation. On the hormonal regulation front, genes in the Zeatin biosynthesis pathway showed higher expression in both S/G and G/S scions relative to homografts, yet with a noteworthy downregulation in S/G scions (Supplementary Figure S9 and Table S12). These findings suggest that grafting significantly influences the molecular regulatory mechanisms governing plant growth, stress responses, and secondary metabolism.
3.5. Exploring Functional Modules and Hub Genes in Post-Grafting Processes
To gain a comprehensive understanding of the molecular underpinnings of post-grafting plants, particularly in the context of scion-induced resistance, an intricate analysis was executed. Employing differential gene expression analysis, a total of 22,717 DEGs were identified across all conditions. To further refine this extensive dataset, WGCNA was used, segregating these DEGs into 24 distinct functional modules. One module, termed the “green module 1886 genes” stood out for its pronounced negative correlation to phenotypic traits of plant height and project area (−0.78 and −0.74, respectively), signaling its potential role in disease resistance mechanisms (Figure 5).
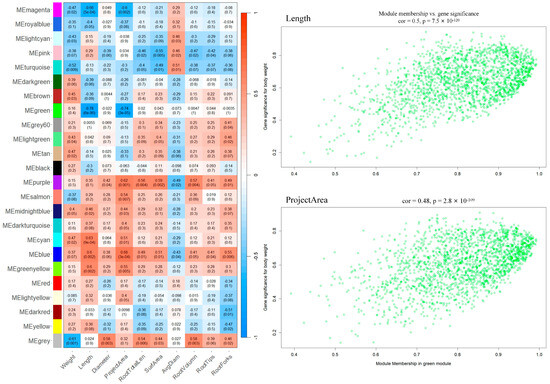
Figure 5.
WGCNA to explore transcriptional regulation and expression of the molecular underpinnings of post-grafting plants. The heatmap indicated the correlation between modules and phenotypic data. The WGCNA R package (v4.3.2) was employed to construct gene co-expression networks. The parameters for the analysis were set as follows: powerEstimate at 16, maxBlockSize at 25,000, minModuleSize at 30, and mergeCutHeight at 0.25. A module—phenotype correlation heatmap was created using ggplot2 package in R. Within the “green” module, hub genes were selected based on GS1 and datKME values that exceeded 0.85.
Within this green module, 27 hub genes were identified with GS1 and datKME values exceeding 0.85, indicating their centrality in the network (Figure 6 and Supplementary Table S13). The hub genes, such as those involved in transcriptional regulation (e.g., 080501, 079312, 144465, 149590, 145952) and transporter genes (e.g., 154560, 151878), cover a wide range of functional categories from modulating growth and development to regulating metabolite fluxes between the rootstock and the scion. Moreover, genes related to protein turnover, defense and stress responses, hormone signaling, carbohydrate metabolism, and secondary metabolism were also identified, each contributing to different aspects of plant growth, development, and physiology, particularly in grafting scenarios.
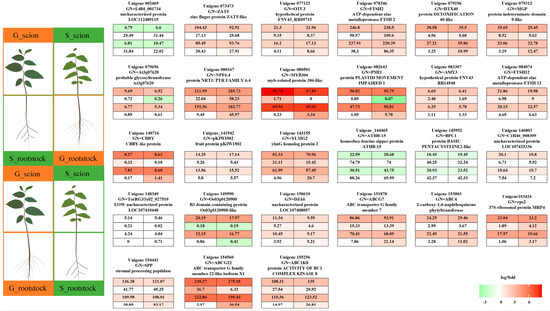
Figure 6.
Selection of hub genes. Within the “green” module, 27 hub genes were identified based on GS1 and datKME values that exceeded 0.85 in the post-grafting plants, indicating their centrality in the network. These range from modulating growth and development to regulating metabolite fluxes between the rootstock and the scion.
Additionally, it is worth noting that the majority of the hub genes were upregulated in the scion parts across both combinations studied, with the exception of genes 002469, 144465, and 145952, which were downregulated. The broad functional spectrum of these genes suggests a complex, multi-layered response to grafting, which combines various biological processes from transcription regulation and metabolite transport to stress response and hormone signaling. Understanding the precise roles of these genes could provide invaluable insights into the mechanisms that contribute to successful grafting and plant resistance.
4. Discussion
The intricate interplay between scions and rootstocks in agricultural systems has been a subject of extensive research, albeit with a predominant focus on the influence of rootstocks on scions [40,41,42,43]. Our comprehensive study, employing advanced transcriptomic data analysis, offers a creative, balanced perspective that illuminates the mutual influences between scions and rootstocks in Ziziphus species. Using advanced transcriptomic analysis, we identified critical regulatory networks governing diverse metabolic pathways, including carbohydrate metabolism and secondary metabolite biosynthesis. Notably, carbon metabolism and pentose/glucuronate interconversion genes were upregulated in heterografts.
