

Physiological Activity of Quercus suber with a High Presence of Cerambyx welensii

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Site and Trees Studied
2.2. Physiological and Morphological Measurements
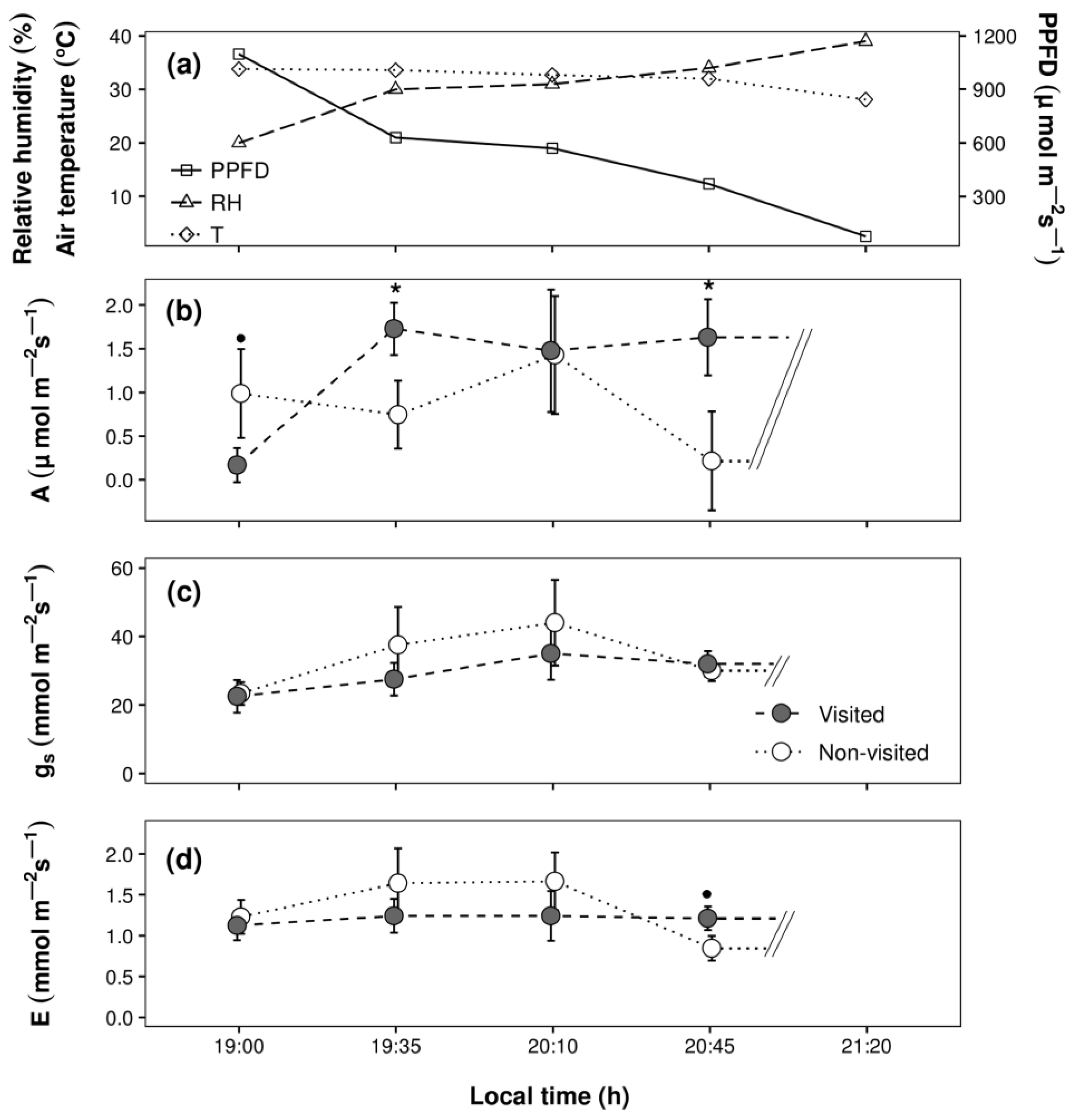
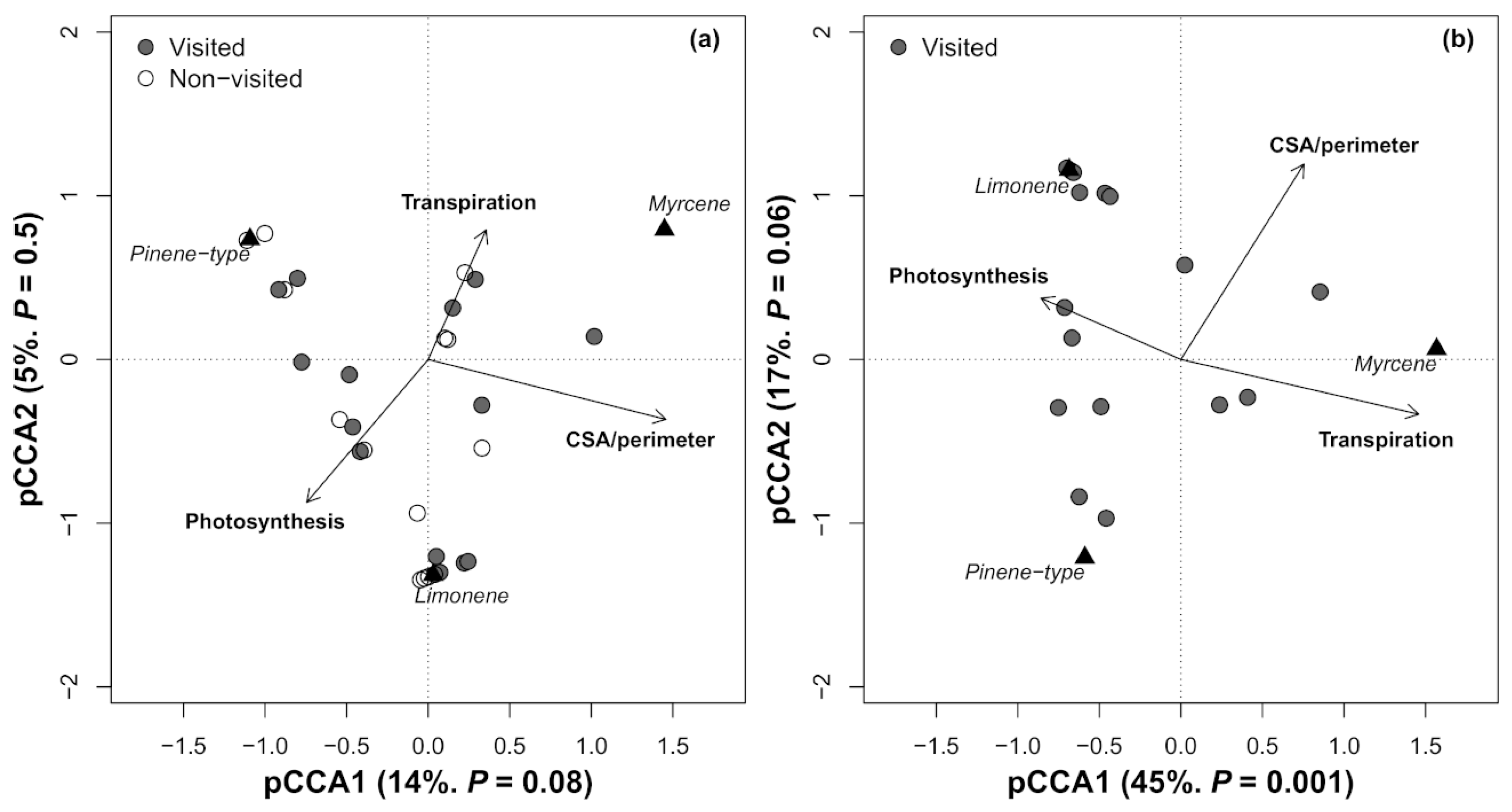
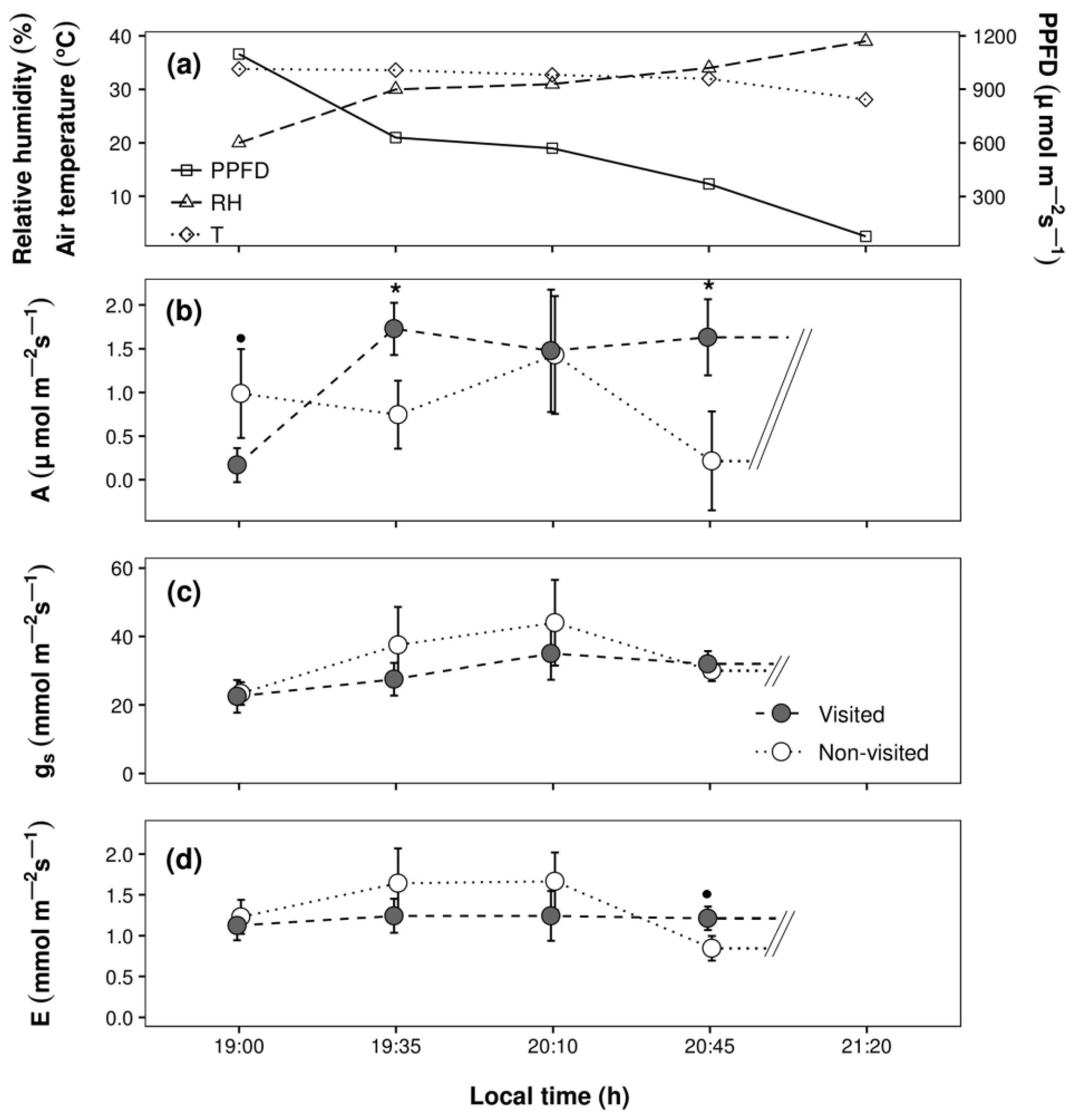
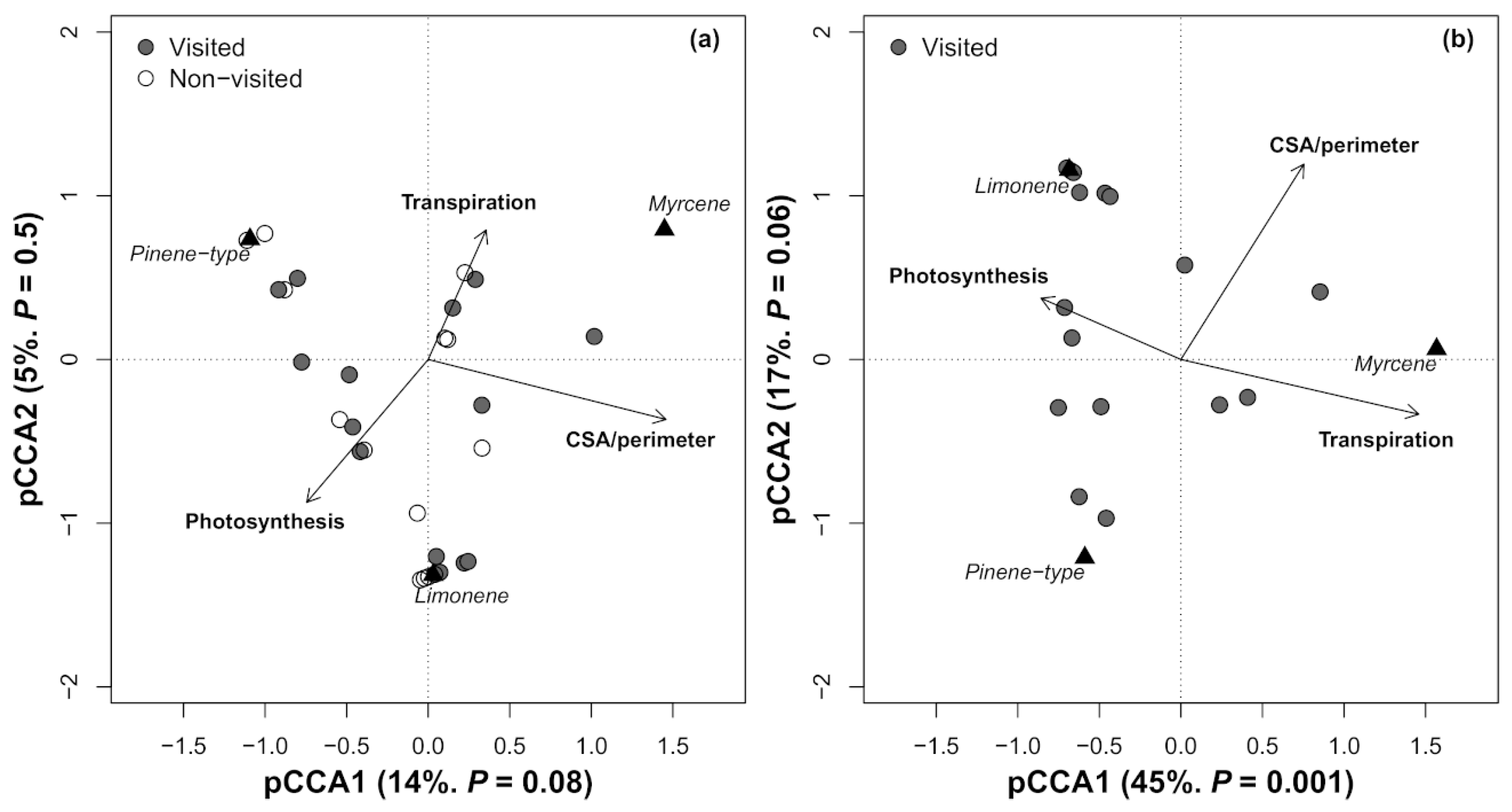
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Wide Fund for Nature (WWF)/Adena. La Dehesa en los Programas de Desarrollo Rural 2007-13. Propuesta; World Wide Fund for Nature (WWF)/Adena: Madrid, Spain, 2006; p. 34. [Google Scholar]
- Aronson, J.; Pereira, J.S.; Pausas, J.G. (Eds.) Cork Oak Woodlands on the Edge. Ecology, Adaptive Management, and Restoration; Society for Ecological Restoration International, Island Press: Washington, DC, USA, 2009. [Google Scholar]
- López-Pantoja, G.; Domínguez, L.; Sánchez-Osorio, I. Mark-recapture estimates of the survival and recapture rates of Cerambyx welensii Küster (Coleoptera Cerambycidae) in a cork oak dehesa in Huelva (Spain). Cent. Eur. J. Biol. 2008, 3, 431–441. [Google Scholar] [CrossRef]
