
A Study of the Influence of the Type of Land Use on the Enzymatic Activity of Soils in Southwestern China


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Soil Sampling
2.3. Determination of Soil Chemical Characters
2.4. Determination of Soil Enzyme Activities
2.5. Fuzzy Comprehensive Evaluation Model
2.6. Statistical Analyses
3. Results
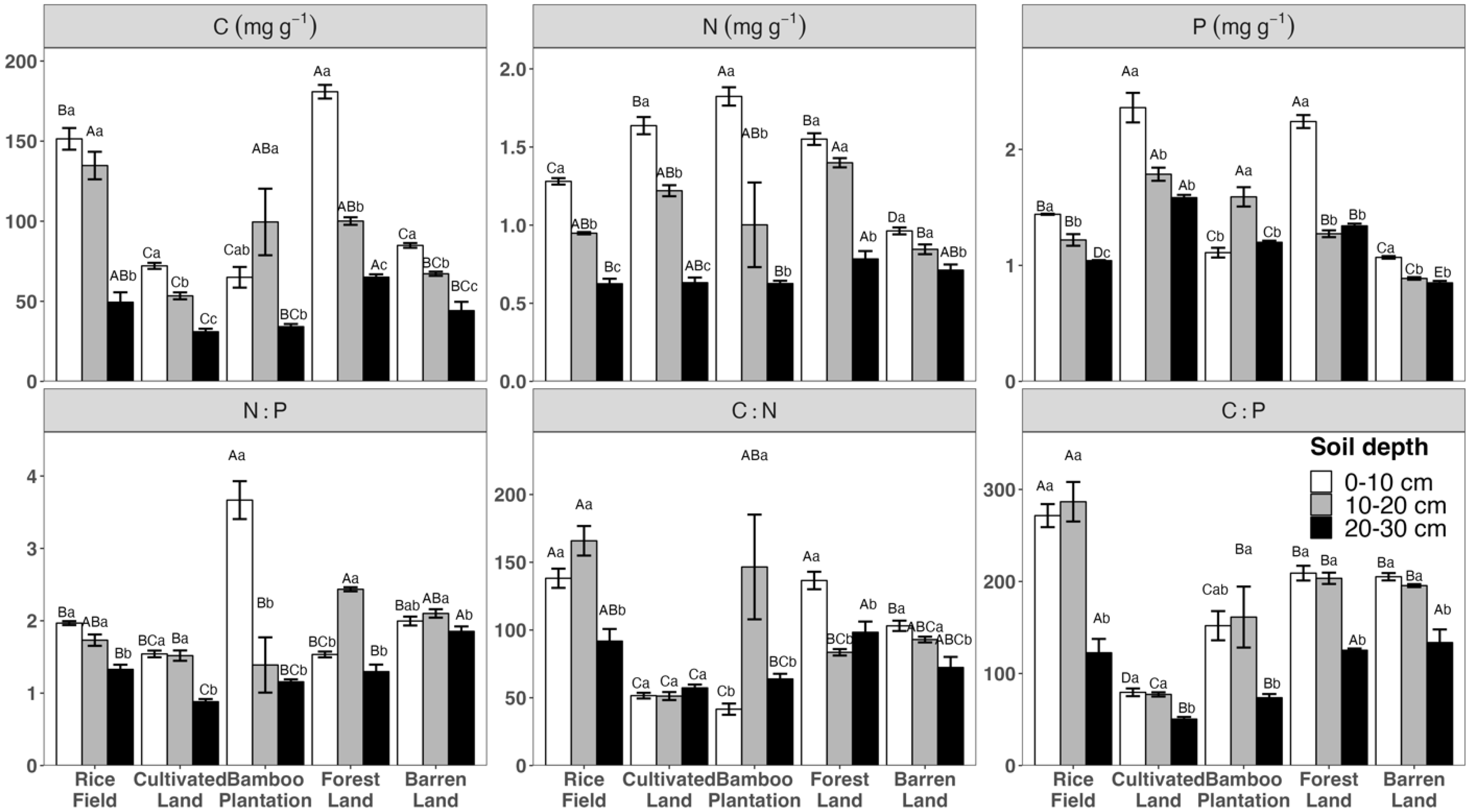
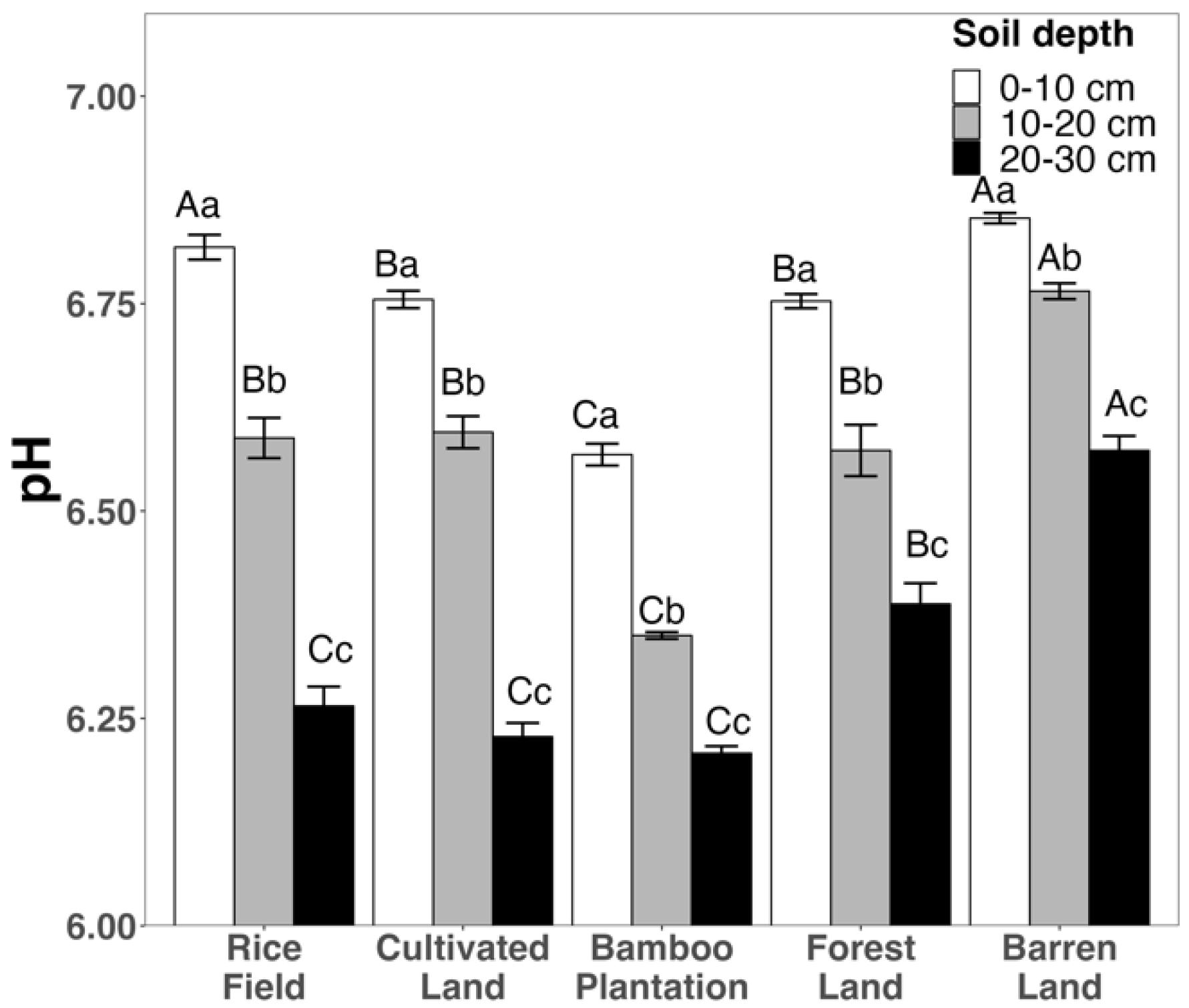
3.1. Variation in Soil Chemical Variables across Different Land Uses and Depths
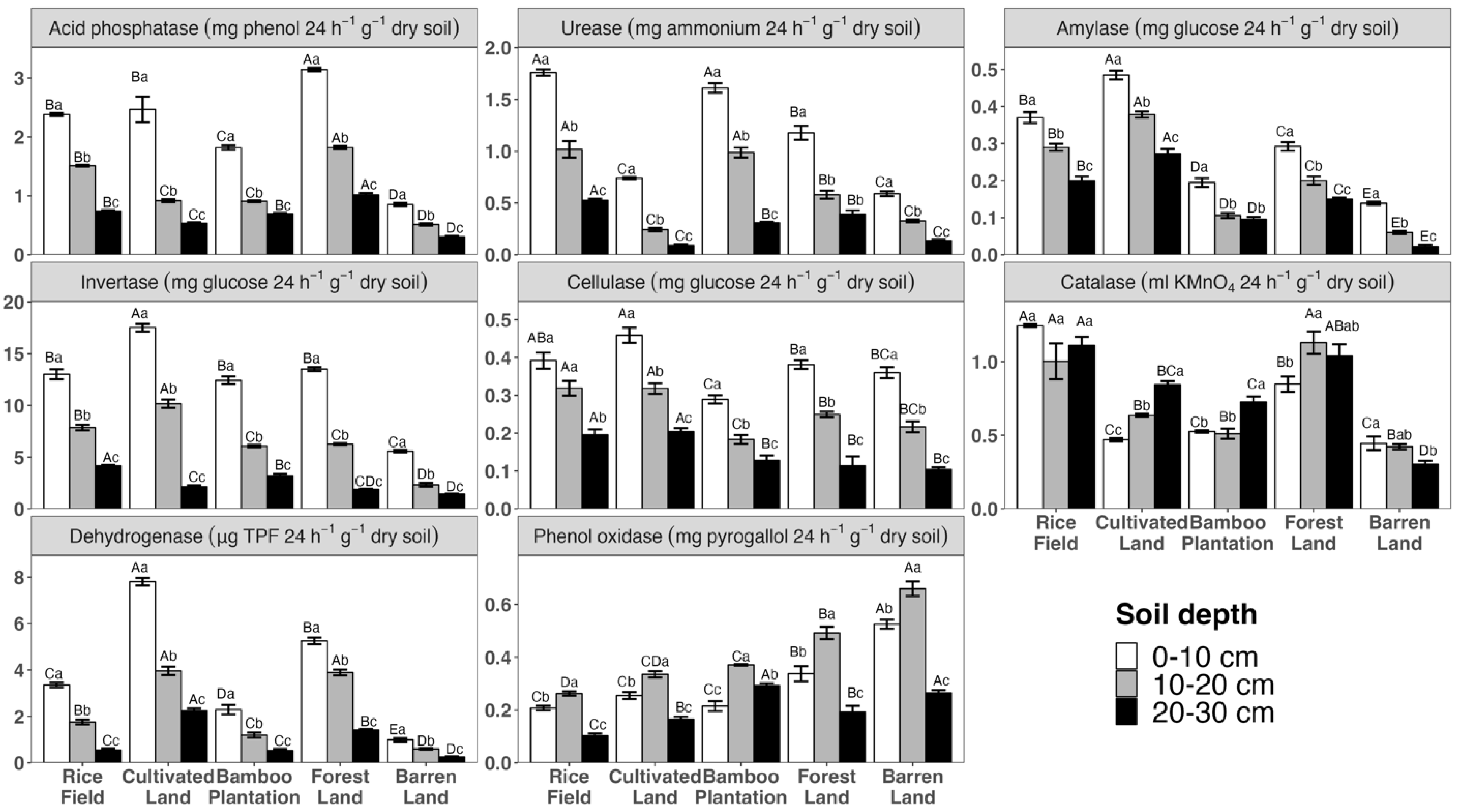
3.2. Variation in Soil Enzyme Activities across Different Land Uses and Depths
3.3. Associations between Soil Nutrients, pH, and Enzyme Activities
3.4. Fuzzy Comprehensive Evaluation
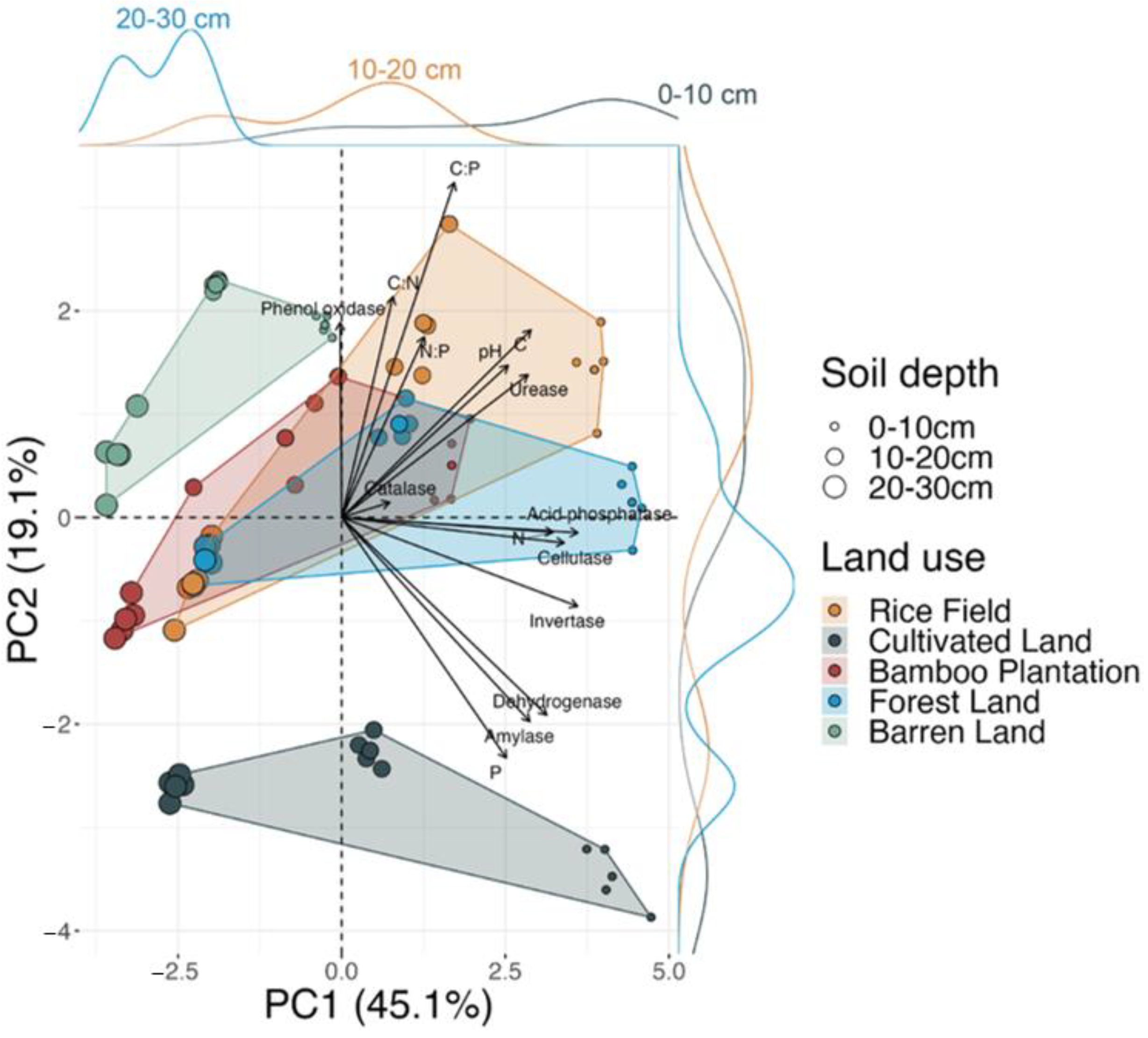
3.5. The Variability in Soil Enzyme Activities and Nutrients
4. Discussion
4.1. The Impacts of Land Use
4.2. The Impacts of Soil Depth
4.3. Fuzzy Comprehensive Evaluation Models
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dawson, J.J.C.; Smith, P. Carbon losses from soil and its consequences for land-use management. Sci. Total Environ. 2007, 382, 165–190. [Google Scholar] [CrossRef]
- Ross, D.J.; Tate, K.R.; Scott, N.A.; Feltham, C.W. Land-use change: Effects on soil carbon, nitrogen and phosphorus pools and fluxes in three adjacent ecosystems. Soil Biol. Biochem. 1999, 31, 803–813. [Google Scholar] [CrossRef]
- Wallenius, K.; Rita, H.; Mikkonen, A.; Lappi, K.; Lindström, K.; Hartikainen, H.; Raateland, A.; Niemi, R.M. Effects of land use on the level, variation and spatial structure of soil enzyme activities and bacterial communities. Soil Biol. Biochem. 2011, 43, 1464–1473. [Google Scholar] [CrossRef]
- Zhang, Y.; Bhattacharyya, R.; Dalal, R.C.; Wang, P.; Menzies, N.M.; Kopittke, P.M. Impact of land use change and soil type on total phosphorus and its fractions in soil aggregates. L. Degrad. Dev. 2020, 31, 828–841. [Google Scholar] [CrossRef]
- Miao, F.; Li, Y.; Cui, S.; Jagamma, S.; Yang, G.; Zhang, Q. Soil extracellular enzyme activities under long-term fertilization management in the croplands of China: A meta-analysis. Nutr. Cycl. Agroecosyst. 2019, 114, 125–138. [Google Scholar] [CrossRef]
- Monreal, C.M.; Bergstrom, D.W. Soil enzymatic factors expressing the influence of land use, tillage system and texture on soil biochemical quality. Can. J. Soil Sci. 2000, 80, 419–428. [Google Scholar] [CrossRef]
- Luo, Y.; Li, Q.; Shen, J.; Wang, C.; Li, B.; Yuan, S.; Zhao, B.; Li, H.; Zhao, J.; Guo, L.; et al. Effects of agricultural land use change on organic carbon and its labile fractions in the soil profile in an urban agricultural area. L. Degrad. Dev. 2019, 30, 1875–1885. [Google Scholar] [CrossRef]