Scions provide essential nutrients like sugars, hormones, and nucleic acids to rootstocks, significantly affecting carbohydrate storage and root growth [15,44,45,46,47]. At the end of the 20th century, farmers in Indonesia developed cassava grafting, using an inedible wild species as a scion to increase yields [48]. By grafting, the total yield and tuber size of cultivated cassava tubers increased by about two-fold [49]. Grafted scions also significantly increased the yield and quality of sweet potato and potato [50]. In tuber crops, the effect of rootstock on crop yield is particularly important. In fruit crops, the effect of scions on rootstocks has received less attention, which may be because the rootstock part grows underground and is difficult to be observed. However, the effect of scions on rootstocks may be significant, as the scion transports photosynthetic products, hormones, and nucleic acids for the rootstock, having a great impact on root growth [44]. Different apple scions affect the root growth rate, especially the main root length, diameter, and root weight [51]. The intergeneric grafting of Schisandraceae shows that the growth of rootstocks is inhibited by the scion [52].
This aligns with previous findings that stronger wild Ziziphus spina-christi (L.) Willd scions promoted rapid growth of Ziziphus jujuba Mill. var. spinosa rootstocks post-grafting. Meanwhile, compared to the control (S/S_rootstock), in the G/S_rootstock the DEGs participate in carbohydrate metabolism, such as ENO and pckaA, which is significantly upregulated expression. Likewise, in the grafting of peaches and apricots, the same upregulated DEGs in carbohydrate metabolism were also found to regulate plant growth after grafting [53]. This spotlight on rootstocks is noteworthy given that their underground location often precludes close observation, resulting in less attention in existing literature [40]. In contrast, rootstocks are commonly used in woody perennial crops to influence scion phenotypes [45].
Using the effect of the rootstock on the scion could control the height and shape of the plant, as well as the absorption and utilization of water and nutrients. For example, apple rootstocks have been developed for dwarfing apples and for improving mineral absorption and scion growth [54]. Rootstocks have been used to improve the growth of scion and improve water use efficiency in kiwifruit grafting [55]. Grafting can alter secondary metabolite accumulation, impacting scion growth, quality, and stress tolerance [18,56,57,58,59,60]. Strong rootstocks were used to enhance the resistance of scions, using rootstocks to enhance the disease resistance of potatoes, using pumpkin as rootstock to enhance the cold resistance of cucumber and watermelon scions, by upregulating the key genes in the carbon fixation, photosynthesis, and polyamine metabolism pathway [61,62]. By overexpressing the key gene, MdFRK2, of sugar metabolism in the apple rootstock, drought resistance was improved [63]. Different tomato rootstocks regulate ABA content and stomatal aperture by regulating ABA signaling pathway gene expression to adapt to drought stress [64]. Based on integrative multi-omics analysis, in grape hetero-grafting, rootstocks promote phenolic compound accumulation [16]. Transcriptomics and metabolomic analysis have found that the key genes, F5H and CCR, play a major role in lignin biosynthesis, so as to regulate the growth of winter jujube, improve the strength of cell walls, provide mechanical support for cells, and improve the resistance [65].
The same grafting regulation effect appeared in our study. Compared with the control, both Ziziphus jujuba Mill. var. spinosa and Ziziphus spina-christi (L.) Willd can promote each other’s scion diameter and root volume increase, and upregulated phenylpropanoid biosynthesis genes (PAL, COMT, 4CL, CCR, CAD, etc.), likely enhancing growth and inherent resistance.
By examining the reciprocal influences of metabolic pathways in both scion and rootstock, our work enriches the understanding of their complex interactions [13,66], providing insights to refine grafting practices for mutual benefits. Moreover, while the prevailing body of literature has largely concentrated on the influence of rootstocks on scion development, our investigation stands as a pioneering effort that employs RNA-Seq technology to identify an extensive array of genes affecting multiple pathways in both jujube scions and rootstocks. This work serves as a springboard for future research endeavors, which could leverage multi-omic methodologies to corroborate these gene expression observations and explore their functional ramifications in greater depth. However, it is important to acknowledge the limitations that constrain our study. The scarcity of existing research specifically targeted at jujube grafting hampers our ability to place our findings within a broader comparative framework. Additionally, although our study illuminates the impact on common metabolic pathways in both the scion and rootstock, this emphasis could inadvertently overshadow the importance of metabolic pathways that are unique to each component. Furthermore, the lack of concurrent secondary metabolite analyses in the scions introduces an element of speculation into our conclusions.
Our research also brings to the fore the substantial role that scions play in shaping rootstock characteristics. Notably, we observed a predominant upregulation of genes related to ABC transporters, specifically “ABC transporter G family member 22-like isoform X1” and “ABC transporter G family member 7”, in the scion following the grafting process. This upregulation suggests a potential enhancement in nutrient absorption and stress resilience, thereby contributing to the overall vigor of the grafted plant [67,68]. Additionally, we noted an upregulation in proteins involved in metalloprotease activities, such as “ATP-dependent zinc metalloprotease FTSH 2, chloroplastic”. This observation implies a likely improvement in cellular repair mechanisms and protein turnover, thereby reinforcing the mutualistic advantages inherent in the grafting process [69].