- Torres-Vila, L.M.; Mendiola-Díaz, F.J.; Sánchez-González, Á. Dispersal differences of a pest and a protected Cerambyx species (Coleoptera: Cerambycidae) in oak open woodlands: A mark-recapture comparative study. Ecol. Entomol. 2017, 42, 18–32. [Google Scholar] [CrossRef]
- Vives, E. Coleoptera, Cerambycidae. In Fauna Ibérica; Ramos, M.A., Ed.; Museo Nacional de Ciencias Naturales, CSIC: Madrid, Spain, 2000; Volume 12. [Google Scholar]
- Martín, J.; Cabezas, J.; Buyolo, T.; Patón, D. The relationship between Cerambyx spp. damage and subsequent Biscogniauxia mediterranum infection on Quercus suber forests. For. Ecol. Manag. 2005, 216, 166–174. [Google Scholar] [CrossRef]
- Domínguez, L.; López-Pantoja, G.; Cremades, D.; Paramio, A.; Hidalgo, P.J.; Sánchez-Osorio, I. Incidence of Large Wood Borers in the Conservation of dehesa Islands Forests in Southwestern Spain. Forests 2022, 13, 413. [Google Scholar] [CrossRef]
- Correia, B.; Rodriguez, J.L.; Valledor, L.; Almeida, T.; Santos, C.; Cañal, M.J.; Pinto, G. Analysis of the expression of putative heat-stress related genes in relation to thermotolerance of cork oak. J. Plant. Physiol. 2014, 171, 399–406. [Google Scholar] [CrossRef]
