
Functional Analysis of the HbREF1 Promoter from Hevea brasiliensis and Its Response to Phytohormones


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
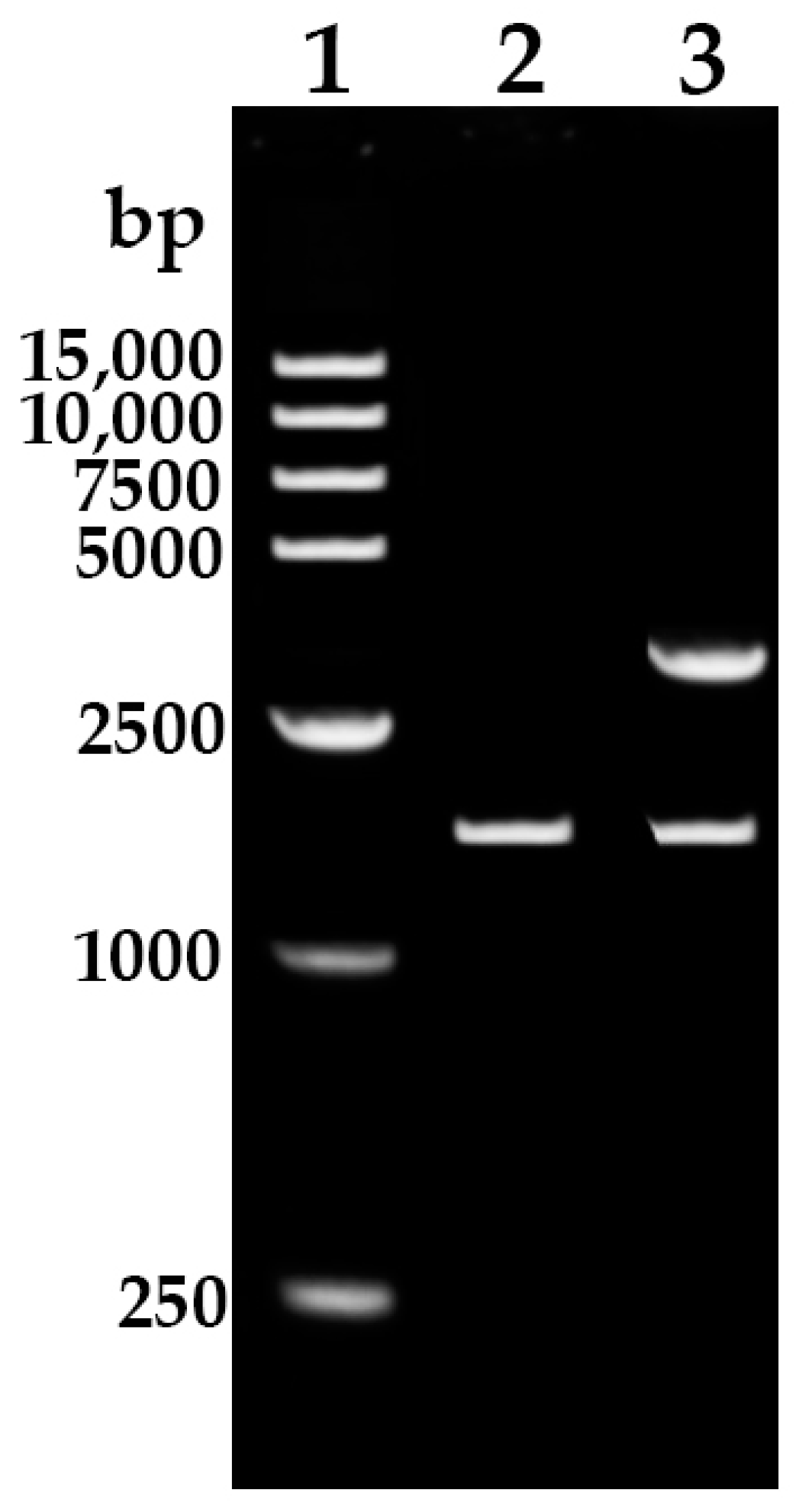
2.2. Cloning and Sequence Analysis of the HbREF1 Promoter
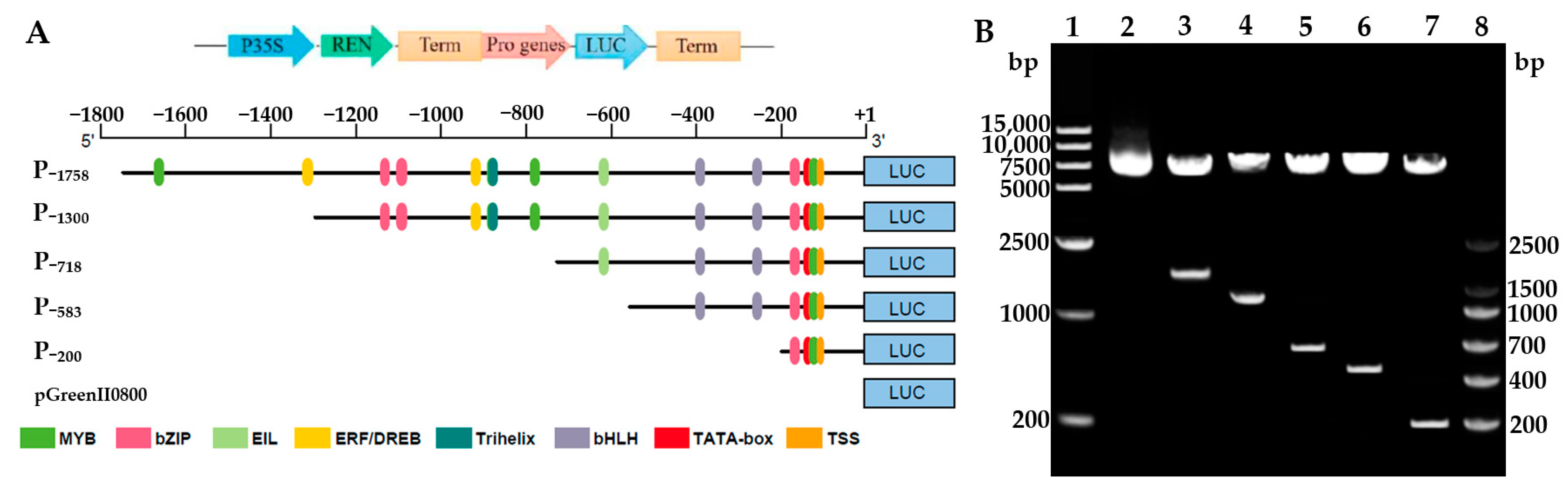
2.3. Construction of the HbREF1 Promoter Deletion Constructs
2.4. Agrobacterium-Mediated Transient Expression Assays
2.5. Plant Hormone Treatments
2.6. Imaging of Luciferase Bioluminescence
2.7. Dual-luciferase Activity Assays
3. Results
3.1. Cloning of HbREF1 Promoter
3.2. HbREF1 Promoter Sequence Analysis
3.3. Construction and Identification of the 5′ Deletion Fragments of HbREF1 Promoter
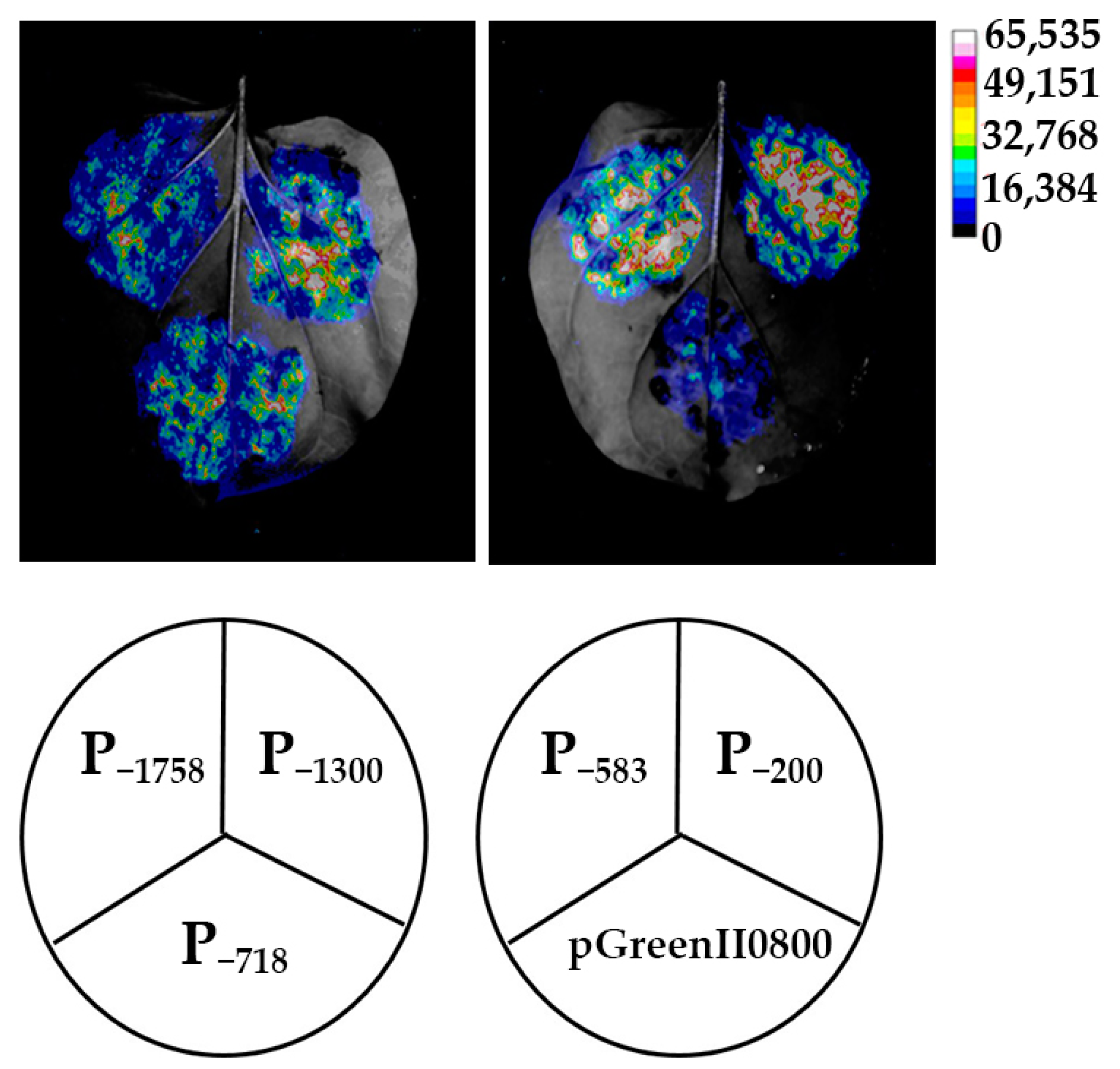
3.4. Expression Analysis of Luciferase Driven by HbREF1 Promoter
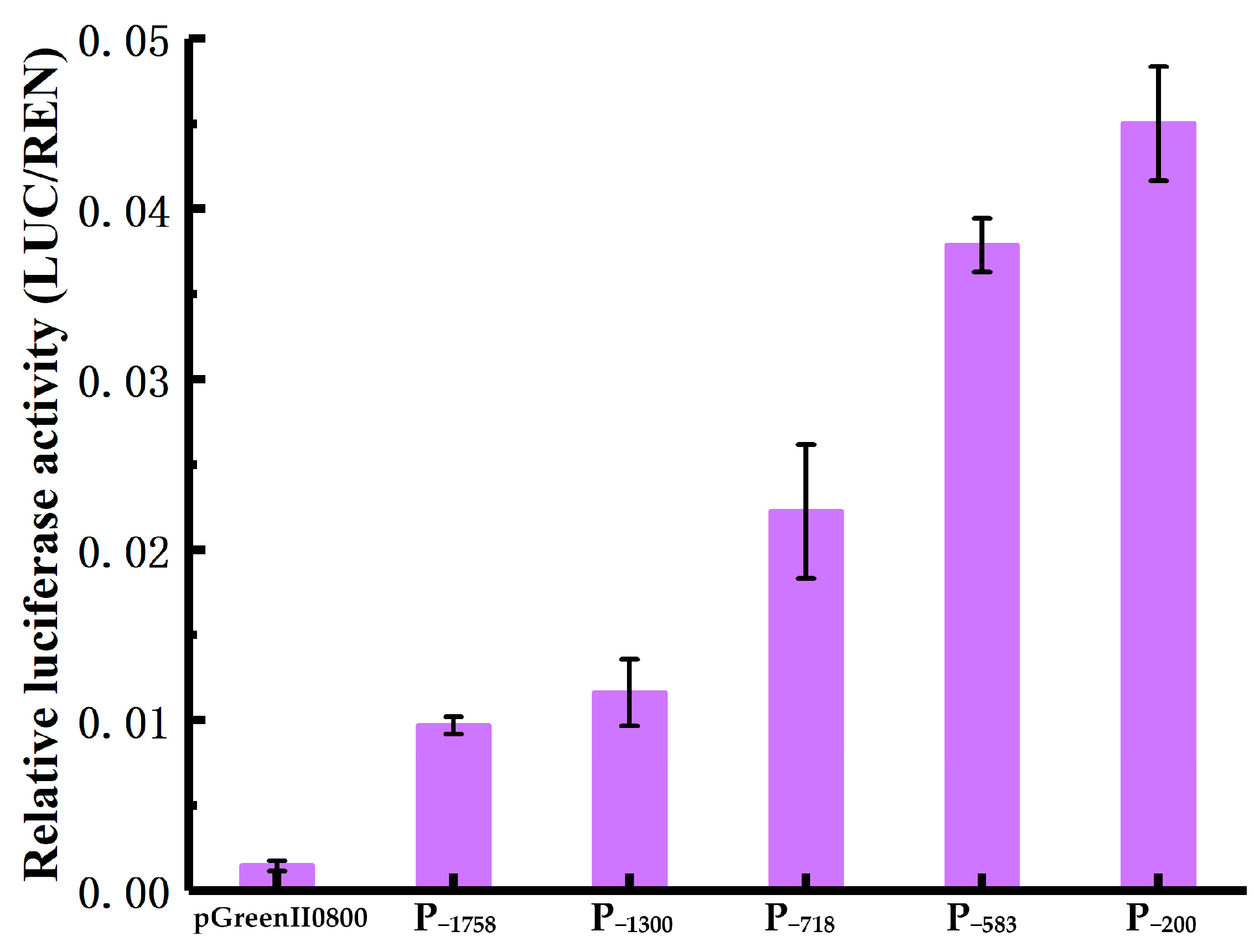
3.5. Basal Transcriptional Activity Analysis of HbREF1 Promoter
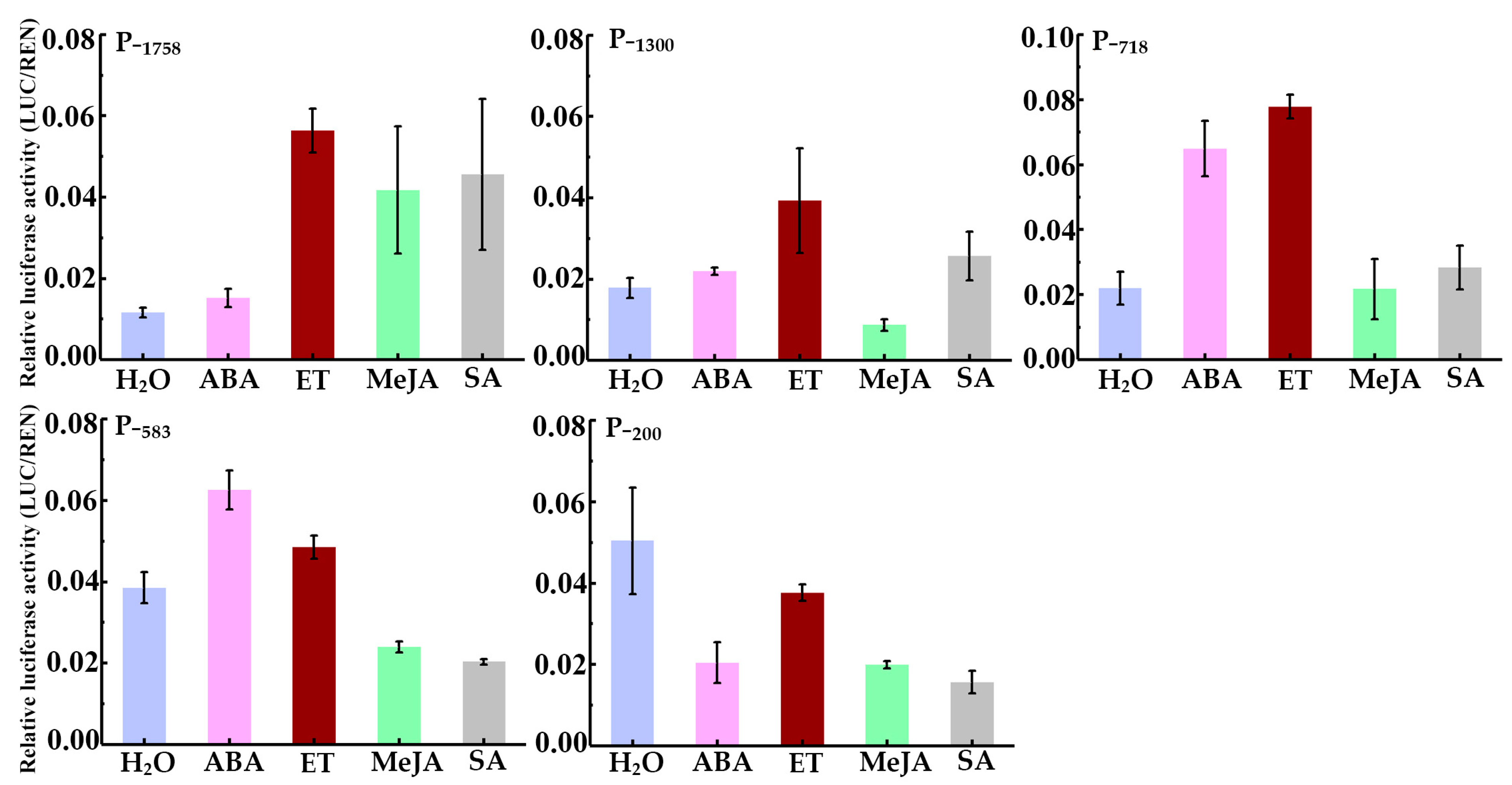
3.6. Responsiveness Analysis of HbREF1 Promoter to Phytohormone Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gireesh, T.; Meenakumari, T.; Mydin, K.K. Fast track evaluation and selection of Hevea brasiliensis clones from a clonal nursery. Ind. Crop Prod. 2017, 103, 195–201. [Google Scholar] [CrossRef]
- Silva, M.J.; Cunha Claro, P.I.; Da Silva, J.C.; Scaloppi Junior, E.J.; Goncalves, P.D.S.; Martins, M.A.; Capparelli Mattoso, L.H. Evaluation of the physicochemical properties of natural rubber from Hevea brasiliensis clones. Ind. Crop Prod. 2021, 171, 113925. [Google Scholar] [CrossRef]
- Men, X.; Wang, F.; Chen, G.Q.; Zhang, H.B.; Xian, M. Biosynthesis of natural rubber: Current state and perspectives. Int. J. Mol. Sci. 2019, 20, 50. [Google Scholar] [CrossRef]
- Wei, X.; Peng, P.; Peng, F.; Dong, J. Natural polymer Eucommia ulmoides rubber: A novel material. J. Agric. Food Chem. 2021, 69, 3797–3821. [Google Scholar] [CrossRef]
- Dennis, M.S.; Light, D.R. Rubber elongation factor from Hevea brasiliensis. Identification, characterization, and role in rubber biosynthesis. J. Biol. Chem. 1989, 264, 18608–18617. [Google Scholar] [CrossRef]