In the early stages of grafting, particularly within the first 30 days, our study posits that the upregulation of specific genes in the scion part enhances nutrient transport towards the rootstock. This is in line with findings from Habibi who discussed the molecular aspects of grafting affecting nutrient transport [19]. The upregulation appears to be a coordinated physiological response to the disruption of the vascular system [70,71]. ABC transporters (BMK_Unigene_154560, BMK_Unigene_151878, BMK_Unigene_080167), known for their role in nutrient uptake, are upregulated, corroborating Vibhuti’s findings on the role of upregulated genes in nutrient uptake [72]. Concurrently, genes related to chloroplast function (BMK_Unigene_078346, BMK_Unigene_084074, BMK_Unigene_155256) are also upregulated, potentially to meet the energy demands of enhanced nutrient transport, as suggested by Martinoia [73]. Transcription factors like myb-related protein 306-like (BMK_Unigene_080501) and zinc finger protein ZAT5-like (BMK_Unigene_073473) may act as regulatory nodes, similar to the regulatory roles of plant ABC transporters [74]. Additionally, enzymes such as glycosyltransferase At3g07620 (BMK_Unigene_079696) could be modifying nutrients for more efficient transport [75]. This orchestrated gene expression likely serves to facilitate nutrient mobilization to the rootstock, even when the vascular system is compromised. However, the incomplete vascular recovery during this period may result in nutrient accumulation at the grafting site, serving as a resource pool for callus growth, thereby temporarily impeding overall plant growth and biomass accumulation, a phenomenon also observed under abiotic stress conditions [76].
Our study identified both upregulated and downregulated genes post-grafting, which collectively form a complex tapestry of mechanisms affecting a wide array of physiological aspects, from nutrient transport to stress tolerance. Notably, the downregulated genes serve as negative regulators, adding a layer of complexity to the grafting process. In the initial stages of grafting, particularly within the first 30 days, nutrient accumulation at the grafting site may occur due to incomplete vascular recovery. This accumulation provides nutrition resources for callus growth, thereby temporarily impeding overall plant growth and biomass accumulation as mentioned above, and similar phenomena were also found in sweet cherries [77]. In this context, the downregulated genes act as negative regulators or “brakes”, slowing down certain metabolic pathways to prevent overexertion or resource depletion. This dynamic interplay ensures a balanced and sustainable growth pattern, optimizing mutual benefits for both the scion and the rootstock. These insights offer a launching pad for our future research, which could employ multi-omic strategies to validate the gene expression and delve deeper into their functional implications. As a result, the agricultural community moves one step closer to optimizing grafting practices that maximize crop yield, quality, and resilience, thereby making a substantive contribution to the global endeavor of sustainable agriculture. In summary, our findings not only validate but also build upon conventional wisdom concerning scion-rootstock interactions in Ziziphus species. The pathways we focused on, i.e., signal transduction, response to stimuli, secondary metabolite regulation, and nutrient growth, are in synchrony, emphasizing that grafting is not a one-dimensional procedure but a complex orchestration of biological processes. This multi-pathway approach confirms that grafting elicits a synergistic and multifaceted interaction, harmonizing various facets of plant growth and resilience. Future work can leverage these insights as a robust foundational framework, utilizing multi-omic approaches for a more complete understanding of these complex relationships. The inclusion of negative regulators and the concept of incomplete vascular recovery in this study have enriched our understanding and offered nuanced avenues for future research in the grafting of plants.
5. Conclusions
Contrary to self-grafting conditions, early-stage heterografting of Ziziphus yielded no discernible advantageous effects on scion biomass accretion. Interestingly, the rootstock biomass was significantly impacted by the scion, varying by species. Regarding he DEGs across graft combinations, pathways were mainly enriched in the vegetative growth of rootstocks, secondary metabolism, and resistance improvement of scions. During early grafting, incomplete vascular recovery led to nutrient accumulation at the graft junction, temporarily limiting plant growth while providing callus development resources. WGCNA identified 27 hub genes which were negatively associated with plant growth and biomass, serving as negative regulators, while the genes, L484_001734, ATHB-15, and BPC1, were involved in positive functions. By uncovering key mechanisms involving negative regulators, our study demonstrated the complex biological interactions enabling successful jujube grafting and the optimization of horticultural practices. This investigation has laid an important foundation for future research related to the biochemical interplay that governs scion-rootstock interactions in Ziziphus species.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/f15040618/s1, Figure S1: The annotation of unigenes; Figure S2: DEGs analysis of multiple graft combinations; Figure S3: Correlation analysis between qRT-PCR and RNA-seq; Figure S4: Enrichment analysis on KEGG pathways; Figure S5: Analysis of differentially expressed genes in the pathways of Flavonoid Biosynthesis in the scion; Figure S6: Analysis of differentially expressed genes in the pathway; Figure S7: Analysis of differentially expressed genes in the pathways of plant-pathogen interaction; Figure S8: Analysis of differentially expressed genes in the pathways of MAPK signaling; Figure S9: Analysis of differentially expressed genes in the pathways of Zeatin biosynthesis; Table S1: Growth parameters and biomass attributes of different grafting combinations; Table S2: Unigenes annotated across eight major databases; Table S3: Unigenes were categorized into 31 GO terms; Table S4: Unigenes were categorized into 137 KEGG pathways; Table S5: The DEGs in the pathway of Carbon Metabolism; Table S6: The DEGs in the pathway of Pentose and Glucuronate Interconversion; Table S7: The DEGs in the pathway of Phenylpropanoid Biosynthesis; Table S8: The DEGs in the pathway of Flavonoid Biosynthesis; Table S9: The DEGs in the pathway of ubiquinone and other terpenoid-quinone biosynthesis, cyanoamino acid metabolism, stilbenoid, diarylheptanoid and gingerol biosynthesis, cution, suberine and wax biosynthesis; Table S10: The DEGs in the pathway of plant-pathogen interaction; Table S11: The DEGs in the pathway of MAPK signaling pathway; Table S12: The DEGs in the pathway of Zeatin biosynthesis; Table S13: Annotations of 27 hub genes; Table S14: The qRT-PCR primer sequence information.
