
The Different Factors Driving SOC Stability under Different N Addition Durations in a Phyllostachys edulis Forest

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Long-Term Experimental Design
2.2. Sample Collection and Analysis
2.3. Determination of Soil Physicochemical Properties
2.4. Determination of Soil Organic Carbon Fractions
2.5. Determination of Soil Microbial Biomass and Composition
2.6. Determination of Soil Enzyme Activities and Microbial Carbon Use Efficiency
2.7. Statistical Analysis
3. Results
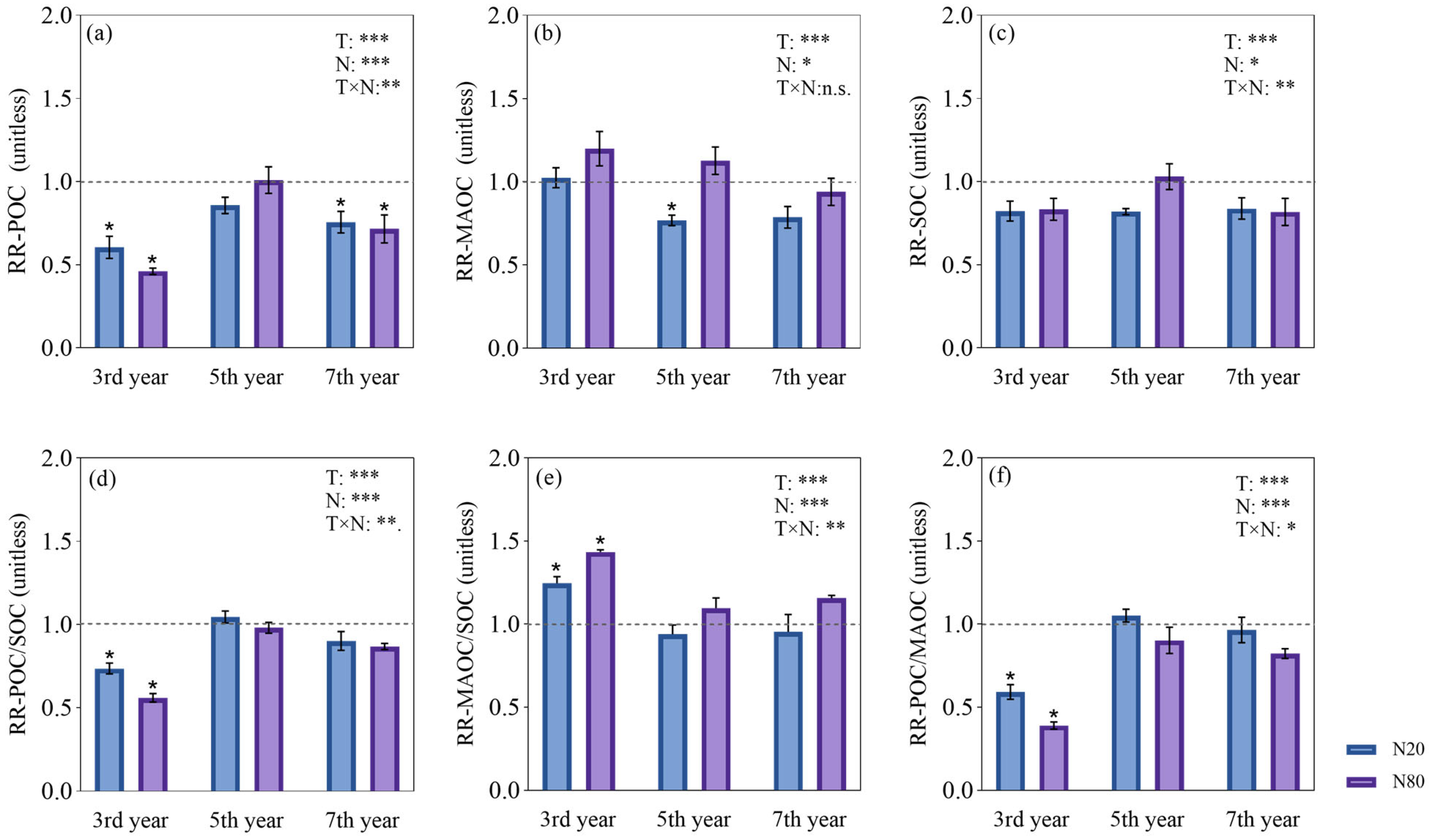
3.1. On the Response of SOC Fractions to Different N Addition Durations
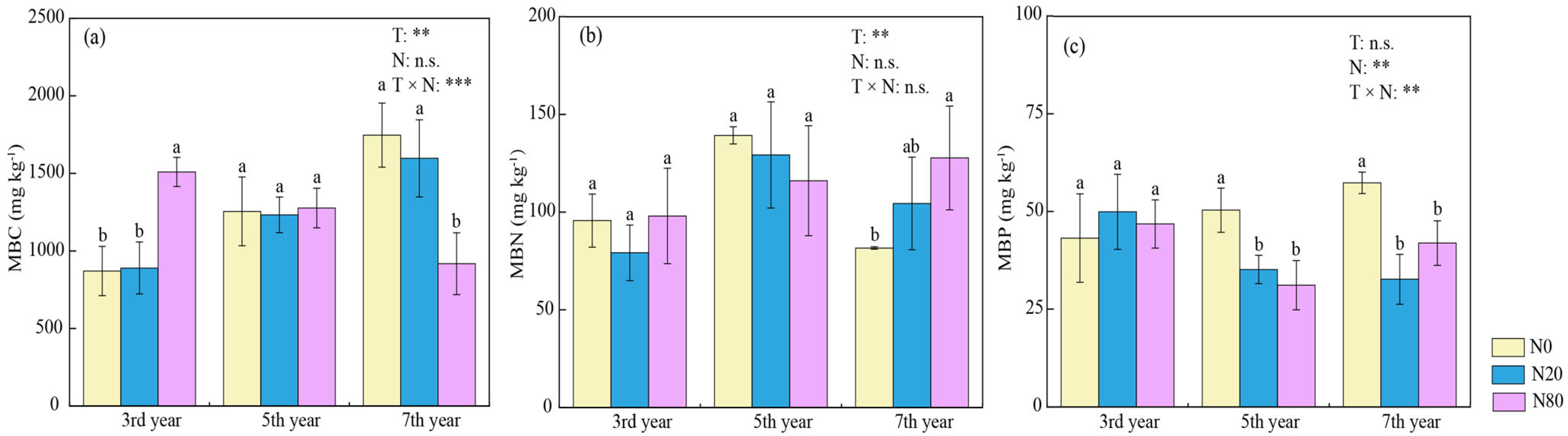
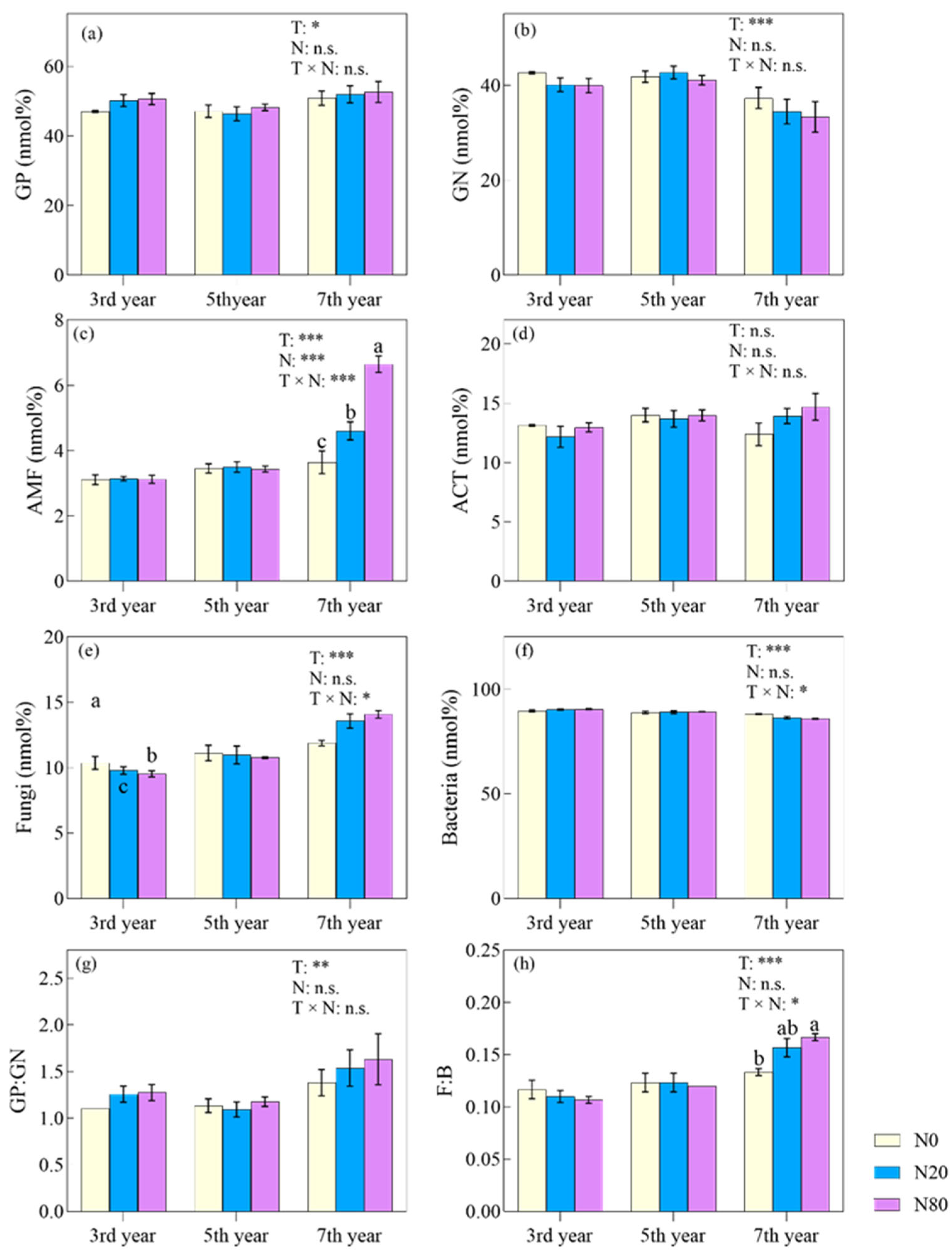
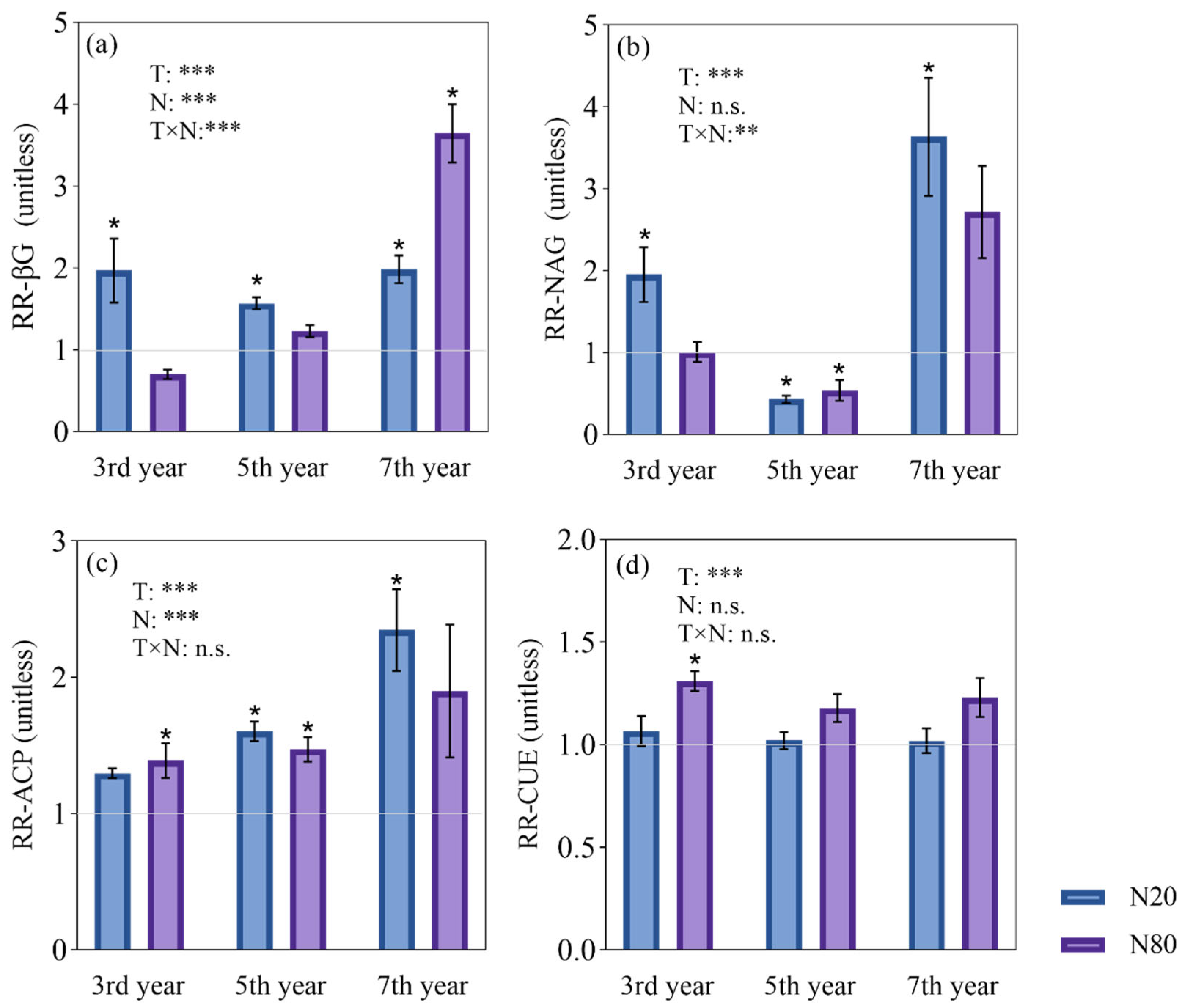
3.2. Effects of Nitrogen Addition on Microbial Characteristics
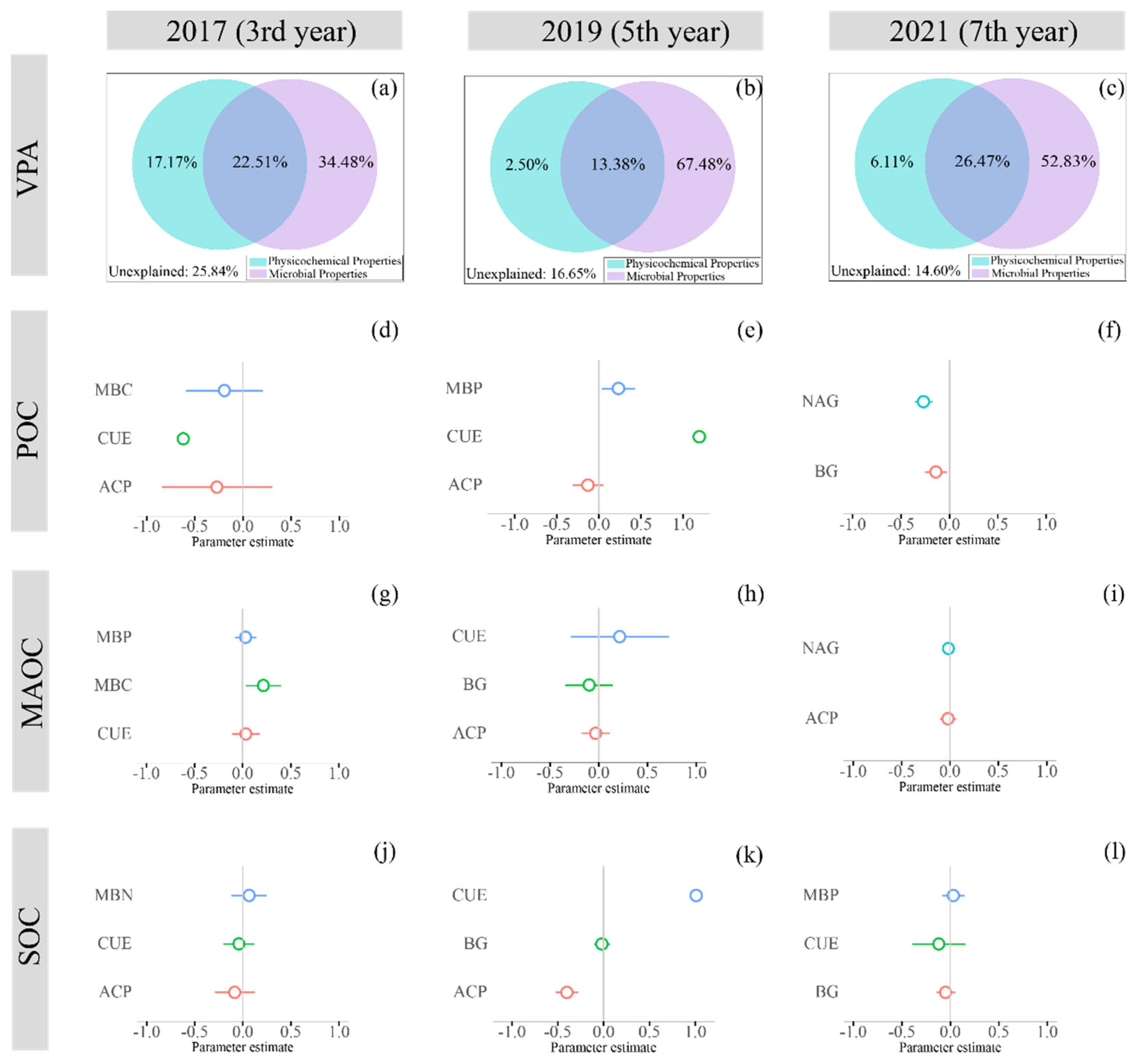
3.3. The Driving Factors of SOC Fractions under Different Durations
4. Discussion
4.1. SOC Fractions Responded Differently under Different N Addition Durations
4.2. Effects of N Addition on Microbial Properties under Different Durations
4.3. The Relationship between Microbial Communities and SOC Fractions under Different Durations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, J.G.; Xiao, W.; Zheng, C.Y.; Zhu, B. Nitrogen addition has contrasting effects on particulate and mineral-associated soil organic carbon in a subtropical forest. Soil Biol. Biochem. 2020, 142, 107708. [Google Scholar] [CrossRef]