- Bahri, A.R.S.; Hamzah, S. Immunocytochemical localisation of rubber membrane protein in Hevea latex. J. Nat. Rubber Res. 1996, 11, 88–95. [Google Scholar]
- Yeang, H.Y.; Cheong, K.F.; Sunderasan, E.; Hamzah, S.; Chew, N.P.; Hamid, S.; Hamilton, R.G.; Cardosa, M.J. The 14.6 kd rubber elongation factor (Hev b 1) and 24 kd (Hev b 3) rubber particle proteins are recognized by IgE from patients with spina bifida and latex allergy. J. Allergy Clin. Immunol. 1996, 98, 628–639. [Google Scholar] [CrossRef]
- Oh, S.K.; Kang, H.; Shin, D.H.; Yang, J.; Chow, K.S.; Yeang, H.Y.; Wagner, B.; Breiteneder, H.; Han, K.H. Isolation, characterization, and functional analysis of a novel cDNA clone encoding a small rubber particle protein from Hevea brasiliensis. J. Biol. Chem. 1999, 274, 17132–17138. [Google Scholar] [CrossRef]
- Kim, I.J.; Ryu, S.B.; Kwak, Y.S.; Kang, H. A novel cDNA from Parthenium argentatum Gray enhances the rubber biosynthetic activity in vitro. J. Exp. Bot. 2004, 55, 377–385. [Google Scholar] [CrossRef]
- Laibach, N.; Hillebrand, A.; Twyman, R.M.; Prüfer, D.; Schulze Gronover, C. Identification of a Taraxacum brevicorniculatum rubber elongation factor protein that is localized on rubber particles and promotes rubber biosynthesis. Plant J. 2015, 82, 609–620. [Google Scholar] [CrossRef]
- Priya, P.; Venkatachalam, P.; Thulaseedharan, A. Differential expression pattern of rubber elongation factor (REF) mRNA transcripts from high and low yielding clones of rubber tree (Hevea brasiliensis Muell. Arg.). Plant Cell Rep. 2007, 26, 1833–1838. [Google Scholar] [CrossRef]
- Priya, P.; Venkatachalam, P.; Thulaseedharan, A. Molecular cloning and characterization of the rubber elongation factor gene and its promoter sequence from rubber tree (Hevea brasiliensis): A gene involved in rubber biosynthesis. Plant Sci. 2006, 171, 470–480. [Google Scholar] [CrossRef]