- Fitts, L.A.; Domke, G.M.; Russell, M.B. Comparing methods that quantify forest disturbances in the United States’ national forest inventory. Environ. Monit. Assess. 2022, 194, 304. [Google Scholar] [CrossRef]
- Yan, H.; Liu, F.; Liu, J.; Xiao, X.; Qin, Y. Status of land use intensity in China and its impacts on land carrying capacity. J. Geogr. Sci. 2017, 27, 387–402. [Google Scholar] [CrossRef]
- Prescott, C.E.; Vesterdal, L. Decomposition and transformations along the continuum from litter to soil organic matter in forest soils. For. Ecol. Manag. 2021, 498, 119522. [Google Scholar] [CrossRef]
- Huang, Z.; Clinton, P.W.; Davis, M.R.; Yang, Y. Impacts of plantation forest management on soil organic matter quality. J. Soils Sediments 2011, 11, 1309–1316. [Google Scholar] [CrossRef]
- Lu, H.; Cai, C.; Zeng, X.; Campbell, D.E.; Fan, S.; Liu, G. Bamboo vs. crops: An integrated emergy and economic evaluation of using bamboo to replace crops in south Sichuan Province, China. J. Clean. Prod. 2018, 177, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Han, M.; Yuan, X.; Cao, G.; Zhu, B. Seasonal changes in soil properties, microbial biomass and enzyme activities across the soil profile in two alpine ecosystems. Soil Ecol. Lett. 2021, 3, 383–394. [Google Scholar] [CrossRef]
- Shirima, D.D.; Totland, Ø.; Moe, S.R. The relative importance of vertical soil nutrient heterogeneity, and mean and depth-specific soil nutrient availabilities for tree species richness in tropical forests and woodlands. Oecologia 2016, 182, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Avazpoor, Z.; Moradi, M.; Basiri, R.; Mirzaei, J.; Taghizadeh-Mehrjardi, R.; Kerry, R. Soil enzyme activity variations in riparian forests in relation to plant species and soil depth. Arab. J. Geosci. 2019, 12, 708. [Google Scholar] [CrossRef]
- Sarto, M.V.M.; Borges, W.L.B.; Bassegio, D.; Pires, C.A.B.; Rice, C.W.; Rosolem, C.A. Soil microbial community, enzyme activity, C and N stocks and soil aggregation as affected by land use and soil depth in a tropical climate region of Brazil. Arch. Microbiol. 2020, 202, 2809–2824. [Google Scholar] [CrossRef] [PubMed]
- Thoms, C.; Gattinger, A.; Jacob, M.; Thomas, F.M.; Gleixner, G. Direct and indirect effects of tree diversity drive soil microbial diversity in temperate deciduous forest. Soil Biol. Biochem. 2010, 42, 1558–1565. [Google Scholar] [CrossRef]
- Hu, A.; Yu, Z.; Liu, X.; Gao, W.; He, Y.; Li, J. The effects of irrigation and fertilization on the migration and transformation processes of main chemical components in the soil profile. Environ. Geochem. Health 2019, 41, 2631–2648. [Google Scholar] [CrossRef] [PubMed]
- Muneer, M.A.; Hou, W.; Li, J.; Huang, X.; Ur Rehman Kayani, M.; Cai, Y.; Yang, W.; Wu, L.; Ji, B.; Zheng, C. Soil pH: A key edaphic factor regulating distribution and functions of bacterial community along vertical soil profiles in red soil of pomelo orchard. BMC Microbiol. 2022, 22, 38. [Google Scholar] [CrossRef]