- Clark, C.M.; Tilman, D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 2008, 451, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.J.; Zhang, Y.; Han, W.X.; Tang, A.H.; Shen, J.L.; Cui, Z.L.; Vitousek, P.; Erisman, J.M.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.A.; Wieder, W.R.; Bonan, G.B.; Fierer, N.; Raymond, P.A.; Crowther, T.W. Managing uncertainty in soil carbon feedbacks to climate change. Nat. Clim. Chang. 2016, 6, 751–758. [Google Scholar] [CrossRef]
- Jackson, R.B.; Lajtha, K.; Crow, S.E.; Hugelius, G.; Kramer, M.G.; Piñeiro, G. The ecology of soil carbon: Pools, vulnerabilities, and biotic and abiotic controls. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 419–445. [Google Scholar] [CrossRef]
- Guenet, B.; Gabrielle, B.; Chenu, C.; Arrouays, D.; Balesdent, J.; Bernoux, M.; Bruni, E.; Caliman, J.P.; Cardinael, R.; Chen, S.C.; et al. Can N2O emissions offset the benefits from soil organic carbon storage? Glob. Chang. Biol. 2021, 27, 237–256. [Google Scholar] [CrossRef]
- Lavallee, J.M.; Soong, J.L.; Cotrufo, M.F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Glob. Chang. Biol. 2020, 26, 261–273. [Google Scholar] [CrossRef]
- Lugato, E.; Lavallee, J.M.; Haddix, M.L.; Panagos, P.; Cotrufo, M.F. Different climate sensitivity of particulate and mineral associated soil organic matter. Nat. Geosci. 2021, 14, 295–300. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ranalli, M.G.; Haddix, M.L.; Six, J.; Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 2019, 12, 989–994. [Google Scholar] [CrossRef]
