
Recent Advances in Flower Color and Fragrance of Osmanthus fragrans


Abstract
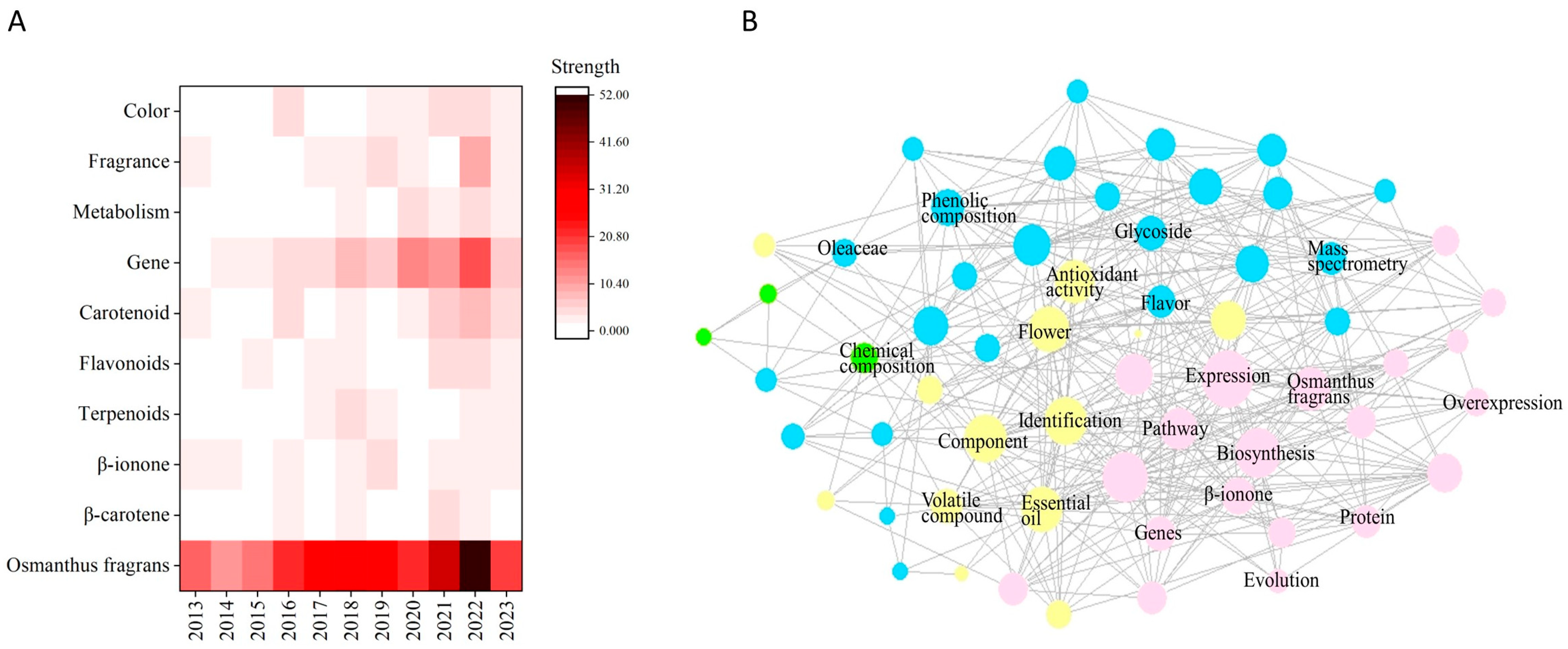
1. Introduction
1.1. Osmanthus fragrans Flower Cultivar Diversity
1.2. Osmanthus Resource Distribution
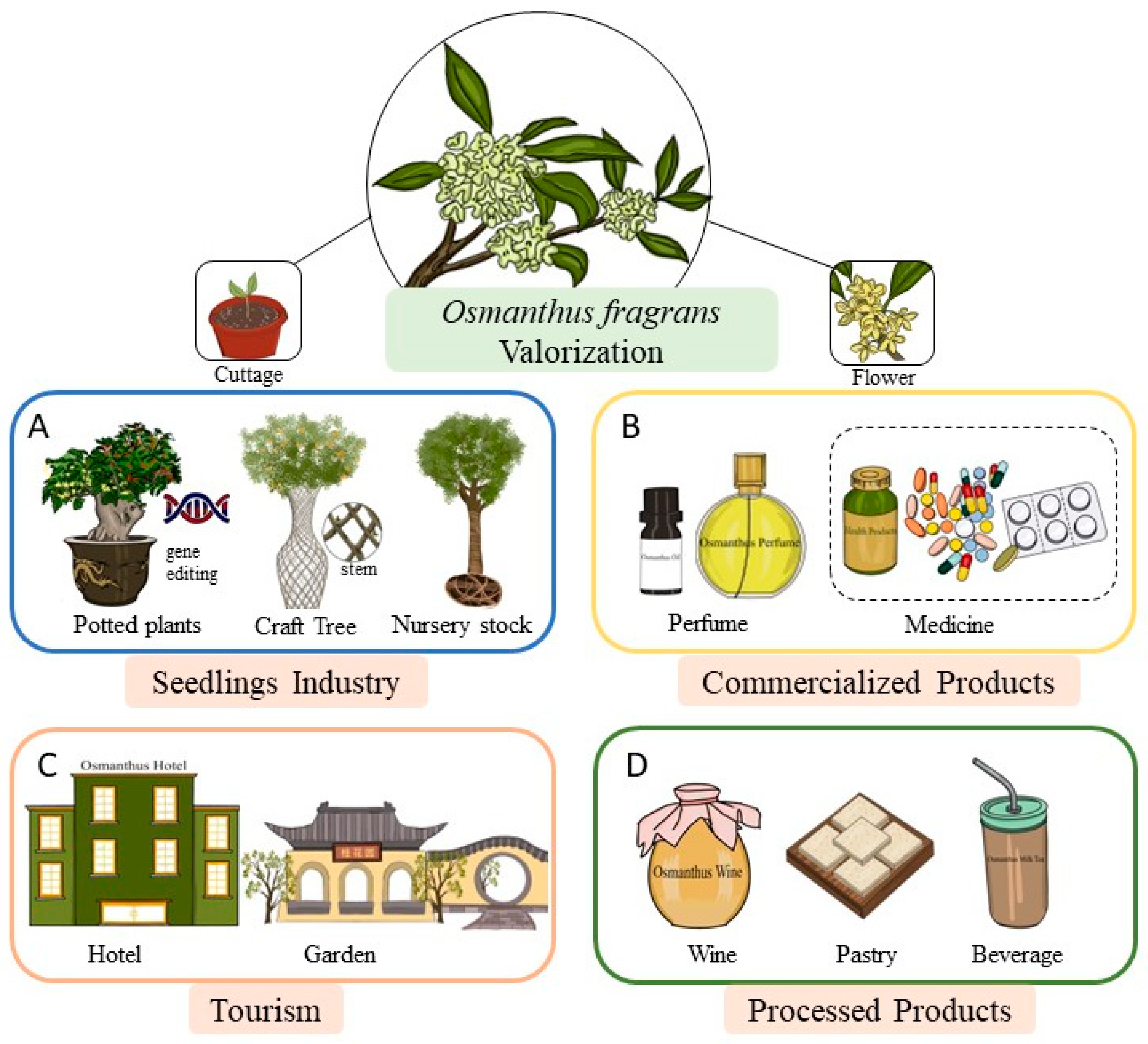
1.3. The Value and Utilization of Osmanthus fragrans
2. Osmanthus fragrans Flower Color
2.1. Major Compounds of Coloration in Osmanthus fragrans
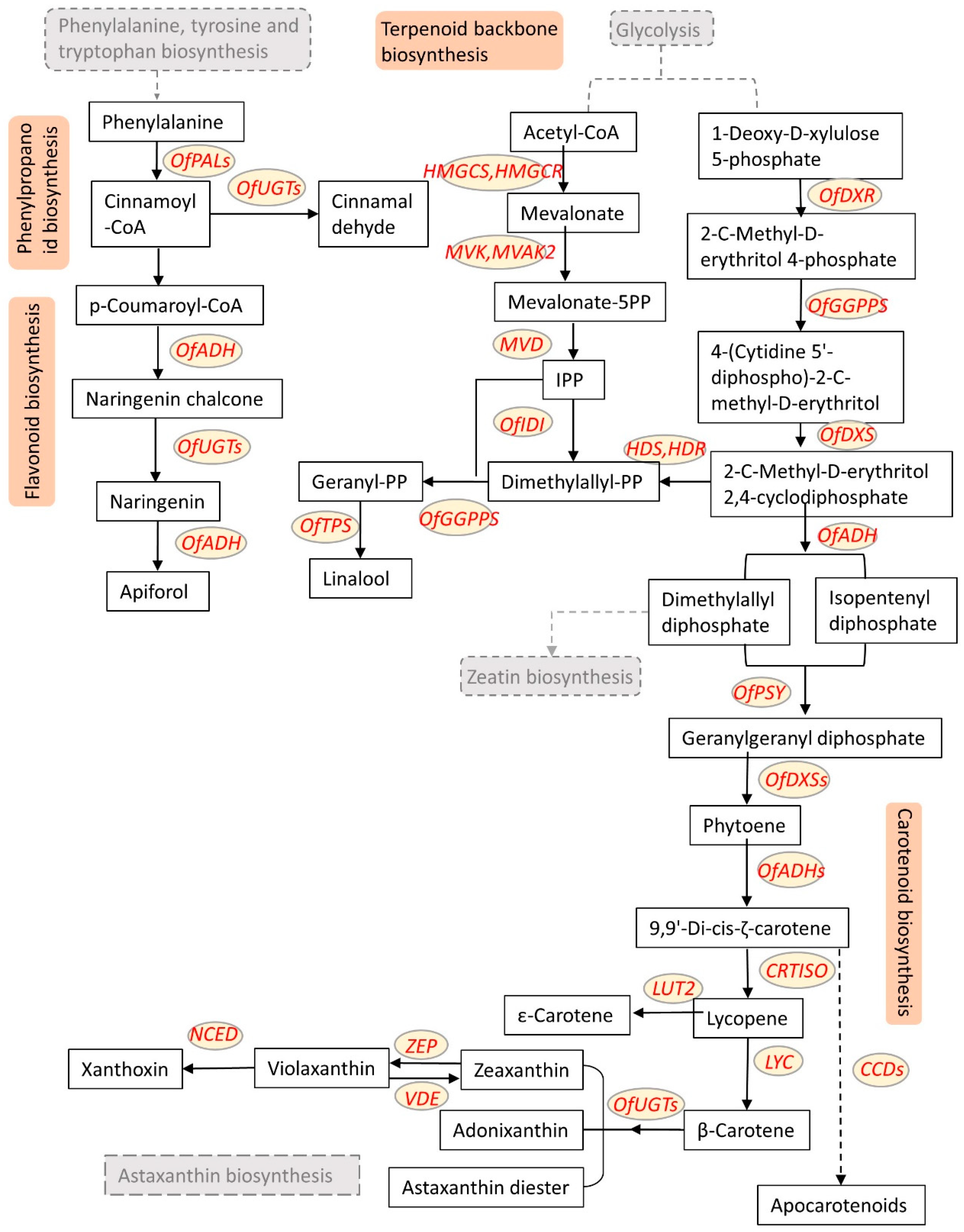
2.1.1. Carotenoids
2.1.2. Flavonoids
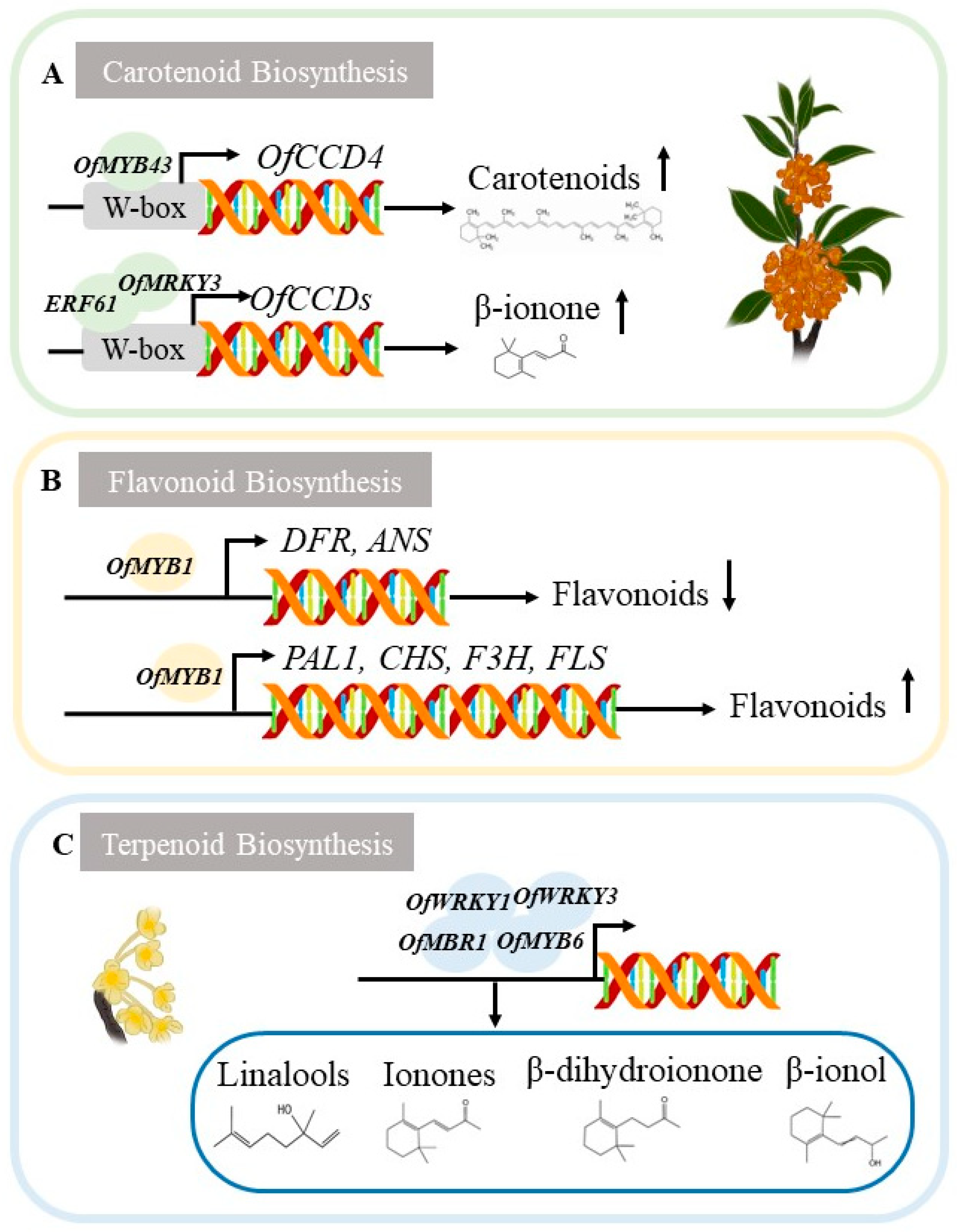
2.2. Regulation of Genes Involved in Osmanthus fragrans Flower Color Formation
3. Osmanthus fragrans Floral Fragrance
3.1. Fragrance Component Analysis
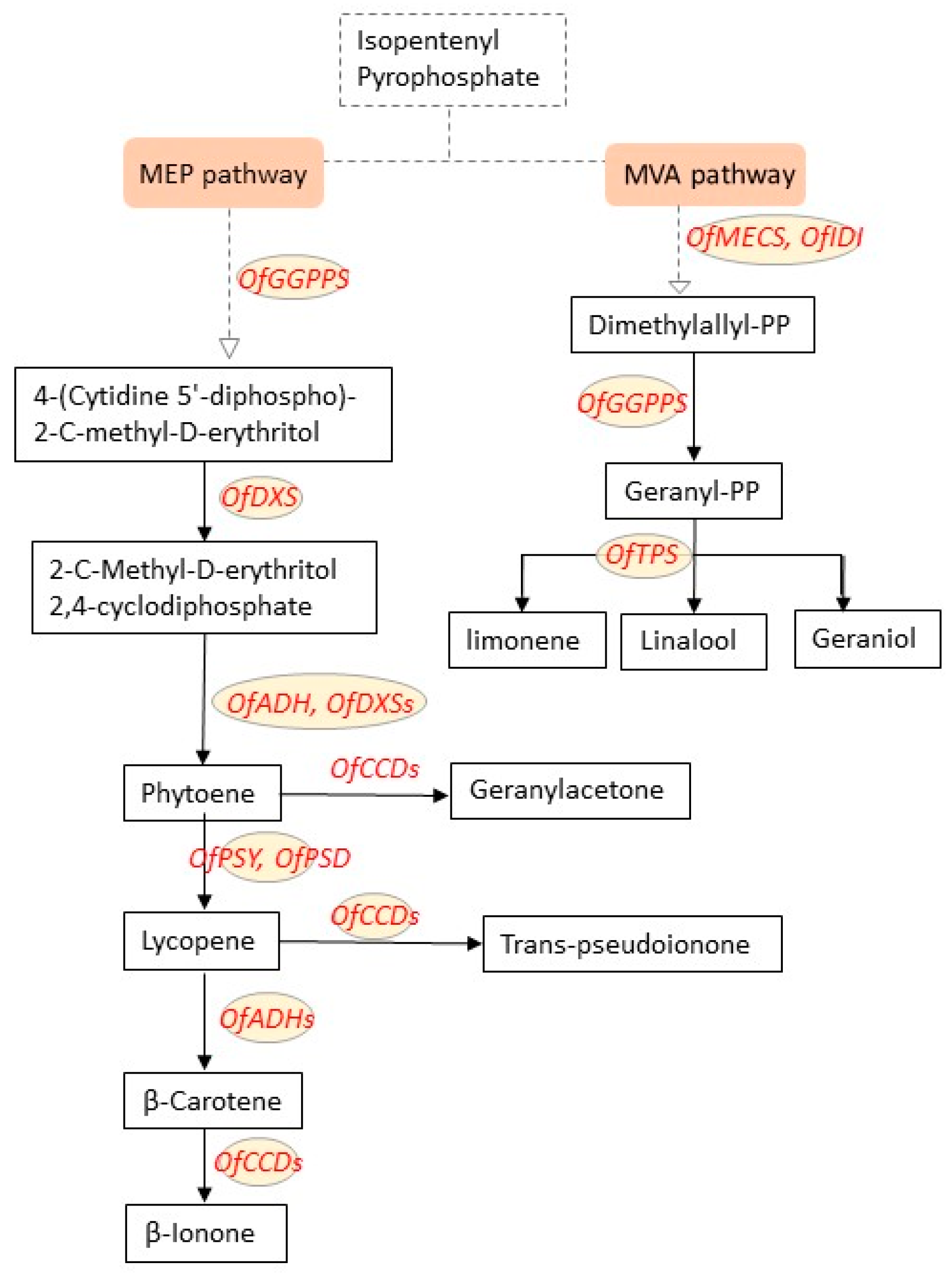
3.1.1. Terpenoids
3.1.2. Fatty Acid Compounds
3.2. Regulation of Genes Related to Fragrance Formation
4. The Relationship between Color and Floral Fragrance
5. High Value-Added Products of Osmanthus fragrans
6. Conclusions and Prospect
6.1. Conclusions
6.2. Prospect
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, B.J.; Luan, F.; Bao, Y.W.; Peng, X.; Rao, Z.L.; Tang, Q.; Zeng, N. Traditional uses, phytochemical constituents and pharmacological properties of Osmanthus fragrans: A review. J. Ethnopharmacol. 2022, 293, 25. [Google Scholar] [CrossRef]
- Zhao, H.B.; Hao, R.M.; Hu, S.Q. Geographic distribution and population characteristics of Osmanthus fragrans. Acta Hortic. Sin. 2015, 42, 1760–1770. [Google Scholar]
- Zang, D.K.; Xiang, Q.B.; Liu, Y.L.; Hao, R.M. The studying history and the application to International Cultivar Registration Authority of sweet osmanthus (Osmanthus fragrans Lour.). J. Plant Resour. Environ. 2003, 12, 49–53. [Google Scholar]
- Liu, Y.L. Cultivar classification of Osmanthus fragrans Lour. and the development of germplasm resources of Osmanthus Lour. J. Plant Resour. Environ. 1993, 2, 44–48. [Google Scholar]
- Cui, Q.; Huang, J.H.; Wu, F.; Li, D.Z.; Zheng, L.Q.; Hu, G.; Hu, S.Q.; Zhang, L. Biochemical and transcriptomic analyses reveal that critical genes involved in pigment biosynthesis influence leaf color changes in a new sweet osmanthus cultivar ‘Qiannan Guifei’. PeerJ 2021, 9, 28. [Google Scholar] [CrossRef]
- Xiang, Q.; Liu, L.; Wu, X. Studies on the Cultivar Classification of Osmanthus. In Proceedings of the 5th International Symposium on the Taxonomy of Cultivated Plants, Wageningen, The Netherlands, 15–19 October 2007. [Google Scholar]
- Kong, F.; Tang, L.; He, H.; Yang, F.X.; Tao, J.; Wang, W.C. Assessing the impact of climate change on the distribution of Osmanthus fragrans using Maxent. Environ. Sci. Pollut. Res. 2021, 28, 34655–34663. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.R.; Fan, D.M.; Guo, S.Q.; Li, D.Z.; Zhang, Z.Y. Development of 29 microsatellite markers for Osmanthus fragrans (oleaceae), a traditional fragrant flowering tree of china. Am. J. Bot. 2011, 98, E356–E359. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.Y. History and cultural significance of sweet Osmanthus in Chinese gardenin. J. Beijing For. Univ. (Soc. Sci.) 2005, 25–29. [Google Scholar]
- Shang, F.D.; Yi, Y.J.; Xiang, Q.B. Osmanthus culture in China. J. Henan Univ. 2003, 136–139. [Google Scholar] [CrossRef]
- Fu, C.C.; Xu, F.Y.; Qian, Y.C.; Koo, H.L.; Duan, Y.F.; Weng, G.M.; Fan, T.P.; Chen, M.X.; Zhu, F.Y. Secondary metabolites of Osmanthus fragrans: Metabolism and medicinal value. Front. Pharmacol. 2022, 13, 922204. [Google Scholar] [CrossRef]