- Vipotnik, Z.; Michelin, M.; Tavares, T. Ligninolytic enzymes production during polycyclic aromatic hydrocarbons degradation: Effect of soil pH, soil amendments and fungal co-cultivation. Biodegradation 2021, 32, 193–215. [Google Scholar] [CrossRef]
- Samuel, A.D.; Domuta, C.; Ciobanu, C.; Sandor, M. Field management effects on soil enzyme activities. Rom. Agric. Res. 2008, 28, 61–68. [Google Scholar]
- Lehmann, J.; Schroth, G. Nutrient leaching. In Trees, Crops and Soil Fertility: Concepts and Research Methods; CABI Publishing: Wallingford, UK, 2002; pp. 151–166. [Google Scholar]
- Lv, H.; Zhao, Y.; Wang, Y.; Wan, L.; Wang, J.; Butterbach-Bahl, K.; Lin, S. Conventional flooding irrigation and over fertilization drives soil pH decrease not only in the top-but also in subsoil layers in solar greenhouse vegetable production systems. Geoderma 2020, 363, 114156. [Google Scholar] [CrossRef]
- Maxwell, T.L.; Augusto, L.; Bon, L.; Courbineau, A.; Altinalmazis-Kondylis, A.; Milin, S.; Bakker, M.R.; Jactel, H.; Fanin, N. Effect of a tree mixture and water availability on soil nutrients and extracellular enzyme activities along the soil profile in an experimental forest. Soil Biol. Biochem. 2020, 148, 107864. [Google Scholar] [CrossRef]
- Gao, Y.; Mao, L.; Miao, C.; Zhou, P.; Cao, J.; Zhi, Y.; Shi, W. Spatial characteristics of soil enzyme activities and microbial community structure under different land uses in Chongming Island, China: Geostatistical modelling and PCR-RAPD method. Sci. Total. Environ. 2010, 408, 3251–3260. [Google Scholar] [CrossRef] [PubMed]
- Dick, R.P. Soil enzyme activities as indicators of soil quality. Defin. Soil Qual. Sustain. Environ. 1994, 35, 107–124. [Google Scholar]
- Sinsabaugh, R.L. Phenol oxidase, peroxidase and organic matter dynamics of soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Jian, S.; Li, J.; Chen, J.I.; Wang, G.; Mayes, M.A.; Dzantor, K.E.; Hui, D.; Luo, Y. Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: A meta-analysis. Soil Biol. Biochem. 2016, 101, 32–43. [Google Scholar] [CrossRef]
- Yokoyama, D.; Imai, N.; Kitayama, K. Effects of nitrogen and phosphorus fertilization on the activities of four different classes of fine-root and soil phosphatases in Bornean tropical rain forests. Plant Soil 2017, 416, 463–476. [Google Scholar] [CrossRef]
- Xiao, W.; Chen, X.; Jing, X.; Zhu, B. A meta-analysis of soil extracellular enzyme activities in response to global change. Soil Biol. Biochem. 2018, 123, 21–32. [Google Scholar] [CrossRef]