Author Contributions
S.Z.: designed, conducted the seedling, grafting, and laboratory experiments, completed the analysis of the data, completed the figures, tables and manuscript; S.S., J.P. and F.S.: participated in data analysis and provided advice on experiments. Z.L.: reviewed drafts of the manuscript. S.W.: provided funding, improved the research program, modified the manuscript. All the authors participated in the discussion and summary of the experiments results, and reviewed the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
For this study, we are very grateful to have the financial supports from the following programs or agencies: Central Financial Forestry Science & Technology Promotion Demonstration Fund project, Southern Fresh Jujube Cultivation Technology Standardization Demonstration Area Construction project (2023XT15), Ministry of Science & Technology of the People’s Republic of China 14th Five-Year National Key Research & Development Plan project, Jujube Precision Water & Fertilizer Control Key Technology Research project (2022YFD2200404), Research & Utilization of Tropical Arid Non wood Forest Germplasm Resources in China and Pakistan (2022WK2021), Screening of Drought & Heat Resistant Rootstocks and Rhizosphere Microbial Strains for Turpan Grape and Jujube (2023WK4008), Functional analysis and regulation mechanism of key genes in jujube embryo abortion (21B0255) and Study on the Mechanism of Interspecific Grafting Rootstock-scion Interaction in Jujube (CX20200707 and CX20201007).
Data Availability Statement
Data relevant to this study are publicly accessible and can be found at the following online repository: [NCBI] (https://www.ncbi.nlm.nih.gov/), with accession number PRJNA922324.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Liu, M.J.; Zhao, J.; Cai, Q.L.; Liu, G.C.; Wang, J.R.; Zhao, Z.H.; Liu, P.; Dai, L.; Yan, G.J.; Wang, W.J.; et al. The complex jujube genome provides insights into fruit tree biology. Nat. Commun. 2014, 5, 12. [Google Scholar] [CrossRef]
- Liu, M.-J.; Wang, J.-R.; Liu, P.; Zhao, J.; Zhao, Z.-H.; Dai, L.; Li, X.-S.; Liu, Z.-G. Historical Achievements and Frontier Advances in the Production and Research of Chinese Jujube (Ziziphus jujuba) in China. Acta Hortic. Sin. 2015, 42, 1683–1698. [Google Scholar]
- Lyu, R.H.; Wang, R.; Wu, C.Y.; Bao, Y.J.; Guo, P. Comparative transcriptome analysis of leaves of sour jujube seedlings under salt stress. Acta Physiol. Plant. 2022, 44, 14. [Google Scholar] [CrossRef]
- Hua, Y.; Xu, X.X.; Guo, S.; Xie, H.; Yan, H.; Ma, X.F.; Niu, Y.; Duan, J.A. Wild Jujube (Ziziphus jujuba var. spinosa): A Review of Its Phytonutrients, Health Benefits, Metabolism, and Applications. J. Agric. Food Chem. 2022, 70, 7871–7886. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Han, L.; Zhang, S.; Dai, L.; Li, B.; Han, S.; Zhao, J.; Liu, P.; Zhao, Z.; Liu, M. Insights into the evolution and spatial chromosome architecture of jujube from an updated gapless genome assembly. Plant Commun. 2023, 4, 100662. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, J.; Wang, L.; Liu, P.; Zhao, J.; Zhao, Z.; Yao, S.; Stanica, F.; Liu, Z.; Wang, L.; et al. The historical and current research progress on jujube-a superfruit for the future. Hortic. Res. 2020, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Koepke, T.; Dhingra, A. Rootstock scion somatogenetic interactions in perennial composite plants. Plant Cell Rep. 2013, 32, 1321–1337. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Maryam; Ercisli, S.; Anjum, M.A.; Ahmad, R. Progress in the Methods of Jujube Breeding. Erwerbs-Obstbau 2023, 65, 1217–1225. [Google Scholar] [CrossRef]
- Zhong, H.X.; Liu, Z.J.; Zhang, F.C.; Zhou, X.M.; Sun, X.X.; Li, Y.Y.; Liu, W.W.; Xiao, H.; Wang, N.; Lu, H.; et al. Metabolomic and transcriptomic analyses reveal the effects of self- and hetero-grafting on anthocyanin biosynthesis in grapevine. Hortic. Res. 2022, 9, 11. [Google Scholar] [CrossRef]