- Yu, W.J.; Huang, W.J.; Weintraub-Leff, S.R.; Hall, S.J. Where and why do particulate organic matter (POM) and mineral-associated organic matter (MAOM) differ among diverse soils? Soil Biol. Biochem. 2022, 172, 108756. [Google Scholar] [CrossRef]
- Duan, P.P.; Wang, K.L.; Li, D.J. Nitrogen addition effects on soil mineral-associated carbon differ between the valley and slope in a subtropical karst forest. Geoderma 2023, 116357. [Google Scholar] [CrossRef]
- Zhu, X.M.; Zhang, Z.L.; Liu, D.Y.; Kou, Y.P.; Zheng, Q.; Liu, M.; Xiao, J.; Liu, Q.; Yin, H.J. Differential impacts of nitrogen addition on rhizosphere and bulk-soil carbon sequestration in an alpine shrubland. J. Ecol. 2020, 108, 2309–2320. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, A.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Kallenbach, C.M.; Frey, S.D.; Grandy, A.S. Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 2016, 7, 13630. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.Y.; Luo, J.F.; Fan, J.L.; Liu, D.Y.; He, J.S.; Perveen, N.; Ding, W.X. Responses of soil microbial communities and functions associated with organic carbon mineralization to nitrogen addition in a Tibetan grassland. Pedosphere 2020, 30, 214–225. [Google Scholar] [CrossRef]
- Zumsteg, A.; Bååth, E.; Stierli, B.; Zeyer, J.; Frey, B. Bacterial and fungal community responses to reciprocal soil transfer along a temperature and soil moisture gradient in a glacier forefield. Soil Biol. Biochem. 2013, 61, 121–132. [Google Scholar] [CrossRef]