- Stefanic, G.; Eliade, G.; Chirnogeanu, I. Researches concerning a biological index of soil fertility. In Proceedings of the 5th Symposium on Soil Biology, Iasi, Romania, 17 February 1981; Romanian National Society of Soil Sciences: Bucharest, Romania, 1984. [Google Scholar]
- Beck, T. Methods and application of soil microbiological analysis at the Landensanstalt fllr Bodenkultur und Pflanzenbau (LBB) in Munich for the determination of some aspects of soil fertility. In Proceedings of the 5th Symposium on Soil Biology, Iasi, Romania, 17 February 1981; Romanian National Society of Soil Sciences: Bucharest, Romania, 1984; pp. 13–20. [Google Scholar]
- Acosta-Martinez, V.; Cano, A.; Johnson, J. Simultaneous determination of multiple soil enzyme activities for soil health-biogeochemical indices. Appl. Soil Ecol. 2018, 126, 121–128. [Google Scholar] [CrossRef]
- Liu, J.; Li, S.; Ouyang, Z.; Tam, C.; Chen, X. Ecological and socioeconomic effects of China’s policies for ecosystem services. Proc. Natl. Acad. Sci. USA 2008, 105, 9477. [Google Scholar] [CrossRef]
- Kovar, J.L.; Pierzynski, G.M. Methods of Phosphorus Analysis for Soils, Sediments, Residuals, and Waters, 2nd ed.; Southern Cooperative Series Bulletin No. 408; North Carolina State University: Raleigh, NC, USA, 2009. [Google Scholar]
- Guan, S.Y. Soil Enzymes and Their Research Methodology; China Agriculture Press: Beijing, China, 1986. [Google Scholar]
- Zadeh, L.A. Fuzzy logic. Computer 1988, 21, 83–93. [Google Scholar] [CrossRef]
- Shen, G.; Lu, Y.; Wang, M.; Sun, Y. Status and fuzzy comprehensive assessment of combined heavy metal and organo-chlorine pesticide pollution in the Taihu Lake region of China. J. Environ. Manag. 2005, 76, 355–362. [Google Scholar] [CrossRef]
- Xue, Y.; Liu, S.; Hu, Y.; Yang, J. Soil quality assessment using weighted fuzzy association rules. Pedosphere 2010, 20, 334–341. [Google Scholar] [CrossRef]
- Wang, D.; Bai, J.; Wang, W.; Zhang, G.; Cui, B.; Liu, X.; Li, X. Comprehensive assessment of soil quality for different wetlands in a Chinese delta. L. Degrad. Dev. 2018, 29, 3783–3794. [Google Scholar] [CrossRef]
- Wu, C.; Liu, G.; Huang, C.; Liu, Q. Soil quality assessment in Yellow River Delta: Establishing a minimum data set and fuzzy logic model. Geoderma 2019, 334, 82–89. [Google Scholar] [CrossRef]
- Tibshirani, R. Regression shrinkage and selection via the lasso. J. R Stat. Soc. Ser. B 1996, 58, 267–288. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; García Marquéz, J.R.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Reddy, K.R.; Patrick, W.H.; Broadbent, F.E. Nitrogen transformations and loss in flooded soils and sediments. Crit. Rev. Environ. Sci. Technol. 1984, 13, 273–309. [Google Scholar] [CrossRef]
- Allison, S.D.; Vitousek, P.M. Responses of extracellular enzymes to simple and complex nutrient inputs. Soil Biol. Biochem. 2005, 37, 937–944. [Google Scholar] [CrossRef]