- Wang, C.R.; Liu, C.; Yuan, B.; Zeng, X.L.; Wang, C.Y. Cultivation history, traditional landscape arrangement and auspicious culture of Osmanthus fragrans in Chinese ancient garden. In Proceedings of the 29th International Horticultural Congress on Horticulture—Sustaining Lives, Livelihoods and Landscapes (IHC)/International Symposium on Impact of Asia-Pacific Horticulture—Resources, Technology and Social Welfare, Brisbane, Australia, 17–22 August 2014. [Google Scholar]
- Wang, Y.G.; Fu, J.X.; Zhang, C.; Zhao, H.B. HPLC-DAD-ESI-MS analysis of flavonoids from leaves of different cultivars of sweet osmanthus. Molecules 2016, 21, 1224. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.G.; Zhang, C.; Dong, B.; Fu, J.X.; Hu, S.Q.; Zhao, H.B. Carotenoid accumulation and its contribution to flower coloration of Osmanthus fragrans. Front. Plant. Sci. 2018, 9, 17. [Google Scholar] [CrossRef]
- Liu, J.; Nakamura, S.; Xu, B.; Matsumoto, T.; Ohta, T.; Fujimoto, K.; Ogawa, K.; Fukaya, M.; Miyake, S.; Yoshikawa, M.; et al. Chemical structures of constituents from the flowers of Osmanthus fragrans var. aurantiacus. J. Nat. Med. 2015, 69, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Wang, B.; Wang, M.; Wang, M. Ultrasound-assisted extraction of Osmanthus fragrans fruit oil and evaluation of its fatty acid composition, physicochemical properties and antioxidant activity. J. Appl. Res. Med. Aroma. 2021, 25, 100331. [Google Scholar] [CrossRef]
- Hermanns, A.S.; Zhou, X.; Xu, Q.; Tadmor, Y.; Li, L. Carotenoid pigment accumulation in horticultural plants. Hortic. Plant J. 2020, 6, 343–360. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Hashimoto, H.; Uragami, C.; Yukihira, N.; Horiuchi, K.; Cogdell, R.J. Ultrafast laser spectroscopic studies on carotenoids in solution and on those bound to photosynthetic pigment-protein complexes. Methods Enzymol. 2022, 674, 1–51. [Google Scholar] [CrossRef]
- Guo, S.; Sun, H.; Zhang, H.; Liu, J.; Ren, Y.; Gong, G.; Jiao, C.; Zheng, Y.; Yang, W.; Fei, Z.; et al. Comparative transcriptome analysis of cultivated and wild watermelon during fruit development. PLoS ONE 2015, 10, e0130267. [Google Scholar] [CrossRef]
- Han, Y.J.; Wang, X.H.; Chen, W.C.; Dong, M.F.; Yuan, W.J.; Liu, X.; Shang, F.D. Differential expression of carotenoid-related genes determines diversified carotenoid coloration in flower petal of Osmanthus fragrans. Tree Genet. Genomes 2014, 10, 329–338. [Google Scholar] [CrossRef]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.-H.; Jaremko, M. Important flavonoids and their role as a therapeutic agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef]
- Lu, W.; Shi, Y.; Wang, R.; Su, D.; Tang, M.; Liu, Y.; Li, Z. Antioxidant activity and healthy benefits of natural pigments in fruits: A review. Int. J. Mol. Sci. 2021, 22, 4945. [Google Scholar] [CrossRef]
- Yu, J.; Lou, Q.; Zheng, X.; Cui, Z.; Fu, J. Sequential combination of microwave- and ultrasound-assisted extraction of total flavonoids from Osmanthus fragrans Lour. flowers. Molecules 2017, 22, 2216. [Google Scholar] [CrossRef]
- Zhao, D.Q.; Tao, J. Recent advances on the development and regulation of flower color in ornamental plants. Front. Plant Sci. 2015, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Iwashina, T. Contribution to flower colors of flavonoids including anthocyanins: A review. Nat. Prod. Commun. 2015, 10, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, M.; Nakatsuka, T. Genetic engineering of flavonoid pigments to modify flower color in floricultural plants. Biotechnol. Lett. 2011, 33, 433–441. [Google Scholar] [CrossRef]