- Chow, K.S.; Wan, K.L.; Isa, M.N.; Bahari, A.; Tan, S.H.; Harikrishna, K.; Yeang, H.Y. Insights into rubber biosynthesis from transcriptome analysis of Hevea brasiliensis latex. J. Exp. Bot. 2007, 58, 2429–2440. [Google Scholar] [CrossRef]
- Tang, C.; Yang, M.; Fang, Y.; Luo, Y.; Gao, S.; Xiao, X.; An, Z.; Zhou, B.; Zhang, B.; Tan, X.; et al. The rubber tree genome reveals new insights into rubber production and species adaptation. Nat. Plants 2016, 2, 16073. [Google Scholar] [CrossRef]
- Ding, Z.; Fu, L.; Tan, D.; Sun, X.; Zhang, J. An integrative transcriptomic and genomic analysis reveals novel insights into the hub genes and regulatory networks associated with rubber synthesis in H. brasiliensis. Ind. Crop Prod. 2020, 153, 112562. [Google Scholar] [CrossRef]
- Fonseca, S.; Rosado, A.; Vaughan-Hirsch, J.; Bishopp, A.; Chini, A. Molecular locks and keys: The role of small molecules in phytohormone research. Front. Plant Sci. 2014, 5, 709. [Google Scholar] [CrossRef]
- Coupe, M.; Chrestin, H. Physio-chemical and biochemical mechanisms of hormonal (ethylene) stimulation. In Physiology of Rubber Tree Latex; d’Auzac, J., Jacob, J.L., Chrestin, H., Eds.; CRC Press: Boca Raton, FL, USA, 1989; pp. 295–319. [Google Scholar]
- Faridah, Y.; Siti Arija, M.A.; Ghandimathi, H. Changes in some physiological latex parameters in relation to over exploitation and onset of induced tapping panel dryness. J. Nat. Rubber Res. 1996, 10, 182–186. [Google Scholar]
- Putranto, R.A.; Herlinawati, E.; Rio, M.; Leclercq, J.; Piyatrakul, P.; Gohet, E.; Sanier, C.; Oktavia, F.; Pirrello, J.; Kuswanhadi; et al. Involvement of ethylene in the latex metabolism and tapping panel dryness of Hevea brasiliensis. Int. J. Mol. Sci. 2015, 16, 17885–17908. [Google Scholar] [CrossRef]
- Wang, X.; Wang, D.; Sun, Y.; Yang, Q.; Chang, L.; Wang, L.; Meng, X.; Huang, Q.; Jin, X.; Tong, Z. Comprehensive proteomics analysis of laticifer latex reveals new insights into ethylene stimulation of natural rubber production. Sci. Rep. 2015, 5, 13778. [Google Scholar] [CrossRef]