- Qi, D.; Wu, Q.; Zhu, J. Nitrogen and phosphorus losses from paddy fields and the yield of rice with different water and nitrogen management practices. Sci. Rep. 2020, 10, 9734. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Lam, S.K.; Wang, S.; Zhou, W.; Chen, D.; Yan, X. Optimizing nitrogen fertilization rate to enhance soil carbon storage and decrease nitrogen pollution in paddy ecosystems with simultaneous straw incorporation. Agric. Ecosyst. Environ. 2020, 298, 106968. [Google Scholar] [CrossRef]
- Haynes, R.J.; Goh, K.M. Some observations on surface soil pH, base saturation and leaching of cations under three contrasting orchard soil management practices. Plant Soil 1980, 56, 429–438. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, Y.; Li, T.; Sun, W.; Huang, Y. Net greenhouse gas balance in China’s croplands over the last three decades and its mitigation potential. Environ. Sci. Technol. 2014, 48, 2589–2597. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Jiang, P.; Xu, Z. Soil microbial functional diversity under intensively managed bamboo plantations in southern China. J. Soils Sediments 2008, 8, 177–183. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, J.; Tian, G. The potential of fertilizer management for reducing nitrous oxide emissions in the cleaner production of bamboo in China. J. Clean. Prod. 2016, 112, 2536–2544. [Google Scholar] [CrossRef]
- Mishra, S.; Singh, K.; Sahu, N.; Singh, S.N.; Manika, N.; Chaudhary, L.B.; Jain, M.K.; Kumar, V.; Behra, S.K. Understanding the relationship between soil properties and litter chemistry in three forest communities in tropical forest ecosystem. Environ. Monit. Assess. 2019, 191, 797. [Google Scholar] [CrossRef] [PubMed]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef]
- Herbien, S.A.; Neal, J.L. Soil pH and phosphatase activity. Commun. Soil Sci. Plant. Anal. 1990, 21, 439–456. [Google Scholar] [CrossRef]
- Nannipieri, P.; Trasar-Cepeda, C.; Dick, R.P. Soil enzyme activity: A brief history and biochemistry as a basis for appropriate interpretations and meta-analysis. Biol. Fertil. Soils 2018, 54, 11–19. [Google Scholar] [CrossRef]
- Neina, D. The role of soil pH in plant nutrition and soil remediation. Appl. Environ. Soil Sci. 2019, 2019, 5794869. [Google Scholar] [CrossRef]
- Hassan, W.; Chen, W.; Cai, P.; Huang, Q. Oxidative enzymes, the ultimate regulator: Implications for factors affecting their efficiency. J. Environ. Qual. 2013, 42, 1779–1790. [Google Scholar] [CrossRef]
- Stursova, M.; Sinsabaugh, R.L. Stabilization of oxidative enzymes in desert soil may limit organic matter accumulation. Soil Biol. Biochem. 2008, 40, 550–553. [Google Scholar] [CrossRef]