- Cenini, V.L.; Fornara, D.A.; McMullan, G.; Ternan, N.; Carolan, R.; Crawley, M.J.; Clément, J.; Lavorel, S. Linkages between extracellular enzyme activities and the carbon and nitrogen content of grassland soils. Soil Biol. Biochem 2016, 96, 198–206. [Google Scholar] [CrossRef]
- Feng, X.H.; Qin, S.Q.; Zhang, D.Y.; Chen, P.D.; Hu, J.; Wang, G.Q.; Liu, Y.; Wei, B.; Li, Q.L.; Yang, Y.H.; et al. Nitrogen input enhances microbial carbon use efficiency by altering plant–microbe–mineral interactions. Glob. Chang. Biol. 2022, 28, 4845–4860. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Wallenstein, M.D.; Boot, C.M.; Denef, K.; Paul, E. The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Chang. Biol. 2013, 19, 988–995. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Hu, Y.L.; Hu, A.; Niu, B. Shifts in the dynamic mechanisms of soil organic matter transformation with nitrogen addition: From a soil carbon/nitrogen-driven mechanism to a microbe driven mechanism. Soil Biol. Biochem. 2021, 160, 108355. [Google Scholar] [CrossRef]
- Averill, C.; Waring, B. Nitrogen limitation of decomposition and decay: How can it occur? Glob. Chang. Biol. 2018, 24, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Stone, B.W.; Li, J.H.; Koch, B.J.; Blazewicz, S.J.; Dijkstra, P.; Hayer, M.; Hofmockel, K.S.; Liu, X.J.A.; Mau, R.L.; Morrissey, E.M.; et al. Nutrients cause consolidation of soil carbon flux to small proportion of bacterial community. Nat. Commun. 2021, 12, 3381. [Google Scholar] [CrossRef] [PubMed]