- Hao, J.F.; Li, Y.Y.; Jia, Y.S.; Wang, Z.J.; Rong, R.; Bao, J.; Zhao, M.Q.; Fu, Z.H.; Ge, G.T. Comparative analysis of major flavonoids among parts of Lactuca indica during different growth periods. Molecules 2021, 26, 7445. [Google Scholar] [CrossRef]
- Zhou, J.L.; Fang, X.Y.; Wang, J.Q.; Zhao, L.G.; Li, Y.; Tang, F.; Yue, Y.D. Structures and bioactivities of seven flavonoids from Osmanthus fragrans ‘Jinqiu’ essential oil extraction residues. Nat. Prod. Res. 2018, 32, 588–591. [Google Scholar] [CrossRef]
- Wu, L.C.; Chang, L.H.; Chen, S.H.; Fan, N.C.; Ho, J.A.A. Antioxidant activity and melanogenesis inhibitory effect of the acetonic extract of Osmanthus fragrans: A potential natural and functional food flavor additive. LWT-Food Sci. Technol. 2009, 42, 1513–1519. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, M.; Jin, J.; Zhao, L.; Xu, Z. Anthocyanins and their biosynthetic genes in three novel-colored Rosa rugosa cultivars and their parents. Plant Physiol. Bioch. 2018, 129, 421–428. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, T.; Dai, Z.; Dai, X.; Li, W.; Cao, M.; Li, C.; Tsai, W.; Wu, X.; Zhai, J. Comparative transcriptomics provides insight into floral color polymorphism in a Pleione limprichtii orchid population. Int. J. Mol. Sci. 2019, 21, 247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, Y.C.; Wang, Y.G.; Fu, J.X.; Zhao, H.B. Cloning and expression analysis of carotenoid isomerase gene in Osmanthus fragrans. Biotechnol. Bull. 2017, 33, 89. [Google Scholar]
- Auldridge, M.E.; McCarty, D.R.; Klee, H.J. Plant carotenoid cleavage oxygenases and their apocarotenoid products. Curr. Opin. Plant Biol. 2006, 9, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Auldridge, M.E.; Block, A.; Vogel, J.T.; Dabney-Smith, C.; Mila, I.; Bouzayen, M.; Magallanes-Lundback, M.; DellaPenna, D.; McCarty, D.R.; Klee, H.J. Characterization of three members of the Arabidopsis carotenoid cleavage dioxygenase family demonstrates the divergent roles of this multifunctional enzyme family. Plant J. 2006, 45, 982–993. [Google Scholar] [CrossRef]
- Floss, D.S.; Walter, M.H. Role of carotenoid cleavage dioxygenase 1 (CCD1) in apocarotenoid biogenesis revisited. Plant Signal. Behav. 2009, 4, 172–175. [Google Scholar] [CrossRef]
- Rubio, A.; Rambla, J.L.; Santaella, M.; Gómez, M.; Orzaez, D.; Granell, A.; Gómez-Gómez, L. Cytosolic and plastoglobule-targeted carotenoid dioxygenases from Crocus sativus are both involved in beta-ionone release. J. Biol. Chem. 2008, 283, 24816–24825. [Google Scholar] [CrossRef]
- Han, Y.; Wang, H.; Wang, X.; Li, K.; Dong, M.; Li, Y.; Zhu, Q.; Shang, F. Mechanism of floral scent production in Osmanthus fragrans and the production and regulation of its key floral constituents, beta-ionone and linalool. Hortic. Res. 2019, 6, 106. [Google Scholar] [CrossRef]
- Zeng, X.L. Research of TPS and CCD function analysis and their influence on petal color and scent in Osmanthus fragrans Lour. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2016. [Google Scholar]
- Han, Y.J.; Lu, M.M.; Yue, S.M.; Li, K.; Dong, M.F.; Liu, L.X.; Wang, H.Y.; Shang, F.D. Comparative methylomics and chromatin accessibility analysis in Osmanthus fragrans uncovers regulation of genic transcription and mechanisms of key floral scent production. Hortic. Res.-Engl. 2022, 9, 19. [Google Scholar] [CrossRef]