- Maharjan, M.; Sanaullah, M.; Razavi, B.S.; Kuzyakov, Y. Effect of land use and management practices on microbial biomass and enzyme activities in subtropical top-and sub-soils. Appl. Soil Ecol. 2017, 113, 22–28. [Google Scholar] [CrossRef]
- Sandoval-Pcbrez, A.L.; Gavito, M.E.; Garcca-Oliza, F.; Jaramillo, V.J. Carbon, nitrogen, phosphorus, and enzymatic activity under different land uses in a tropical, dry ecosystem. Soil Use Manag. 2009, 25, 419–426. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Penuelas, J. The elemental stoichiometry of aquatic and terrestrial ecosystems and its relationships with organismic lifestyle and ecosystem structure and function: A review and perspectives. Biogeochemistry 2012, 111, 1–39. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Moorhead, D.L. Resource allocation to extracellular enzyme production: A model for nitrogen and phosphorus control of litter decomposition. Soil Biol. Biochem. 1994, 26, 1305–1311. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Gong, J.; Hou, W.; Liu, J.; Malik, K.; Kong, X.; Wang, L.; Chen, X.; Tang, M.; Zhu, R.; Cheng, C.; et al. Effects of different land use types and soil depths on soil mineral elements, soil enzyme activity, and fungal community in Karst area of Southwest China. Int. J. Environ. Res. Public Health 2022, 19, 3120. [Google Scholar] [CrossRef]
- Uksa, M.; Schloter, M.; Kautz, T.; Athmann, M.; Köpke, U.; Fischer, D. Spatial variability of hydrolytic and oxidative potential enzyme activities in different subsoil compartments. Biol. Fertil. Soils 2015, 51, 517–521. [Google Scholar] [CrossRef]
- Gu, C.; Zhang, S.; Han, P.; Hu, X.; Xie, L.; Li, Y.; Brooks, M.; Liao, X.; Qin, L. Soil enzyme activity in soils subjected to flooding and the effect on nitrogen and phosphorus uptake by oilseed rape. Front. Plant Sci. 2019, 10, 368. [Google Scholar] [CrossRef]
- Drenovsky, R.E.; Steenwerth, K.L.; Jackson, L.E.; Scow, K.M. Land use and climatic factors structure regional patterns in soil microbial communities. Glob. Ecol. Biogeogr. 2010, 19, 27–39. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, R.; Frey, B.; Yang, L.; Li, M.; Ni, H. Land use change effects on diversity of soil bacterial, Acidobacterial and fungal communities in wetlands of the Sanjiang Plain, northeastern China. Sci. Rep. 2019, 9, 18535. [Google Scholar] [CrossRef]
- Patrick, W.H., Jr.; Mikkelsen, D.S.; Wells, B.R. Plant nutrient behavior in flooded soil. In Fertilizer Technology and Use, 3rd ed.; Wiley: Hoboken, NJ, USA, 1985; pp. 197–228. [Google Scholar]
- Krauss, U.; Deacon, J.W. Root turnover of groundnut (Arachis hypogaea L.) in soil tubes. Plant Soil 1994, 166, 259–270. [Google Scholar] [CrossRef]
- Basak, M.; Dutta, S.; Biswas, S.; Chakraborty, S.; Sarkar, A.; Rahaman, T.; Dey, S.; Das, M.; Biswas, P. Genomic insights into growth and development of bamboos: What have we learnt and what more to discover? Trees 2021, 35, 1771–1791. [Google Scholar] [CrossRef]