- Montoro, P.; Wu, S.; Favreau, B.; Herlinawati, E.; Labrune, C.; Martin-Magniette, M.L.; Pointet, S.; Rio, M.; Leclercq, J.; Ismawanto, S.; et al. Transcriptome analysis in Hevea brasiliensis latex revealed changes in hormone signalling pathways during ethephon stimulation and consequent tapping panel dryness. Sci. Rep. 2018, 8, 8483. [Google Scholar] [CrossRef]
- Tungngoen, K.; Viboonjun, U.; Kongsawadworakul, P.; Katsuhara, M.; Julien, J.L.; Sakr, S.; Chrestin, H.; Narangajavana, J. Hormonal treatment of the bark of rubber trees (Hevea brasiliensis) increases latex yield through latex dilution in relation with the differential expression of two aquaporin genes. J. Plant Physiol. 2011, 168, 253–262. [Google Scholar] [CrossRef]
- Laosombut, T.; Arreewichit, P.; Nirapathpongporn, K.; Traiperm, P.; Kongsawadworakul, P.; Viboonjun, U.; Narangajavana, J. Differential expression of methyl jasmonate-responsive genes correlates with laticifer vessel proliferation in phloem tissue of rubber tree (Hevea brasiliensis). J. Plant Growth Regul. 2016, 35, 1049–1063. [Google Scholar] [CrossRef]
- Cao, X.; Yan, J.; Lei, J.; Li, J.; Zhu, J.; Zhang, H. De novo transcriptome sequencing of MeJA-induced Taraxacum koksaghyz Rodin to identify genes related to rubber formation. Sci. Rep. 2017, 7, 15697. [Google Scholar] [CrossRef]
- Liu, J.P.; Hu, J.; Liu, Y.H.; Yang, C.P.; Zhuang, Y.F.; Guo, X.L.; Li, Y.J.; Zhang, L. Transcriptome analysis of Hevea brasiliensis in response to exogenous methyl jasmonate provides novel insights into regulation of jasmonate-elicited rubber biosynthesis. Physiol. Mol. Biol. Plants 2018, 24, 349–358. [Google Scholar] [CrossRef]
- Wu, C.; Lan, L.; Li, Y.; Nie, Z.; Zeng, R. The relationship between latex metabolism gene expression with rubber yield and related traits in Hevea brasiliensis. BMC Genom. 2018, 19, 897. [Google Scholar] [CrossRef]
- Wahler, D.; Gronover, C.S.; Richter, C.; Foucu, F.; Twyman, R.M.; Moerschbacher, B.M.; Fischer, R.; Muth, J.; Prüfer, D. Polyphenoloxidase silencing affects latex coagulation in Taraxacum species. Plant Physiol. 2009, 151, 334–346. [Google Scholar] [CrossRef]