- Jang, Y.; Yang, E.; Cho, M.; Um, Y.; Ko, K.; Chun, C. Effect of grafting on growth and incidence of phytophthora blight and bacterial wilt of pepper (Capsicum annuum L.). Hortic. Environ. Biotechnol. 2012, 53, 9–19. [Google Scholar] [CrossRef]
- Zhou, Z.; Yuan, Y.Q.; Wang, K.T.; Wang, H.J.; Huang, J.Q.; Yu, H.; Cui, X. Rootstock-scion interactions affect fruit flavor in grafted tomato. Hortic. Plant J. 2022, 8, 499–510. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, P.; Zhu, W.Y.; Wang, F. De novo Comparative Transcriptome Analysis of Genes Differentially Expressed in the Scion of Homografted and Heterografted Tomato Seedlings. Sci. Rep. 2019, 9, 12. [Google Scholar] [CrossRef]
- Rasool, A.; Mansoor, S.; Bhat, K.M.; Hassan, G.I.; Baba, T.R.; Alyemeni, M.N.; Alsahli, A.A.; El-Serehy, H.A.; Paray, B.A.; Ahmad, P. Mechanisms Underlying Graft Union Formation and Rootstock Scion Interaction in Horticultural Plants. Front. Plant Sci. 2020, 11, 590847. [Google Scholar] [CrossRef]
- Tedesco, S.; Erban, A.; Gupta, S.; Kopka, J.; Fevereiro, P.; Kragler, F.; Pina, A.J.M. The Impact of Metabolic Scion-Rootstock Interactions in Different Grapevine Tissues and Phloem Exudates. Metabolites 2021, 11, 349. [Google Scholar] [CrossRef]
- Tandonnet, J.P.; Cookson, S.; Vivin, P.; Ollat, N. Scion genotype controls biomass allocation and root development in grafted grapevine. Aust. J. Grape Wine Res. 2010, 16, 290–300. [Google Scholar] [CrossRef]
- Zhang, F.C.; Zhong, H.X.; Zhou, X.M.; Pan, M.Q.; Xu, J.; Liu, M.B.; Wang, M.; Liu, G.T.; Xu, T.F.; Wang, Y.J.; et al. Grafting with rootstocks promotes phenolic compound accumulation in grape berry skin during development based on integrative multi-omics analysis. Hortic. Res. 2022, 9, uhac055. [Google Scholar] [CrossRef]
- Yin, W.C.; Wang, X.H.; Liu, H.; Wang, Y.; van Nocker, S.; Tu, M.X.; Fang, J.H.; Guo, J.Q.; Li, Z.; Wang, X.P. Overexpression of VqWRKY31 enhances powdery mildew resistance in grapevine by promoting salicylic acid signaling and specific metabolite synthesis. Hortic. Res. 2022, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Shi, Y.N.; Mou, Z.M.; Chen, S.Y.; Zhao, D.K. Grafting: A potential method to reveal the differential accumulation mechanism of secondary metabolites. Hortic. Res. 2022, 9, 9. [Google Scholar] [CrossRef]
- Habibi, F.; Liu, T.; Folta, K.; Sarkhosh, A. Physiological, biochemical, and molecular aspects of grafting in fruit trees. Hortic. Res. 2022, 9, 18. [Google Scholar] [CrossRef]
- Jiao, S.Z.; Zeng, F.W.; Huang, Y.P.; Zhang, L.B.; Mao, J.; Chen, B.H. Physiological, biochemical and molecular responses associated with drought tolerance in grafted grapevine. BMC Plant Biol. 2023, 23, 18. [Google Scholar] [CrossRef]
- Li, Q.S.; Gao, Y.; Wang, K.; Feng, J.R.; Sun, S.M.; Lu, X.; Liu, Z.; Zhao, D.Y.; Li, L.W.; Wang, D.J. Transcriptome Analysis of the Effects of Grafting Interstocks on Apple Rootstocks and Scions. Int. J. Mol. Sci. 2023, 24, 807. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Machin, F.; Wang, S.F.; Saplaoura, E.; Kragler, F. Heritable transgene-free genome editing in plants by grafting of wild-type shoots to transgenic donor rootstocks. Nat. Biotechnol. 2023, 41, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Thieme, C.J.; Rojas-Triana, M.; Stecyk, E.; Schudoma, C.; Zhang, W.N.; Yang, L.; Minambres, M.; Walther, D.; Schulze, W.X.; Paz-Ares, J.; et al. Endogenous Arabidopsis messenger RNAs transported to distant tissues. Nat. Plants 2015, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, X.T.; Xu, W.Y.; Liu, T.; Cai, C.M.; Chen, L.Y.; Clark, C.B.; Ma, J.X. Unidirectional movement of small RNAs from shoots to roots in interspecific heterografts. Nat. Plants 2021, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.Q.; Xiang, C.G.; Li, X.J.; Wang, T.; Lu, X.H.; Liu, Z.X.; Gao, L.H.; Zhang, W.N. Identification of Long-Distance Transmissible mRNA between Scion and Rootstock in Cucurbit Seedling Heterografts. Int. J. Mol. Sci. 2020, 21, 5253. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.Y.; Luo, H.; Wang, X.L.; Wang, X.M.; Qiu, X.J.; Liu, H.; Zhou, S.S.; Jia, K.H.; Nie, S.; Bao, Y.T.; et al. Chromosome-Scale Genome Assembly for Chinese Sour Jujube and Insights Into Its Genome Evolution and Domestication Signature. Front. Plant Sci. 2021, 12, 773090. [Google Scholar] [CrossRef] [PubMed]