- Yang, T.; Wang, P.; Wang, W.; Jin, G.; Qiu, Y.; Shen, H.; Zhang, Z.; Zhou, Z. Early growth evaluation and biomass allocation difference between clones and families in Cupressus funebris. Eur. J. For. Res. 2023, 142, 839–850. [Google Scholar] [CrossRef]
- Pedersen, O.; Sauter, M.; Colmer, T.D.; Nakazono, M. Regulation of root adaptive anatomical and morphological traits during low soil oxygen. New Phytol. 2021, 229, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, X.; Huffman, T.; Liu, X.; Yang, J. Influence of topography and land management on soil nutrients variability in Northeast China. Nutr. Cycl. Agroecosystems 2011, 89, 427–438. [Google Scholar] [CrossRef]
- de Paul Obade, V.; Lal, R. Soil quality evaluation under different land management practices. Environ. Earth Sci. 2014, 72, 4531–4549. [Google Scholar] [CrossRef]
- Acín-Carrera, M.; José Marques, M.; Carral, P.; Álvarez, A.M.; López, C.; Martín-López, B.; González, J.A. Impacts of land-use intensity on soil organic carbon content, soil structure and water-holding capacity. Soil Use Manag. 2013, 29, 547–556. [Google Scholar] [CrossRef]
- Tischer, A.; Sehl, L.; Meyer, U.-N.; Kleinebecker, T.; Klaus, V.; Hamer, U. Land-use intensity shapes kinetics of extracellular enzymes in rhizosphere soil of agricultural grassland plant species. Plant Soil 2019, 437, 215–239. [Google Scholar] [CrossRef]
- Khormali, F.; Ajami, M.; Ayoubi, S.; Srinivasarao, C.; Wani, S.P. Role of deforestation and hillslope position on soil quality attributes of loess-derived soils in Golestan province, Iran. Agric. Ecosyst. Environ. 2009, 134, 178–189. [Google Scholar] [CrossRef]
- Wei, X.; Shao, M.; Gale, W.; Li, L. Global pattern of soil carbon losses due to the conversion of forests to agricultural land. Sci. Rep. 2014, 4, 4062. [Google Scholar] [CrossRef] [PubMed]
- Utobo, E.B.; Tewari, L. Soil enzymes as bioindicators of soil ecosystem status. Appl. Ecol. Environ. Res. 2015, 13, 147–169. [Google Scholar]
- Kautz, T.; Amelung, W.; Ewert, F.; Gaiser, T.; Horn, R.; Jahn, R.; Javanux, M.; Kemma, A.; Kuzyakov, Y.; Munch, J.; et al. Nutrient acquisition from arable subsoils in temperate climates: A review. Soil Biol. Biochem. 2013, 57, 1003–1022. [Google Scholar] [CrossRef]
- Tian, Q.; Wang, X.; Wang, D.; Wang, M.; Liao, C.; Yang, X.; Liu, F. Decoupled linkage between soil carbon and nitrogen mineralization among soil depths in a subtropical mixed forest. Soil Biol. Biochem. 2017, 109, 135–144. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, Y.; Zhang, J.; Chen, Y.; Yang, L.; Li, H.; Wang, L. Factors influencing soil enzyme activity in China’s forest ecosystems. Plant Ecol. 2018, 219, 31–44. [Google Scholar] [CrossRef]