- Xi, W.; He, Y.H.; Zhu, L.L.; Hu, S.Y.; Xiong, S.Y.; Zhang, Y.; Zou, J.J.; Chen, H.G.; Wang, C.Y.; Zheng, R.R. CPTA treatment reveals potential transcription factors associated with carotenoid metabolism in flowers of Osmanthus fragrans. Hortic. Plant J. 2021, 7, 479–487. [Google Scholar] [CrossRef]
- Han, Y.J.; Wu, M.; Cao, L.Y.; Yuan, W.J.; Dong, M.F.; Wang, X.H.; Chen, W.C.; Shang, F.D. Characterization of OfWRKY3, a transcription factor that positively regulates the carotenoid cleavage dioxygenase gene OfCCD4 in Osmanthus fragrans. Plant Mol. Biol. 2016, 91, 485–496. [Google Scholar] [CrossRef]
- Han, Y.J.; Li, L.X.; Dong, M.F.; Yuan, W.J.; Shang, F.D. cDNA cloning of the phytoene synthase (PSY) and expression analysis of PSY and carotenoid cleavage dioxygenase genes in Osmanthus fragrans. Biologia 2013, 68, 258–263. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, P.; Lin, W.; Zheng, X.; Cai, M.; Peng, C. Sequencing of anthocyanin synthesis-related enzyme genes and screening of reference genes in leaves of four dominant subtropical forest tree species. Gene 2019, 716, 144024. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, B.; Liu, J.; Guo, D.; Hou, J.; Chen, S.; Song, B.; Xie, C. Analysis of structural genes and key transcription factors related to anthocyanin biosynthesis in potato tubers. Sci. Hortic. 2017, 225, 310–316. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Li, J.; Ding, Y.; Tian, J.L.; Wang, Z.; Xiong, B.; Xu, T.; Kou, G.; Zheng, Y.; et al. Analysis of anthocyanin accumulation and related gene expression during fig fruit development. Plant Mol. Biol. Rep. 2023. [Google Scholar] [CrossRef]
- Zhou, C.; Mei, X.; Rothenberg, D.O.N.; Yang, Z.; Zhang, W.; Wan, S.; Yang, H.; Zhang, L. Metabolome and transcriptome analysis reveals putative genes involved in anthocyanin accumulation and coloration in white and pink tea (Camellia sinensis) flower. Molecules 2020, 25, 190. [Google Scholar] [CrossRef]
- Zhang, S.M.; Tu, J.L.; Li, C.W.; Hu, Q.; Chen, L.; Dong, L.L. Cloning and expression analysis of chalcone isomerase OfCHI from Osmanthus fragrans. Acta Bot. Bor-Occid. Sin. 2016, 36, 1728–1734. [Google Scholar]
- Liu, Y.C.; Dong, B.; Zhang, C.; Yang, L.Y.; Wang, Y.G.; Zhao, H.B. Effects of exogenous abscisic acid (ABA) on carotenoids and petal color in Osmanthus fragrans ‘Yanhonggui’. Plants 2020, 9, 454. [Google Scholar] [CrossRef]
- Han, Y.; Chen, W.; Yang, F.; Wang, X.; Dong, M.; Zhou, P.; Shang, F. cDNA-AFLP analysis on 2 Osmanthus fragrans cultivars with different flower color and molecular characteristics of OfMYB1 gene. Trees-Struct. Funct. 2015, 29, 931–940. [Google Scholar] [CrossRef]
- Zeng, X.; Liu, C.; Zheng, R.; Cai, X.; Luo, J.; Zou, J.; Wang, C. Emission and accumulation of monoterpene and the key terpene synthase (TPS) associated with monoterpene biosynthesis in Osmanthus fragrans Lour. Front. Plant Sci. 2016, 6, 1232. [Google Scholar] [CrossRef] [PubMed]
- Baldermann, S.; Kato, M.; Fleischmann, P.; Watanabe, N. Biosynthesis of alpha- and beta-ionone, prominent scent compounds, in flowers of Osmanthus fragrans. Acta Biochim. Pol. 2012, 59, 79–81. [Google Scholar] [CrossRef] [PubMed]