- Dai, L.; Kang, G.; Nie, Z.; Li, Y.; Zeng, R. Comparative proteomic analysis of latex from Hevea brasiliensis treated with ethrel and methyl jasmonate using iTRAQ-coupled two-dimensional LC-MS/MS. J. Proteom. 2016, 132, 167–175. [Google Scholar] [CrossRef]
- Deng, X.; Guo, D.; Yang, S.; Shi, M.; Chao, J.; Li, H.; Peng, S.; Tian, W. Jasmonate signalling in the regulation of rubber biosynthesis in laticifer cells of rubber tree, Hevea brasiliensis. J. Exp. Bot. 2018, 69, 3559–3571. [Google Scholar] [CrossRef]
- Yamashita, S.; Yamaguchi, H.; Waki, T.; Aoki, Y.; Mizuno, M.; Yanbe, F.; Ishii, T.; Funaki, A.; Tozawa, Y.; Miyagi-Inoue, Y.; et al. Identification and reconstitution of the rubber biosynthetic machinery on rubber particles from Hevea brasiliensis. eLife 2016, 5, e19022. [Google Scholar] [CrossRef]
- Brown, D.; Feeney, M.; Ahmadi, M.; Lonoce, C.; Sajari, R.; Di Cola, A.; Frigerio, L. Subcellular localization and interactions among rubber particle proteins from Hevea brasiliensis. J. Exp. Bot. 2017, 68, 5045–5055. [Google Scholar] [CrossRef]
- Dai, L.; Nie, Z.; Kang, G.; Li, Y.; Zeng, R. Identification and subcellular localization analysis of two rubber elongation factor isoforms on Hevea brasiliensis rubber particles. Plant Physiol. Biochem. 2017, 111, 97–106. [Google Scholar] [CrossRef]
- Wang, D.; Xie, Q.; Sun, Y.; Tong, Z.; Chang, L.; Yu, L.; Zhang, X.; Yuan, B.; He, P.; Jin, X.; et al. Proteomic landscape has revealed small rubber particles are crucial rubber biosynthetic machines for ethylene-stimulation in natural rubber production. Int. J. Mol. Sci. 2019, 20, 5082. [Google Scholar] [CrossRef]
- Shahmuradov, I.A.; Umarov, R.K.; Solovyev, V.V. TSSPlant: A new tool for prediction of plant Pol II promoters. Nucl. Acids Res. 2017, 45, e65. [Google Scholar] [CrossRef]
- Sandelin, A.; Alkema, W.; Engström, P.; Wasserman, W.W.; Lenhard, B. (2004). JASPAR: An open-access database for eukaryotic transcription factor binding profiles. Nucl. Acids Res. 2004, 32, D91–D94. [Google Scholar] [CrossRef]