- Piayda, A.; Dubbert, M.; Rebmann, C.; Kolle, O.; Costa, F.; Correia, A.; Pereira, J.S.; Werner, C.; Cuntz, M. Drought impact on carbon and water cycling in a Mediterranean Quercus suber L. woodland during the extreme drought event in 2012. Biogeosciences 2014, 11, 7159–7178. [Google Scholar] [CrossRef]
- Llusià, J.; Roahtyn, S.; Yakir, D.; Rotenberg, E.; Seco, R.; Guenther, A.; Peñuelas, J. Photosynthesis, stomatal conductance and terpene emission response to water availability in dry and mesic Mediterranean forests. Trees 2015, 30, 749–759. [Google Scholar] [CrossRef]
- Lavoir, A.V.; Duffet, C.; Mouillot, F.; Rambal, S.; Ratte, J.P.; Schnitzler, J.P.; Staudt, M. Scaling-up leaf monoterpene emissions from a water limited Quercus ilex woodland. Atmos. Environ. 2011, 45, 2888–2897. [Google Scholar] [CrossRef]
- Grote, R.; Monson, R.; Niinemets, Ü. Leaf-level models of constitutive and stress-driven volatile organic compound emissions. In Biology, Controls and Models of Tree Volatile Organic Compound Emission; Niinemets, Ü., Monson, R.K., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 315–355. [Google Scholar]