- Baldermann, S.; Kato, M.; Kurosawa, M.; Kurobayashi, Y.; Fujita, A.; Fleischmann, P.; Watanabe, N. Functional characterization of a carotenoid cleavage dioxygenase 1 and its relation to the carotenoid accumulation and volatile emission during the floral development of Osmanthus fragrans Lour. J. Exp. Bot. 2010, 61, 2967–2977. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.C.; Molnar, P.; Schwab, W. Cloning and functional characterization of carotenoid cleavage dioxygenase 4 genes. J. Exp. Bot. 2009, 60, 3011–3022. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Pei, J.; Zhao, L.; Tang, F.; Fang, X.; Xie, J. Overexpression and characterization of CCD4 from Osmanthus fragrans and beta-ionone biosynthesis from beta-carotene in vitro. J. Mol. Catal. B-Enzym. 2016, 134, 105–114. [Google Scholar] [CrossRef]
- Zheng, R.; Zhu, Z.; Wang, Y.; Hu, S.; Xi, W.; Xiao, W.; Qu, X.; Zhong, L.; Fu, Q.; Wang, C. UGT85A84 Catalyzes the glycosylation of aromatic monoterpenes in Osmanthus fragrans Lour. Flowers. Front. Plant Sci. 2019, 10, 1376. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, Z.; Chen, R.; He, H.; Ma, J.; Zhang, D. Gene cloning and gene expression characteristics of alcohol dehydrogenase in Osmanthus fragrans var. Semperflorens. Emir. J. Food Agr. 2018, 30, 820–827. [Google Scholar] [CrossRef]
- Chen, X.; Yang, X.; Xie, J.; Ding, W.; Li, Y.; Yue, Y.; Wang, L. Biochemical and comparative transcriptome analyses reveal key genes involved in major metabolic regulation related to colored leaf formation in Osmanthus fragrans ‘Yinbi Shuanghui’ during development. Biomolecules 2020, 10, 549. [Google Scholar] [CrossRef]
- Li, T.; Deng, Y.J.; Liu, J.X.; Duan, A.Q.; Liu, H.; Xiong, A.S. DcCCD4 catalyzes the degradation of alpha-carotene and beta-carotene to affect carotenoid accumulation and taproot color in carrot. Plant J. 2021, 108, 1116–1130. [Google Scholar] [CrossRef]
- Cai, X.; Mai, R.Z.; Zou, J.J.; Zhang, H.Y.; Zeng, X.L.; Zheng, R.R.; Wang, C.Y. Analysis of aroma-active compounds in three sweet osmanthus (Osmanthus fragrans) cultivars by GC-olfactometry and GC-MS. J. Zhejiang Univ.-SCI. B 2014, 15, 638–648. [Google Scholar] [CrossRef]
- Sheng, X.J.; Lin, Y.N.; Cao, J.M.; Ning, Y.; Pang, X.L.; Wu, J.H.; Kong, F.Y. Comparative evaluation of key aroma-active compounds in sweet osmanthus (Osmanthus fragrans Lour.) with different enzymatic treatments. J. Agr. Food Chem. 2021, 69, 332–344. [Google Scholar] [CrossRef]
- Chen, P.Z. Optimization of extraction technology and analysis of components of Osmanthus fragrans essential oil. Cereals Oils 2016, 29, 4. [Google Scholar]
- Orlova, I.; Marshall-Colon, A.; Schnepp, J.; Wood, B.; Varbanova, M.; Fridman, E.; Blakeslee, J.J.; Peer, W.A.; Murphy, A.S.; Rhodes, D.; et al. Reduction of benzenoid synthesis in petunia flowers reveals multiple pathways to benzoic acid and enhancement in auxin transport. Plant Cell. 2006, 18, 3458–3475. [Google Scholar] [CrossRef] [PubMed]
- Henry, L.K.; Gutensohn, M.; Thomas, S.T.; Noel, J.P.; Dudareva, N. Orthologs of the archaeal isopentenyl phosphate kinase regulate terpenoid production in plants. Proc. Natl. Acad. Sci. USA 2015, 112, 10050–10055. [Google Scholar] [CrossRef] [PubMed]
- Hudak, K.A.; Thompson, J.E. Subcellular localization of secondary lipid metabolites including fragrance volatiles in carnation petals. Plant Physiol. 1997, 114, 705–713. [Google Scholar] [CrossRef]
- Xu, C.; Yang, X.L.; Huang, S.C.; Chen, H.; Zhang, Y.B. Cloning and expression analysis of OfCCD1 and OfCCD4 genes in sweet osmanthus. Pak. J. Bot. 2023, 55, 563–569. [Google Scholar] [CrossRef]
- Sun, P.; Chen, X.Y.; Chantarasuwan, B.; Zhu, X.Y.; Deng, X.X.; Bao, Y.; Yu, H. Composition diversity and expression specificity of the TPS gene family among 24 Ficus species. Diversity 2022, 14, 721. [Google Scholar] [CrossRef]