- Rauluseviciute, I.; Riudavets-Puig, R.; Blanc-Mathieu, R.; Castro-Mondragon, J.A.; Ferenc, K.; Kumar, V.; Lemma, R.B.; Lucas, J.; Chèneby, J.; Baranasic, D.; et al. JASPAR 2024: 20th anniversary of the open-access database of transcription factor binding profiles. Nucl. Acids Res. 2024, 52, D174–D182. [Google Scholar] [CrossRef]
- Tian, F.; Yang, D.C.; Meng, Y.Q.; Jin, J.; Gao, G. PlantRegMap: Charting functional regulatory maps in plants. Nucl. Acids Res. 2020, 48, D1104–D1113. [Google Scholar] [CrossRef]
- Yang, Y.; Li, R.; Qi, M. In vivo analysis of plant promoters and transcription factors by agroinfiltration of tobacco leaves. Plant J. 2000, 22, 543–551. [Google Scholar] [CrossRef]
- Strader, L.; Weijers, D.; Wagner, D. Plant transcription factors—Being in the right place with the right company. Curr. Opin. Plant Biol. 2022, 65, 102136. [Google Scholar] [CrossRef]
- Majewska, M.; Wysokińska, H.; Kuźma, Ł.; Szymczyk, P. Eukaryotic and prokaryotic promoter databases as valuable tools in exploring the regulation of gene transcription: A comprehensive overview. Gene 2018, 644, 38–48. [Google Scholar] [CrossRef]
- Li, J.; Han, G.; Sun, C.; Sui, N. Research advances of MYB transcription factors in plant stress resistance and breeding. Plant Signal. Behav. 2019, 14, 1613131. [Google Scholar] [CrossRef]
- Hao, Y.; Zong, X.; Ren, P.; Qian, Y.; Fu, A. Basic helix-loop-helix (bHLH) transcription factors regulate a wide range of functions in Arabidopsis. Int. J. Mol. Sci. 2021, 22, 7152. [Google Scholar] [CrossRef]
- Binder, B.M. Ethylene signaling in plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar] [CrossRef]
- Feng, K.; Hou, X.L.; Xing, G.M.; Liu, J.X.; Duan, A.Q.; Xu, Z.S.; Li, M.Y.; Zhuang, J.; Xiong, A.S. Advances in AP2/ERF super-family transcription factors in plant. Crit. Rev. Biotechnol. 2020, 40, 750–776. [Google Scholar] [CrossRef]
- Han, H.; Wang, C.; Yang, X.; Wang, L.; Ye, J.; Xu, F.; Liao, Y.; Zhang, W. Role of bZIP transcription factors in the regulation of plant secondary metabolism. Planta 2023, 258, 13. [Google Scholar] [CrossRef]