- Loreto, F.; Pollastri, S.; Fineschi, S.; Velikovac, V. Volatile isoprenoids and their importance for protection against environmental constraints in the Mediterranean area. Environ. Exp. Bot. 2014, 103, 99–106. [Google Scholar] [CrossRef]
- Hakola, H.; Laurila, T.; Lindfors, V.; Hellén, H.; Gaman, A.; Rinne, J. Variation of the VOC emission rates of birch species during the growing season. Boreal. Environ. Res. 2001, 6, 237–249. [Google Scholar]
- Loreto, F.; Ciccioli, P.; Cecinato, A.; Bracaleoni, E.; Frattoni, M.; Tricoli, D. Influence of environmental α-pinene from factors and air composition on the emission of Quercus ilex leaves. Plant. Physiol. 1996, 110, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Llusia, J. Seasonal emission of monoterpenes by the Mediterranean tree Quercus ilex in field conditions: Relations with photosynthetic rates, temperature and volatility. Physiol. Plant. 1999, 105, 641–647. [Google Scholar] [CrossRef]
- Pearse, I.S.; Gee, W.S.; Beck, J.J. Headspace Volatiles from 52 oak species advertise induction, species identity, and evolution, but not Defense. J. Chem. Ecol. 2013, 39, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Pio, C.A.; Silva, P.A.; Cerqueira, M.A.; Nunes, T.V. Diurnal and seasonal emissions of volatile organic compounds from cork oak (Quercus suber) trees. Atmos. Environ. 2005, 39, 1817–1827. [Google Scholar] [CrossRef]
- Staudt, M.; Mir, C.; Joffre, R.; Rambal, S.; Bonin, A.; Landais, D.; Lumaret, R. Isoprenoid emission of Quercus spp. (Q. suber and Q. ilex) in mixed stands contrasting in interspecific genetic introgression. New Phytol. 2004, 163, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Allison, J.D.; Borden, J.H.; Seybold, J.H. A review of the chemical ecology of the Cerambycidae (Coleoptera). Chemoecology 2004, 14, 123–150. [Google Scholar] [CrossRef]
- Sánchez-Osorio, I.; López-Pantoja, L.; Tapias, R.; Pareja-Sánchez, E.; Domínguez, L. Monoterpene emission of Quercus suber L. highly infested by Cerambyx welensii Küster. Ann. For. Sci. 2019, 76, 98. [Google Scholar] [CrossRef]
- Bense, U. Longhorn Beetles: Illustrated Key to the Cerambycidae and Vesperidae of Europe; Margraf Verlag: Weikersheim, Germany, 1995. [Google Scholar]
- Google. Satellite Image of Dehesa San Enrique (37°15’43.73” N, 6°28’34.65” O). Almonte, Huelva, Spain. October 2015. Available online: http://earth.google.com (accessed on 3 December 2023).