- Ouyang, Q.X.; Ding, W.J.; Wu, X.Y.; Yue, Y.Z.; Yang, X.L.; Wang, L.G. Cloning and expression characteristic analysis of RAP2-12 in Osmanthus fragrans. Acta Bot. Bor-Occid. Sin. 2020, 40, 10. [Google Scholar]
- Liu, Y.C.; Zhang, C.; Dong, B.; Zhao, H.B. Advances of CCD subfamily in higher plants. J. Agric. Biotech. 2019, 027, 720–734. [Google Scholar]
- Li, H.Y.; Yue, Y.Z.; Ding, W.J.; Chen, G.W.; Li, L.; Li, Y.L.; Shi, T.T.; Yang, X.L.; Wang, L.G. Genome-wide identification, classification, and expression profiling reveals R2R3-MYB transcription factors related to monoterpenoid biosynthesis in Osmanthus fragrans. Genes 2020, 11, 353. [Google Scholar] [CrossRef]
- Yan, X.; Ding, W.J.; Wu, X.Y.; Wang, L.G.; Yang, X.L.; Yue, Y.Z. Insights into the MYB-Related transcription factors involved in regulating floral aroma synthesis in sweet osmanthus. Front. Plant Sci. 2022, 13, 14. [Google Scholar] [CrossRef]
- Yeon, J.Y.; Kim, W.S. Biosynthetic linkage between the color and scent of flowers: A review. Hortic. Sci. Technol. 2021, 39, 697–713. [Google Scholar] [CrossRef]
- Wang, S.G.; Feng, X.; Li, G.C. Research on the development environment of the extraction enterprises of sweet osmanthus essential oil. Ekoloji 2018, 27, 163–171. [Google Scholar]
- Pan, C.; Zhao, L.; Zhao, D. Microwave-assisted green extraction of antioxidant components from Osmanthus fragrans (Lour) flower using natural deep eutectic solvents. J. Appl. Res. Med. Aroma 2021, 20, 100285. [Google Scholar] [CrossRef]
- Usuki, T.; Munakata, K. Extraction of essential oils from the flowers of Osmanthus fragrans var. aurantiacus using an ionic liquid. Bull. Chem. Soc. Jpn. 2017, 90, 1105–1110. [Google Scholar] [CrossRef]
- Wang, L.; Tan, N.N.; Hu, J.Y.; Wang, H.; Duan, D.Z.; Ma, L.; Xiao, J.; Wang, X.L. Analysis of the main active ingredients and bioactivities of essential oil from Osmanthus fragrans var. thunbergii using a complex network approach. BMC Syst. Biol. 2017, 11, 12. [Google Scholar] [CrossRef]
- Chen, G.L.; Chen, S.G.; Xiao, Y.; Fu, N.L. Antioxidant capacities and total phenolic contents of 30 flowers. Ind. Crop. Prod. 2018, 111, 430–445. [Google Scholar] [CrossRef]
- Wu, L.P.; Liu, J.Y.; Huang, W.S.; Wang, Y.X.; Chen, Q.; Lu, B.Y. Exploration of Osmanthus fragrans Lour.’s composition, nutraceutical functions and applications. Food Chem. 2022, 377, 21. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.Z.; Xu, Y.Y.; Zhou, Z.C. Management Systems of Flower-Themed Tourism in China: A Value Chain Analysis. In Proceedings of the 18th International Symposium on Horticultural Economics and Management, Alnarp, Sweden, 31 May–3 June 2015. [Google Scholar]
- Chen, M.X.; Zhang, K.L.; Gao, B.; Yang, J.F.; Tian, Y.; Das, D.; Fan, T.; Dai, L.; Hao, G.F.; Yang, G.F.; et al. Phylogenetic comparison of 5′ splice site determination in central spliceosomal proteins of the U1-70K gene family, in response to developmental cues and stress conditions. Plant J. 2020, 103, 357–378. [Google Scholar] [CrossRef]
- Song, T.; Das, D.; Ye, N.H.; Wang, G.Q.; Zhu, F.Y.; Chen, M.X.; Yang, F.; Zhang, J.H. Comparative transcriptome analysis of coleorhiza development in japonica and Indica rice. BMC Plant Biol. 2021, 21, 514. [Google Scholar] [CrossRef]
- Zhao, Y.; Xin, D.; Lu, W.; Zong, X.; Niu, Y.; Guo, X.; Ma, Y.; Qiang, W.; Su, H.; Zhang, S.; et al. PeMPK7 is induced in an ROS-dependent manner and confers poplar para-hydroxybenzoic acid stress resistance through the removal of ROS. Ind. Crop. Prod. 2022, 182, 114861. [Google Scholar] [CrossRef]