- Luo, J.L.; Zhao, N.; Lu, C.M. Plant Trihelix transcription factors family. Hereditas 2012, 34, 1551–1560. [Google Scholar] [CrossRef]
- Guo, D.; Zhou, Y.; Li, H.L.; Zhu, J.H.; Wang, Y.; Chen, X.T.; Peng, S.Q. Identification and characterization of the abscisic acid (ABA) receptor gene family and its expression in response to hormones in the rubber tree. Sci. Rep. 2017, 7, 45157. [Google Scholar] [CrossRef]
- Deng, X.D.; Fei, X.W.; Huang, J.S.; Zheng, X.Q. Isolation and analysis of rubber elongation factor gene and its 5′-end promoter region. Acta Agron. Sin. 2002, 28, 528–532. [Google Scholar]
- Hellens, R.P.; Allan, A.C.; Friel, E.N.; Bolitho, K.; Grafton, K.; Templeton, M.D.; Karunairetnam, S.; Gleave, A.P.; Laing, W.A. Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant Methods 2005, 1, 13. [Google Scholar] [CrossRef]
- Chao, J.; Wu, S.; Shi, M.; Xu, X.; Gao, Q.; Du, H.; Gao, B.; Guo, D.; Yang, S.; Zhang, S.; et al. Genomic insight into domestication of rubber tree. Nat. Commun. 2023, 14, 4651. [Google Scholar] [CrossRef]
- Fricke, J.; Hillebrand, A.; Twyman, R.M.; Prüfer, D.; Schulze Gronover, C. Abscisic acid-dependent regulation of small rubber particle protein gene expression in Taraxacum brevicorniculatum is mediated by TbbZIP1. Plant Cell Physiol. 2013, 54, 448–464. [Google Scholar] [CrossRef]
- Yokota, S.; Gotoh, T. Effects of rubber elongation factor and small rubber particle protein from rubber-producing plants on lipid metabolism in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2019, 128, 585–592. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Matrix ID | Name | Relative Score a | p-Value b | q-Value b | Start | End | Strand | Predicted Sequence | TF Class |
|---|---|---|---|---|---|---|---|---|---|
| MA2077.1 | PK18474.1 | 1.00 | 5.30 × 10−6 | 0.0185 | −404 | −398 | - | CACGTGC | bHLH |
| MA2057.1 | PK01630.1 | 0.92 | 2.20 × 10−5 | 0.0383 | −270 | −264 | - | GCACATG | bHLH |
| MA2407.1 | PK24978.1 | 0.93 | 8.72 × 10−5 | 0.153 | −1145 | −1139 | - | CACGAGT | bZIP |