- Mueller, N.D.; Gerber, J.S.; Johnston, M.; Ray, D.K.; Ramankutty, N.; Foley, J.A. Closing yield gaps through nutrient and water management. Nature 2012, 490, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Horn, E.L.; Cooledge, E.C.; Jones, D.L.; Hoyle, F.C.; Brailsford, F.L.; Murphy, D.V. Addition of base cations increases microbial carbon use efficiency and biomass in acidic soils. Soil Biol. Biochem. 2021, 161, 108392. [Google Scholar] [CrossRef]
- Zeng, Q.X.; Yuan, X.C.; Zhou, J.C.; Wu, J.M. Effects of nitrogen addition on the kinetic parameters of soil acid phosphomonoesterase in a Moso bamboo forest. Chin. J. Appl. Ecol. 2022, 33, 2178–2186. (In Chinese) [Google Scholar]
- Cheng, L.; Lin, K.M.; Zhou, J.C.; Zhang, Q.F.; Zeng, X.M.; Ji, Y.H.; Zheng, Y.; Xu, J.G.; Chen, Y.M. Effects of nitrogen deposition on the concentration and spectral characteristics of dissolved organic matter in soil in Moso bamboo plantations. Chin. J. Appl. Ecol. 2019, 30, 1754–1762. (In Chinese) [Google Scholar]
- Chen, J.; Luo, T.S.; Zhou, Z.; Xu, H.; Chen, D.X.; Li, Y.D.; Jiang, Z.Y. Research advances in nitrogen deposition effects on microbial processes involved in soil nitrogen cycling in tropical and subtropical forests. Acta Ecol. Sin. 2020, 40, 8528–8538. (In Chinese) [Google Scholar]
- Yu, G.R.; Jia, Y.L.; He, N.P.; Zhu, J.X.; Chen, Z.; Wang, Q.F.; Piao, S.L.; Liu, X.J.; He, H.L.; Guo, X.B.; et al. Stabilization of atmospheric nitrogen deposition in China over the past decade. Nat. Geosci. 2019, 12, 424–429. [Google Scholar] [CrossRef]
- Jia, Y.L.; Yu, G.R.; Gao, Y.N.; He, N.P.; Wang, Q.F.; Jiao, C.C.; Zuo, Y. Global inorganic nitrogen dry deposition inferred from ground- and space-based measurements. Sci. Rep. 2016, 6, 19810. [Google Scholar] [CrossRef]