- Zou, J.J.; Gao, W.; Cai, X.; Zeng, X.L.; Wang, C.Y. Somatic embryogenesis and plant regeneration in Osmanthus fragrans Lour. Propag. Ornam. Plants 2014, 14, 32–39. [Google Scholar]
- Zhong, S.; Dong, B.; Zhou, J.; Miao, Y.; Yang, L.; Wang, Y.; Xiao, Z.; Fang, Q.; Wan, Q.; Zhao, H. Highly efficient transient gene expression of three tissues in Osmanthus fragrans mediated by Agrobacterium tumefaciens. Sci. Hortic. 2023, 310, 111725. [Google Scholar] [CrossRef]
- Gu, H.; Ding, W.; Shi, T.; Ouyang, Q.; Yang, X.; Yue, Y.; Wang, L. Integrated transcriptome and endogenous hormone analysis provides new insights into callus proliferation in Osmanthus fragrans. Sci. Rep. 2022, 12, 7609. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.C.; Chen, M.X.; Xiao, T.; Zhang, C.; Shang, J.; Zhang, K.L.; Zhu, F.Y. Global proteome response to Pb(II) toxicity in poplar using SWATH-MS-based quantitative proteomics investigation. Ecotox. Environ. Safe. 2021, 220, 8. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | Structure | Function |
|---|---|---|
| α-carotene |  | Antioxidants destroy free radicals and prevent chronic diseases such as heart disease and cancer. |
| β-carotene |  | Reduces lipid peroxidation, scavenges free radicals, and prevents aging and disease in the body. |
| anthocyanins |  | Flavonoids, anti-staphylococcus aureus and hemolytic streptococcus, anti-aging and hypoxic, anti-adverse irritant, anti-fatigue, inhibition of blood sugar elevation. |
| chalcone |  | Flavonoids, one of the largest groups of bioactive natural products, anticancer, anti-inflammatory, antibacterial, antioxidant and anti-parasitic. |
| quercetin |  | Flavonoids, with a variety of biological activities, are antioxidants and play an important role in the clinical treatment of cancer and cardiovascular diseases. |
| rutin |  | Anti-inflammatory, maintains vascular resistance, promotes cell proliferation, prevents blood cell coagulation, diuretic, cough suppressant, hypolipidemic, hypotensive, protects ulcer surface. |
| verbascoside |  | Pharmacological effects include immune modulation, antioxidant, energy enhancement and anti-fatigue. |
| genistin |  | Improve insulin resistance, anti-diabetic effect, inhibit cancer cell activity, inhibit prostate cancer and antioxidant activity. |
| kaempferol |  | Anticancer, fertility inhibitor, anti-epileptic, anti-inflammatory, antioxidant, antispasmodic, anti-ulcer, choleretic diuretic, cough suppressant. |
| isorhamnetin |  | Various biological effects include antitumor, anti-myocardial hypoxia and ischemia, relieving angina pectoris, anti-arrhythmia, treating coronary heart disease and hypertension, scavenging oxygen free radicals, lowering serum cholesterol, etc. It is widely used in clinical practice. |
| naringin |  | Naringin can significantly improve local microcirculation and nutrient supply, inhibit the proliferation and migration of melanocytes, and effectively fight against dark circles caused by hyperpigmentation. |
| camellianin A |  | Flavonoids, anticancer activity and anti-angiotensin-converting enzyme (ACE) activity. |
| cyanidin chloride |  | Strong antioxidant, anti-inflammatory, anti-allergic, antimicrobial, anticancer, vision protection and cardiovascular disease prevention. |