- Zhang, Q.-H.; Birgersson, G.; Zhu, J.; Lofstedt, C.; Lofqvist, J.; Schlyter, F. Leaf volatiles from nonhost deciduous trees: Variation by tree species, season and temperature, and electrophysiological activity in Ips typographus. J. Chem. Ecol. 1999, 8, 1923–1943. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Lme4: Linear Mixed-Effects Models Using Eigen and S4. R Package, version 1.1-7; 2014; Available online: http://github.com/lme4/lme4/ (accessed on 1 December 2023).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. 1995, B57, 289–300. [Google Scholar] [CrossRef]
- Lenth, R.V. Lsmeans: Least-Squares Means. R Package; Version 2.10; 2014; Available online: http://CRAN.R-project.org/package=lsmeans (accessed on 3 June 2016).
- Oksanen, F.; Simpson, G.L.; Blanchet, G.F.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package. R Package; Version 2.2-1; 2015; Available online: http://CRAN.R-project.org/package=vegan (accessed on 5 June 2016).
- Munné-Bosch, S.; Peñuelas, J.; Asensio, D.; Llusià, J. Airborne ethylene may alter antioxidant protection and reduce tolerance of holm oak to heat and drought stress. Plant Physiol. 2004, 136, 2937–2947. [Google Scholar] [CrossRef]
- Vaz, M.; Pereira, J.S.; Gazarini, L.C.; David, T.S.; David, J.S.; Rodrigues, A.; Maroco, J.; Chaves, M.M. Drought-induced photosynthetic inhibition and autumn recovery in two Mediterranean oak species (Quercus ilex and Quercus suber). Tree Physiol. 2010, 30, 946–956. [Google Scholar] [CrossRef]
- Vaz, M.; Maroco, J.; Ribeiro, N.; Gazarini, L.C.; Pereira, J.S.; Chaves, M.M. Leaf-level responses to light in two co-occurring Quercus (Quercus ilex and Quercus suber): Leaf structure, chemical composition and photosynthesis. Agroforest. Syst. 2011, 82, 173–181. [Google Scholar] [CrossRef]
- Peñuelas, J.; Llusià, J. Linking photorespiration, monoterpenes and thermotolerance in Quercus. New Phytol. 2002, 155, 227–237. [Google Scholar] [CrossRef]
- Llusià., J.; Peñuelas, J.; Asensio, D.; Munné-Bosch, S. Airborne limonene confers limited thermotolerance to Quercus ilex. Physiol. Plant. 2005, 123, 40–48. [Google Scholar] [CrossRef]
- Tenhunen, J.D.; Reynolds, J.F.; Lange, O.L.; Dougherty, R.L.; Harley, P.C.; Kummerow, J.; Rambal, S. Quinta. A physiologically-based growth simulator for drought adapted woody plant species. In Biomass Production by Fast-Growing Trees; Pereira, J.S., Landsberg, J.J., Eds.; NATO ASI Series; Springer: Dordrecht, The Netherlands, 1989; Volume 166, pp. 135–168. [Google Scholar]
- Oliveira, G.; Werner, C.; Mertens, C.; Correia, O. Influencia de la Posición de la Copa Sobre la Fenología y las Relaciones Hídricas en Alcornoque (Quercus suber); Actas del I. Congreso Forestal Español: Lourizán, Spain, 1993; pp. 277–282. [Google Scholar]
- Niinemets, U.; Seufert, G.; Steinbrecher, R.; Tenhunen, J.D. A model coupling foliar monoterpene emissions to leaf photosynthetic characteristics in Mediterranean evergreen Quercus species. New Phytol. 2002, 153, 257–275. [Google Scholar] [CrossRef]