- Robertson, A.D.; Paustian, K.; Ogle, S.; Wallenstein, M.D.; Lugato, E.; Cotrufo, M.F. Unifying soil organic matter formation and persistence frameworks: The MEMS model. Biogeosciences 2019, 16, 1225–1248. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- White, D.C.; Davis, W.M.; Nickels, J.S.; King, J.D.; Bobbie, R.J. Determination of the sedimentary microbial biomass by extractible lipid phosphate. Oecologia 1979, 40, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Hobbs, P.J.; Frostegård, Å. Changes in soil fungal: Bacterial biomass ratios following reductions in the intensity of management of an upland grassland. Biol. Fertil. Soils 1996, 22, 261–264. [Google Scholar] [CrossRef]
- Joergensen, R.G. Phospholipid fatty acids in soil-drawbacks and future prospects. Biol. Fertil. Soils 2022, 58, 1–6. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Turner, B.L.; Talbot, J.M.; Waring, B.G.; Powers, J.S.; Kuske, C.R.; Moorhead, D.L.; Shah, J.J.F. Stoichiometry of microbial carbon use efficiency in soils. Ecol. Monogr. 2016, 86, 172–189. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef]
- Schmidt-Nielsen, K. Scaling: Why Is Animal Size So Important? Cambridge University Press: New York, NY, USA, 1984. [Google Scholar]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Burnham, K.; Anderson, D. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Bartoń, K. MuMIn: Multi-Model Inference. R Package v. 1.43.17. 2020. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 15 May 2023).