| paeoniflorin |  | Hepatoprotective effect; immune modulating effect and anti-hyperglycemic effect, inhibits nerve cell damage and improves motor retardation, can be used to treat Alzheimer’s disease, Parkinson’s disease, epilepsy and other brain disorders. |
| ocimene |  | Ocimene is used in the production of spices and in daily chemical flavors. |
| linalool |  | Important spice, a raw material for the blending of various artificial essential oils, used in the formulation of cosmetics, soap, detergent, food and other flavors |
| β-ionone |  | Used in daily cosmetics, beauty products, food flavors, and in large quantities for the production of vitamin A, E and carotene |
| α-ionone |  | Permission to use edible spice temporarily by GB 2760-96, mainly used to formulate longan, raspberry, blackberry, cherry, citrus and other types of spices. |
| phenylalanine |  | Amino acid derivative, used as a spice for food. |
| shikimic acid |  | Inhibit platelet aggregation, inhibit arterial and venous thrombosis and cerebral thrombosis, have anti-inflammatory and analgesic effects, can also be used as antiviral and anticancer drug intermediates. |
| trans-cinnamic acid |  | With fragrance, sterilization, antiseptic, preservation, can be used in the pharmaceutical industry, beauty, pesticide industry, organic synthesis chemical. |
| Genes | Description | Gene_ID | Homeosis Gene | Function | Refs |
|---|---|---|---|---|---|
| PSY | phytoene synthase | ofr.gene9262 | AT5G17230 | Color | [44] |
| Z-ISO | ζ-carotene isomerase | ofr.gene50192 | AT1G10830 | Color | [50] |
| CRTISO | carotenoid isomerase | ofr.gene51545 | AT1G06820 | Color | [50] |
| PAL | phenylalanine ammonia-lyase | ofr.gene722 | AT3G10340 | Color | [51] |
| CHI | chalcone isomerase | ofr.gene57827 | AT3G55120 | Color | [51] |
| TPS | Terpene Synthase | ofr.gene51840 | AT4G20230 | Fragrance | [52] |
| CCDs | carotenoid cleavage dioxygenase | ofr.gene58593 | AT4G19170 | Fragrance &color | [21,43,44,53,54,55,56] |
| UGT | UDP-glucosyl transferase | ofr.gene54750 | AT2G36780 | Fragrance | [57] |
| ADH | Alcohol dehydrogenase | ofr.gene58337 | AT3G15880 | Fragrance | [58] |
| MECS | 2-C-methyl-D-erythritol 2,4-cyclodiphosphate synthase | ofr.gene9262 | AT1G63970 | Fragrance | [39] |
| HDR | 4-hydroxy-3-methylbut-2-enyl diphosphate reductase | ofr.gene5420 | AT4G34350 | Fragrance | [39] |
| DXS | 1-Deoxy-D-xylulose-5-phosphate synthase | ofr.gene30635 | AT4G15560 | Fragrance | [59] |
| GGPPS | Geranylgeranyl pyrophosphate synthase | ofr.gene53350 | AT4G36810 | Fragrance | [44] |
| NCED1 | 9-Cis-epoxycarotenoid dioxygenases | ofr.gene29722 | AT3G63520 | Fragrance &color | [60] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, Y.; Shan, L.; Zhao, R.; Tang, J.; Zhang, C.; Chen, M.; Duan, Y.; Zhu, F. Recent Advances in Flower Color and Fragrance of Osmanthus fragrans. Forests 2023, 14, 1403. https://doi.org/10.3390/f14071403
Qian Y, Shan L, Zhao R, Tang J, Zhang C, Chen M, Duan Y, Zhu F. Recent Advances in Flower Color and Fragrance of Osmanthus fragrans. Forests. 2023; 14(7):1403. https://doi.org/10.3390/f14071403
Chicago/Turabian StyleQian, Yuchen, Linmiao Shan, Rong Zhao, Jinwu Tang, Cheng Zhang, Moxian Chen, Yifan Duan, and Fuyuan Zhu. 2023. "Recent Advances in Flower Color and Fragrance of Osmanthus fragrans" Forests 14, no. 7: 1403. https://doi.org/10.3390/f14071403
APA StyleQian, Y., Shan, L., Zhao, R., Tang, J., Zhang, C., Chen, M., Duan, Y., & Zhu, F. (2023). Recent Advances in Flower Color and Fragrance of Osmanthus fragrans. Forests, 14(7), 1403. https://doi.org/10.3390/f14071403