- Saied, A.S.; Gebauer, J.; Hammer, K.; Buerkert, A.J.G.R.; Evolution, C. Ziziphus spina-christi (L.) Willd.: A multipurpose fruit tree. Genet. Resour. Crop Evol. 2008, 55, 929–937. [Google Scholar] [CrossRef]
- Sohail, M.; Saied, A.S.; Gebauer, J.; Buerkert, A. Effect of NaCl Salinity on Growth and Mineral Composition of Ziziphus spina-christi (L.) Willd. J. Agric. Rural Dev. Trop. Subtrop. 2009, 110, 107–114. [Google Scholar]
- Gorai, M.; Ben Romdhane, R.; Maraghni, M.; Neffati, M. Relationship between leaf gas-exchange characteristics and the performance of Ziziphus spina-christi (L.) Willd. seedlings subjected to salt stress. Photosynthetica 2019, 57, 897–903. [Google Scholar] [CrossRef]
- Wiehle, M.; Prinz, K.; Kehlenbeck, K.; Goenster, S.; Mohamed, S.A.; Buerkert, A.; Gebauer, J. The role of homegardens and forest ecosystems for domestication and conservation of Ziziphus spina-christi (L.) Willd. in the Nuba Mountains, Sudan. Genet. Resour. Crop. Evol. 2014, 61, 1491–1506. [Google Scholar] [CrossRef]
- Pawlowska, A.M.; Camangi, F.; Bader, A.; Braca, A. Flavonoids of Zizyphus jujuba L. and Zizyphus spina-christi (L.) Willd (Rhamnaceae) fruits. Food Chem. 2009, 112, 858–862. [Google Scholar] [CrossRef]
- Singh, A.; Kumar, A.; Kumar, R.; Sheoran, P.; Yadav, R.K.; Sharma, P.C. Multivariate analyses discern shared and contrasting eco-physiological responses to salinity stress of Ziziphus rootstocks and budded trees. S. Afr. J. Bot. 2022, 146, 573–584. [Google Scholar] [CrossRef]
- Zandiehvakili, G.; Khadivi, A. Identification of the promising Ziziphus spina-christi (L.) Willd.genotypes using pomological and chemical proprieties. Food Sci. Nutr. 2021, 9, 5698–5711. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, L.B.; Wu, R.L. Plant grafting: How genetic exchange promotes vascular reconnection. New Phytol. 2017, 214, 56–65. [Google Scholar] [CrossRef]
- Belmonte-Urena, L.J.; Garrido-Cardenas, J.A.; Camacho-Ferre, F. Analysis of World Research on Grafting in Horticultural Plants. Hortscience 2020, 55, 112–120. [Google Scholar] [CrossRef]
- Pal, M.D.; Mitre, I.; Asanica, A.C.; Sestras, A.F.; Peticila, A.G.; Mitre, V. The Influence of Rootstock on the Growth and Fructification of Cherry Cultivars in a High Density Cultivation System. Not. Bot. Hortic. Agrobot. Cluj-Napoca 2017, 45, 451–457. [Google Scholar] [CrossRef]
- Gautier, A.T.; Chambaud, C.; Brocard, L.; Ollat, N.; Gambetta, G.A.; Delrot, S.; Cookson, S.J. Merging genotypes: Graft union formation and scion-rootstock interactions. J. Exp. Bot. 2019, 70, 747–755. [Google Scholar] [CrossRef]
- Banerjee, A.K.; Chatterjee, M.; Yu, Y.; Suh, S.-G.; Miller, W.A.; Hannapel, D.J. Dynamics of a mobile RNA of potato involved in a long-distance signaling pathway. Plant Cell 2006, 18, 3443–3457. [Google Scholar] [CrossRef]
- Warschefsky, E.J.; Klein, L.L.; Frank, M.H.; Chitwood, D.H.; Londo, J.P.; von Wettberg, E.J.B.; Miller, A.J. Rootstocks: Diversity, Domestication, and Impacts on Shoot Phenotypes. Trends Plant Sci. 2016, 21, 418–437. [Google Scholar] [CrossRef]
- Ferlito, F.; Distefano, G.; Gentile, A.; Allegra, M.; Lakso, A.N.; Nicolosi, E. Scion-rootstock interactions influence the growth and behaviour of the grapevine root system in a heavy clay soil. Aust. J. Grape Wine Res. 2020, 26, 68–78. [Google Scholar] [CrossRef]