- Shi, L.; Dech, J.P.; Yao, H.; Zhao, P.; Shu, Y.; Zhou, M. The effects of nitrogen addition on dissolved carbon in boreal forest soils of northeastern China. Sci. Rep. 2019, 9, 8274. [Google Scholar] [CrossRef]
- Tang, B.; Rocci, K.S.; Lehmann, A.; Rillig, M.C. Nitrogen increases soil organic carbon accrual and alters its functionality. Glob. Chang. Biol. 2023, 29, 1971–1983. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, D.; Feng, W.; Niu, S.; Plante, A.; Luo, Y.; Wang, K. Different responses of soil organic carbon fractions to additions of nitrogen. Eur. J. Soil Sci. 2018, 69, 1098–1104. [Google Scholar] [CrossRef]
- Wan, D.; Ma, M.K.; Peng, N.; Luo, X.S.; Chen, W.L.; Cai, P.; Wu, L.H.; Pan, H.B.; Chen, J.B.; Yu, G.H.; et al. Effects of long-term fertilization on calcium-associated soil organic carbon: Implications for C sequestration in agricultural soils. Sci. Total Environ. 2021, 772, 145037. [Google Scholar] [CrossRef]
- Zak, D.R.; Freedman, Z.B.; Upchurch, R.A.; Steffens, M.; Kögel-Knabner, I. Anthropogenic N deposition increases soil organic matter accumulation without altering its biochemical composition. Glob. Chang. Biol. 2017, 23, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Argiroff, W.A.; Zak, D.R.; Upchurch, R.A.; Salley, S.O.; Grandy, A.S. Anthropogenic N deposition alters soil organic matter biochemistry and microbial communities on decaying fine roots. Glob. Chang. Biol. 2019, 25, 4369–4382. [Google Scholar] [CrossRef]
- Lai, Y.N.; Zhong, Q.L.; Li, B.Y.; Cheng, D.L.; Yu, H.; Xu, C.B.; Xue, X.H.; Zheng, W.T. Effect of nitrogen addition on the growth of Phyllostachys edulis and evergreen broad-leaved trees in different Vegetation Types. Prot. For. Sci. Technol. 2021, 2, 1–7. (In Chinese) [Google Scholar]
- Niu, L.M.; Qin, H.; Xu, Q.F.; Wu, Q.F.; Miao, J.J.; Peng, D.C. Effects of Long-term plantation of Phyllostachys edulis on evolution of arbuscular mycorrhizal fungus community in soil. Acta Pet. Sin. 2017, 54, 722–734. (In Chinese) [Google Scholar]
- Ye, C.L.; Chen, D.M.; Hall, S.J.; Pan, S.; Yan, X.B.; Bai, T.S.; Guo, H.; Zhang, Y.; Bai, Y.F.; Hu, S.J. Reconciling multiple impacts of nitrogen enrichment on soil carbon: Plant, microbial and geochemical controls. Ecol. Lett. 2018, 21, 1162–1173. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef]
- Liang, C.; Amelung, W.; Lehmann, J.; Kästner, M. Quantitative assessment of microbial necromass contribution to soil organic matter. Glob. Chang. Biol. 2019, 25, 3578–3590. [Google Scholar] [CrossRef]
- Treseder, K.K. A meta-analysis of mycorrhizal responses to nitrogen, phosphorus, and atmospheric CO2 in field studies. New Phytol. 2004, 164, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Feng, J.; Han, M.; Zhu, B. Responses of arbuscular mycorrhizal fungi to nitrogen addition: A meta-analysis. Glob. Chang. Biol. 2020, 26, 7229–7241. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.X.; Zhong, X.J.; Lin, F.; Liu, C.C.; Yang, L.M.; Wang, M.H.; Chen, G.S.; Chen, Y.M.; Yang, Y.S. Responses of soil phosphorus fractions after nitrogen addition in a subtropical forest ecosystem: Insights from decreased Fe and Al oxides and increased plant roots. Geoderma 2019, 337, 246–255. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Wang, C.K.; Zheng, M.H.; Jiang, L.F.; Luo, Y.Q. Patterns and mechanisms of responses by soil microbial communities to nitrogen addition. Soil Biol. Biochem. 2017, 115, 433–441. [Google Scholar] [CrossRef]
- Allison, S.D.; Czimczik, C.I.; Treseder, K.K. Microbial activity and soil respiration under nitrogen addition in Alaskan boreal forest. Glob. Chang. Biol. 2008, 14, 1156–1168. [Google Scholar] [CrossRef]