- Zhu, Y.; Sun, L.; Fu, Q.; Guo, B.; Lin, Y.; Liu, X. Long-term rice cultivation improved coastal saline soil properties and multifunctionality of subsoil layers. Soil Use Manag. 2024, 40, e12918. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Land Use | Soil Depth | Land Use × Soil Depth | ||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | p | Df | F | p | df | F | p | |
| Acid phosphatase | 4 | 226.77 | *** | 2 | 765.45 | *** | 8 | 32.35 | *** |
| Urease | 4 | 256.34 | *** | 2 | 720.99 | *** | 8 | 27.03 | *** |
| Amylase | 4 | 493.47 | *** | 2 | 311.22 | *** | 8 | 6.78 | *** |
| Invertase | 4 | 310.59 | *** | 2 | 1989.9 | *** | 8 | 72.28 | *** |
| Cellulase | 4 | 36.49 | *** | 2 | 283.52 | *** | 8 | 2.9 | 0.011 |
| Catalase | 4 | 104.57 | *** | 2 | 4.74 | 0.014 | 8 | 7.45 | *** |
| Dehydrogenase | 4 | 641.71 | *** | 2 | 843.69 | *** | 8 | 76.24 | *** |
| Phenol oxidase | 4 | 133.28 | *** | 2 | 222.51 | *** | 8 | 19.79 | *** |
| C | 4 | 54.39 | *** | 2 | 122.01 | *** | 8 | 14.22 | *** |
| N | 4 | 13.89 | *** | 2 | 123.31 | *** | 8 | 6.83 | *** |
| P | 4 | 173.75 | *** | 2 | 103.9 | *** | 8 | 33.14 | *** |
| C:N | 4 | 19.02 | *** | 2 | 9.33 | * | 8 | 8 | *** |
| N:P | 4 | 15.35 | *** | 2 | 52.11 | *** | 8 | 23.88 | *** |
| C:P | 4 | 62.93 | *** | 2 | 67.21 | *** | 8 | 4.41 | * |
| pH | 4 | 161.53 | *** | 2 | 736.91 | *** | 8 | 14.83 | *** |
| Acid Phosphatase | Urease | Amylase | Invertase | Cellulase | Catalase | Dehydrogenase | Phenol Oxidase | |
|---|---|---|---|---|---|---|---|---|
| R2 | 0.74 | 0.55 | 0.52 | 0.67 | 0.6 | 0.37 | 0.79 | 0.26 |
| C | 0.44 | 0.91 | 0.04 | 0.25 | 0.08 | 0.49 | ||
| N | 0.39 | 0.13 | 0.28 | 0.66 | 0.17 | 0.67 | ||
| P | 0.78 | 1.62 | 1.02 | 0.55 | 0.15 | 2.05 | ||
| C:N | 0.03 | |||||||
| N:P | ||||||||
| C:P | ||||||||
| pH | 3.18 | 6.62 | −8.24 | 2.29 | 5.27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Xu, W.; Chen, Y.; Liu, M.; Wen, J. A Study of the Influence of the Type of Land Use on the Enzymatic Activity of Soils in Southwestern China. Forests 2024, 15, 581. https://doi.org/10.3390/f15040581
Ma Z, Xu W, Chen Y, Liu M, Wen J. A Study of the Influence of the Type of Land Use on the Enzymatic Activity of Soils in Southwestern China. Forests. 2024; 15(4):581. https://doi.org/10.3390/f15040581
Chicago/Turabian StyleMa, Zhiliang, Wenjuan Xu, Yamei Chen, Mei Liu, and Jiahao Wen. 2024. "A Study of the Influence of the Type of Land Use on the Enzymatic Activity of Soils in Southwestern China" Forests 15, no. 4: 581. https://doi.org/10.3390/f15040581
APA StyleMa, Z., Xu, W., Chen, Y., Liu, M., & Wen, J. (2024). A Study of the Influence of the Type of Land Use on the Enzymatic Activity of Soils in Southwestern China. Forests, 15(4), 581. https://doi.org/10.3390/f15040581