- Dahniya, M.; Oputa, C.; Hahn, S. Investigating source-sink relations in cassava by reciprocal grafts. Exp. Agric. 1982, 18, 399–402. [Google Scholar] [CrossRef]
- Mudge, K.; Janick, J.; Scofield, S.; Goldschmidt, E.E. A History of Grafting; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; Volume 35. [Google Scholar]
- Ahit, O.P.; Abit, S.E.; Posas, M.B. Growth and development of cassava under the traditional and the Mukibat systems of planting [in the Philippines]. Ann. Trop. Res. 1981, 21, 187–198. [Google Scholar]
- Abelenda, J.A.; Navarro, C.; Prat, S. From the model to the crop: Genes controlling tuber formation in potato. Curr. Opin. Biotechnol. 2011, 22, 287–292. [Google Scholar] [CrossRef]
- Li, G.; Ma, J.; Tan, M.; Mao, J.; An, N.; Sha, G.; Zhang, D.; Zhao, C.; Han, M. Transcriptome analysis reveals the effects of sugar metabolism and auxin and cytokinin signaling pathways on root growth and development of grafted apple. BMC Genom. 2016, 17, 150. [Google Scholar] [CrossRef]
- Liao, H.-Y.; Wang, S.; Zhou, C.-Y. Effects of Intergeneric Grafting of Schisandraceae on Root Morphology, Anatomy and Physiology of Rootstocks. Forests 2023, 14, 1183. [Google Scholar] [CrossRef]
- Sun, X.; Tian, L.; Xu, W.; Feng, L.; Jia, W.; Liu, Y.; Chen, Z.; Zhang, S.; Zhang, X.; Ru, G. Integrative Analysis of the Identified Transcriptome and Proteome Major Metabolism Pathways Involved in the Development of Grafted Apricot Hybrids. Forests 2023, 14, 417. [Google Scholar] [CrossRef]
- Amiri, M.E.; Fallahi, E. Influence of Rootstocks on Scion Growth and Leaf Mineral Content of Apple (Malus pumila ‘Golden Delicious’ and ‘Red Gala’) Trees. Acta Hortic 2012, 932, 433–440. [Google Scholar] [CrossRef]
- Clearwater, M.J.; Lowe, R.G.; Hofstee, B.J.; Barclay, C.; Mandemaker, A.J.; Blattmann, P. Hydraulic conductance and rootstock effects in grafted vines of kiwifruit. J. Exp. Bot. 2004, 55, 1371–1382. [Google Scholar] [CrossRef]
- Deng, W.W.; Han, J.Y.; Fan, Y.B.; Tai, Y.L.; Zhu, B.Y.; Lu, M.Q.; Wang, R.J.; Wan, X.C.; Zhang, Z.Z. Uncovering tea-specific secondary metabolism using transcriptomic and metabolomic analyses in grafts of Camellia sinensis and C. oleifera. Tree Genet. Genomes 2018, 14, 16. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y.; Colla, G.; Zrenner, R.; Schwarz, D. Vegetable Grafting: The Implications of a Growing Agronomic Imperative for Vegetable Fruit Quality and Nutritive Value. Front. Plant Sci. 2017, 8, 23. [Google Scholar] [CrossRef] [PubMed]
- Shitan, N.; Dalmas, F.; Dan, K.; Kato, N.; Ueda, K.; Sato, F.; Forestier, C.; Yazaki, K. Characterization of Coptis japonica CjABCB2, an ATP-binding cassette protein involved in alkaloid transport. Phytochemistry 2013, 91, 109–116. [Google Scholar] [CrossRef]
- Xu, D.B.; Yuan, H.W.; Tong, Y.F.; Zhao, L.; Qiu, L.L.; Guo, W.B.; Shen, C.J.; Liu, H.J.; Yan, D.L.; Zheng, B.S. Comparative Proteomic Analysis of the Graft Unions in Hickory (Carya cathayensis) Provides Insights into Response Mechanisms to Grafting Process. Front. Plant Sci. 2017, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Migicovsky, Z.; Harris, Z.N.; Klein, L.L.; Li, M.; McDermaid, A.; Chitwood, D.H.; Fennell, A.; Kovacs, L.G.; Kwasniewski, M.; Londo, J.P.; et al. Rootstock effects on scion phenotypes in a ‘Chambourcin’ experimental vineyard. Hortic. Res. 2019, 6, 13. [Google Scholar] [CrossRef]