- Delfine, S.; Csiky, O.; Seufert, G.; Loreto, F. Fumigation with exogenous monoterpenes of a non-isoprenoid-emitting oak (Quercus suber): Monoterpene acquisition, translocation, and effect on the photosynthetic properties at high temperatures. New Phytol. 2000, 146, 27–36. [Google Scholar] [CrossRef]
- Peñuelas, J.; Llusià, J.; Asensio, D.; Munné-Bosch, S. Linking isoprene with plant thermotolerance, antioxidants and monoterpene emissions. Plant. Cell Environ. 2005, 28, 278–286. [Google Scholar] [CrossRef]
- Núñez, L.; Plaza, J.; Pérez-Pastor, R.; Pujadas, M.; Gimeno, B.; Bermejo, V.; García-Alonso, S. High water vapour pressure deficit influence on Quercus ilex and Pinus pinea field monoterpene emission in the central Iberian Peninsula (Spain). Atmos. Environ. 2002, 36, 4441–4452. [Google Scholar] [CrossRef]
- Millar, J.G.; Hanks, L.M. Chemical ecology of cerambycid beetles. In Cerambycidae of the World: Biology and Management; Wang, Q., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2017. [Google Scholar]
- Torres-Vila, L.M.; Sanchez-González, A.; Ponce-Escudero, F.; Martín-Vertedor, D.; Ferrero-García, J.J. Assessing mass trapping efficiency and population density of Cerambyx welensii Küster by mark-recapture in dehesa open woodlands. Eur. J. Forest. Res. 2012, 131, 1103–1116. [Google Scholar] [CrossRef]
- Sánchez-Osorio, I.; Tapias, R.; Domínguez, L.; López-Pantoja, G.; González, M.d.M. Electroantennographic Responses of Cerambyx welensii Küster to Host-Related Volatiles. Forests 2021, 12, 1168. [Google Scholar] [CrossRef]
- Sánchez-Osorio, I.; López-Pantoja, L.; Paramio, A.M.; Lencina, J.L.; Gallego, D.; Domínguez, L. Field attraction of Cerambyx welensii to fermentation odors and host monoterpenes. J. Pest. Sci. 2016, 89, 59–68. [Google Scholar] [CrossRef]
- Grote, R. Sensitivity of volatile monoterpene emission to changes in canopy structure: A model-based exercise with a process-based emission model. New Phytol. 2007, 173, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Hoeber, S.; Leuschner, C.; Köhler, L.; Arias-Aguilar, D.; Schuldt, B. The importance of hydraulic conductivity and wood density to growth performance in eight tree species from a tropical semi-dry climate. For. Ecol. Manag. 2014, 330, 126–136. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Osorio, I.; López-Pantoja, G.; Tapias, R.; Pareja-Sánchez, E.; Domínguez, L. Physiological Activity of Quercus suber with a High Presence of Cerambyx welensii. Forests 2024, 15, 282. https://doi.org/10.3390/f15020282
Sánchez-Osorio I, López-Pantoja G, Tapias R, Pareja-Sánchez E, Domínguez L. Physiological Activity of Quercus suber with a High Presence of Cerambyx welensii. Forests. 2024; 15(2):282. https://doi.org/10.3390/f15020282
Chicago/Turabian StyleSánchez-Osorio, Israel, Gloria López-Pantoja, Raúl Tapias, Evangelina Pareja-Sánchez, and Luis Domínguez. 2024. "Physiological Activity of Quercus suber with a High Presence of Cerambyx welensii" Forests 15, no. 2: 282. https://doi.org/10.3390/f15020282
APA StyleSánchez-Osorio, I., López-Pantoja, G., Tapias, R., Pareja-Sánchez, E., & Domínguez, L. (2024). Physiological Activity of Quercus suber with a High Presence of Cerambyx welensii. Forests, 15(2), 282. https://doi.org/10.3390/f15020282