| MA2409.1 | ZmbZIP25 | 0.93 | 6.21 × 10−5 | 0.217 | −1107 | −1098 | + | GATCACATCA | bZIP |
| MA2091.1 | PK27109.1 | 0.96 | 2.17 × 10−6 | 0.0076 | −182 | −175 | - | GCCAGCTG | bZIP |
| MA2351.1 | OsEIL3 | 1.00 | 1.19 × 10−5 | 0.0309 | −631 | −624 | + | AGATACAT | EIL |
| MA2417.1 | EREB71 | 0.94 | 7.06 × 10−5 | 0.247 | −1328 | −1319 | - | TACCGGCTAC | ERF/DREB |
| MA2065.1 | PK08602.1 | 0.94 | 2.49 × 10−5 | 0.0871 | −932 | −925 | + | TCACCGCT | ERF/DREB |
| MA2083.1 | PK21166.1 | 0.91 | 3.81 × 10−5 | 0.0642 | −894 | −887 | - | AACCAAAC | MYB |
| MA2083.1 | PK21166.1 | 0.91 | 4.97 × 10−5 | 0.0642 | −793 | −786 | + | AACCAAAC | MYB |
| MA2083.1 | PK21166.1 | 0.91 | 5.66 × 10−5 | 0.0642 | −137 | −130 | - | AACCAAAC | MYB |
| MA2352.1 | PC-MYB1 | 1.00 | 8.17 × 10−6 | 0.0278 | −1677 | −1669 | + | TAACCGTTG | MYB |
| MA2403.1 | DVH24_040989 | 1.00 | 4.74 × 10−5 | 0.153 | −890 | −885 | - | TTAACC | Trihelix |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.-T.; Guo, D.; Zhu, J.-H.; Wang, Y.; Li, H.-L.; An, F.; Tang, Y.-Q.; Peng, S.-Q. Functional Analysis of the HbREF1 Promoter from Hevea brasiliensis and Its Response to Phytohormones. Forests 2024, 15, 276. https://doi.org/10.3390/f15020276
Chen L-T, Guo D, Zhu J-H, Wang Y, Li H-L, An F, Tang Y-Q, Peng S-Q. Functional Analysis of the HbREF1 Promoter from Hevea brasiliensis and Its Response to Phytohormones. Forests. 2024; 15(2):276. https://doi.org/10.3390/f15020276
Chicago/Turabian StyleChen, Lin-Tao, Dong Guo, Jia-Hong Zhu, Ying Wang, Hui-Liang Li, Feng An, Yan-Qiong Tang, and Shi-Qing Peng. 2024. "Functional Analysis of the HbREF1 Promoter from Hevea brasiliensis and Its Response to Phytohormones" Forests 15, no. 2: 276. https://doi.org/10.3390/f15020276
APA StyleChen, L.-T., Guo, D., Zhu, J.-H., Wang, Y., Li, H.-L., An, F., Tang, Y.-Q., & Peng, S.-Q. (2024). Functional Analysis of the HbREF1 Promoter from Hevea brasiliensis and Its Response to Phytohormones. Forests, 15(2), 276. https://doi.org/10.3390/f15020276