- Lu, J.; Cheng, F.; Huang, Y.; Bie, Z. Grafting Watermelon Onto Pumpkin Increases Chilling Tolerance by Up Regulating Arginine Decarboxylase to Increase Putrescine Biosynthesis. Front. Plant Sci. 2021, 12, 812396. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Q.; Zhang, R.; Liu, M.; Wang, C.; Liu, Z.; Xiang, C.; Lu, X.; Zhang, X.; Li, X.; et al. Rootstock-scion exchanging mRNAs participate in the pathways of amino acids and fatty acid metabolism in cucumber under early chilling stress. Hortic. Res. 2022, 9, uhac031. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhan, R.; Wang, L.; Li, J.; Ma, B.; Ma, F.; Li, M. Overexpression of MdFRK2 enhances apple drought resistance by promoting carbohydrate metabolism and root growth under drought stress. Hortic. Plant J. 2023, 9, 884–897. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, B.; Li, N.; Chen, Z.; Xu, K. Comparative transcriptome analysis of the regulation of ABA signaling genes in different rootstock grafted tomato seedlings under drought stress. Environ. Exp. Bot. 2019, 166, 103814. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, L.; Wang, Z.; Zhang, R.; Liu, P.; Liu, M.; Liu, Z.; Zhao, Z.; Wang, L.; Chen, X.; et al. The regulation of cell wall lignification and lignin biosynthesis during pigmentation of winter jujube. Hortic. Res. 2021, 8, 238. [Google Scholar] [CrossRef]
- Liu, N.; Yang, J.; Fu, X.; Zhang, L.; Tang, K.; Guy, K.M.; Hu, Z.; Guo, S.; Xu, Y.; Zhang, M. Genome-wide identification and comparative analysis of grafting-responsive mRNA in watermelon grafted onto bottle gourd and squash rootstocks by high-throughput sequencing. Mol. Genet. Genom. MGG 2016, 291, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Asahina, M.; Satoh, S. Molecular and physiological mechanisms regulating tissue reunion in incised plant tissues. J. Plant Res. 2015, 128, 381–388. [Google Scholar] [CrossRef]
- Harada, T. Grafting and RNA transport via phloem tissue in horticultural plants. Sci. Hortic. 2010, 125, 545–550. [Google Scholar] [CrossRef]
- Cookson, S.J.; Clemente Moreno, M.J.; Hevin, C.; Nyamba Mendome, L.Z.; Delrot, S.; Trossat-Magnin, C.; Ollat, N. Graft union formation in grapevine induces transcriptional changes related to cell wall modification, wounding, hormone signalling, and secondary metabolism. J. Exp. Bot. 2013, 64, 2997–3008. [Google Scholar] [CrossRef] [PubMed]
- Melnyk, C.W.; Schuster, C.; Leyser, O.; Meyerowitz, E.M. A Developmental Framework for Graft Formation and Vascular Reconnection in Arabidopsis thaliana. Curr. Biol. CB 2015, 25, 1306–1318. [Google Scholar] [CrossRef]
- Melnyk, C.W.; Gabel, A.; Hardcastle, T.J.; Robinson, S.; Miyashima, S.; Grosse, I.; Meyerowitz, E.M. Transcriptome dynamics at Arabidopsis graft junctions reveal an intertissue recognition mechanism that activates vascular regeneration. Proc. Natl. Acad. Sci. USA. 2018, 115, E2447–E2456. [Google Scholar] [CrossRef]
- Vibhuti, M.; Kumar, A.; Sheoran, N.; Nadakkakath, A.V.; Eapen, S.J. Molecular Basis of Endophytic Bacillus megaterium-induced Growth Promotion in Arabidopsis thaliana: Revelation by Microarray-based Gene Expression Analysis. J. Plant Growth Regul. 2017, 36, 118–130. [Google Scholar] [CrossRef]
- Martinoia, E.; Klein, M.; Geisler, M.; Bovet, L.; Forestier, C.; Kolukisaoglu, U.; Muller-Rober, B.; Schulz, B. Multifunctionality of plant ABC transporters—More than just detoxifiers. Planta 2002, 214, 345–355. [Google Scholar] [CrossRef]
- Hwang, J.U.; Song, W.Y.; Hong, D.; Ko, D.; Yamaoka, Y.; Jang, S.; Yim, S.; Lee, E.; Khare, D.; Kim, K.; et al. Plant ABC Transporters Enable Many Unique Aspects of a Terrestrial Plant’s Lifestyle. Mol. Plant. 2016, 9, 338–355. [Google Scholar] [CrossRef]
- Lefevre, F.; Boutry, M. Towards Identification of the Substrates of ATP-Binding Cassette Transporters. Plant Physiol. 2018, 178, 18–39. [Google Scholar] [CrossRef]
- Barzana, G.; Rios, J.J.; Lopez-Zaplana, A.; Nicolas-Espinosa, J.; Yepes-Molina, L.; Garcia-Ibanez, P.; Carvajal, M. Interrelations of nutrient and water transporters in plants under abiotic stress. Physiol. Plant. 2021, 171, 595–619. [Google Scholar] [CrossRef]
- Olmstead, M.A.; Lang, N.S.; Ewers, F.W.; Owens, S.A. Xylem vessel anatomy of sweet cherries grafted onto dwarfing and nondwarfing rootstocks. J. Am. Soc. Hortic. Sci. 2006, 131, 577–585. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).