- Sistla, S.A.; Schimel, J.P. Seasonal patterns of microbial extracellular enzyme activities in an arctic tundra soil: Identifying direct and indirect effects of long-term summer warming. Soil Biol. Biochem. 2013, 66, 119–129. [Google Scholar] [CrossRef]
- Manzoni, S.; Taylor, P.; Richter, A.; Porporato, A.; Ågren, G.I. Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytol. 2012, 196, 79–91. [Google Scholar] [CrossRef]
- Wang, C.; Qu, L.R.; Yang, L.M.; Liu, D.W.; Morrissey, E.; Miao, R.H.; Liu, Z.P.; Wang, Q.K.; Fang, Y.T.; Bai, E. Large-scale importance of microbial carbon use efficiency and necromass to soil organic carbon. Glob. Chang. Biol. 2021, 27, 2039–2048. [Google Scholar] [CrossRef]
- Samson, M.-È.; Chantigny, M.H.; Vanasse, A.; Menasseri-Aubry, S.; Royer, I.; Angers, D.A. Management practices differently affect particulate and mineral-associated organic matter and their precursors in arable soils. Soil Biol. Biochem. 2022, 148, 107867. [Google Scholar] [CrossRef]
- Almeida, L.F.J.; Souza, I.F.; Hurtarte, L.C.C.; Teixeira, P.P.D.C.; Inagaki, T.M.; Silva, I.; Mueller, C.M. Forest litter constraints on the pathways controlling soil organic matter formation. Soil Biol. Biochem. 2021, 163, 108447. [Google Scholar] [CrossRef]
- Tian, J.; Zong, N.; Hartley, I.P.; He, N.P.; Zhang, J.J.; Powlson, D.; Zhou, J.Z.; Kuzyakov, Y.; Zhang, F.S.; Yu, G.R.; et al. Microbial metabolic response to winter warming stabilizes soil carbon. Glob. Chang. Biol. 2021, 27, 2011–2028. [Google Scholar] [CrossRef] [PubMed]
- DeForest, J.L.; Moorhead, D.L. Effects of elevated pH and phosphorus fertilizer on soil C, N and P enzyme stoichiometry in an acidic mixed mesophytic deciduous forest. Soil Biol. Biochem. 2020, 150, 107996. [Google Scholar] [CrossRef]
- Fanin, N.; Mooshammer, M.; Sauvadet, M.; Meng, C.; Alvarez, G.; Bernard, L.; Bertrand, I.; Blagodatskaya, E.; Bon, L.; Fontaine, S.; et al. Soil enzymes in response to climate warming: Mechanisms and feedbacks. Funct. Ecol. 2022, 36, 1378–1395. [Google Scholar] [CrossRef]
- Poeplau, C.; Helfrich, M.; Dechow, R.; Szoboszlay, M.; Tebbe, C.C.; Don, A.; Greiner, B.; Zopf, D.; Thumm, U.; Korevaar, H.; et al. Increased microbial anabolism contributes to soil carbon sequestration by mineral fertilization in temperate grasslands. Soil Biol. Biochem. 2019, 130, 167–176. [Google Scholar] [CrossRef]
- Soares, M.; Rousk, J. Microbial growth and carbon use efficiency in soil: Links to fungal-bacterial dominance, SOC-quality, and stoichiometry. Soil Biol. Biochem. 2019, 131, 195–205. [Google Scholar] [CrossRef]
- Tian, D.; Jiang, L.; Ma, S.H.; Fang, W.J.; Schmid, B.; Xu, L.C.; Zhu, J.X.; Li, P.; Losapio, G.; Jing, X.; et al. Effects of nitrogen deposition on soil microbial communities in temperate and subtropical forests in China. Sci. Total Environ. 2017, 607–608, 1367–1375. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Zeng, Q.; Su, X.; Zheng, W.; Zhang, Q.; Chen, Y. The Different Factors Driving SOC Stability under Different N Addition Durations in a Phyllostachys edulis Forest. Forests 2023, 14, 1890. https://doi.org/10.3390/f14091890
Wu Y, Zeng Q, Su X, Zheng W, Zhang Q, Chen Y. The Different Factors Driving SOC Stability under Different N Addition Durations in a Phyllostachys edulis Forest. Forests. 2023; 14(9):1890. https://doi.org/10.3390/f14091890
Chicago/Turabian StyleWu, Yue, Quanxin Zeng, Xianchu Su, Wei Zheng, Qiufang Zhang, and Yuehmin Chen. 2023. "The Different Factors Driving SOC Stability under Different N Addition Durations in a Phyllostachys edulis Forest" Forests 14, no. 9: 1890. https://doi.org/10.3390/f14091890
APA StyleWu, Y., Zeng, Q., Su, X., Zheng, W., Zhang, Q., & Chen, Y. (2023). The Different Factors Driving SOC Stability under Different N Addition Durations in a Phyllostachys edulis Forest. Forests, 14(9), 1890. https://doi.org/10.3390/f14091890