

Top-Leader Growth in Nordmann Fir (Abies nordmanniana)
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Climate Data
2.3. Plant Material
2.4. Measurements
2.5. Statistical Data Analysis
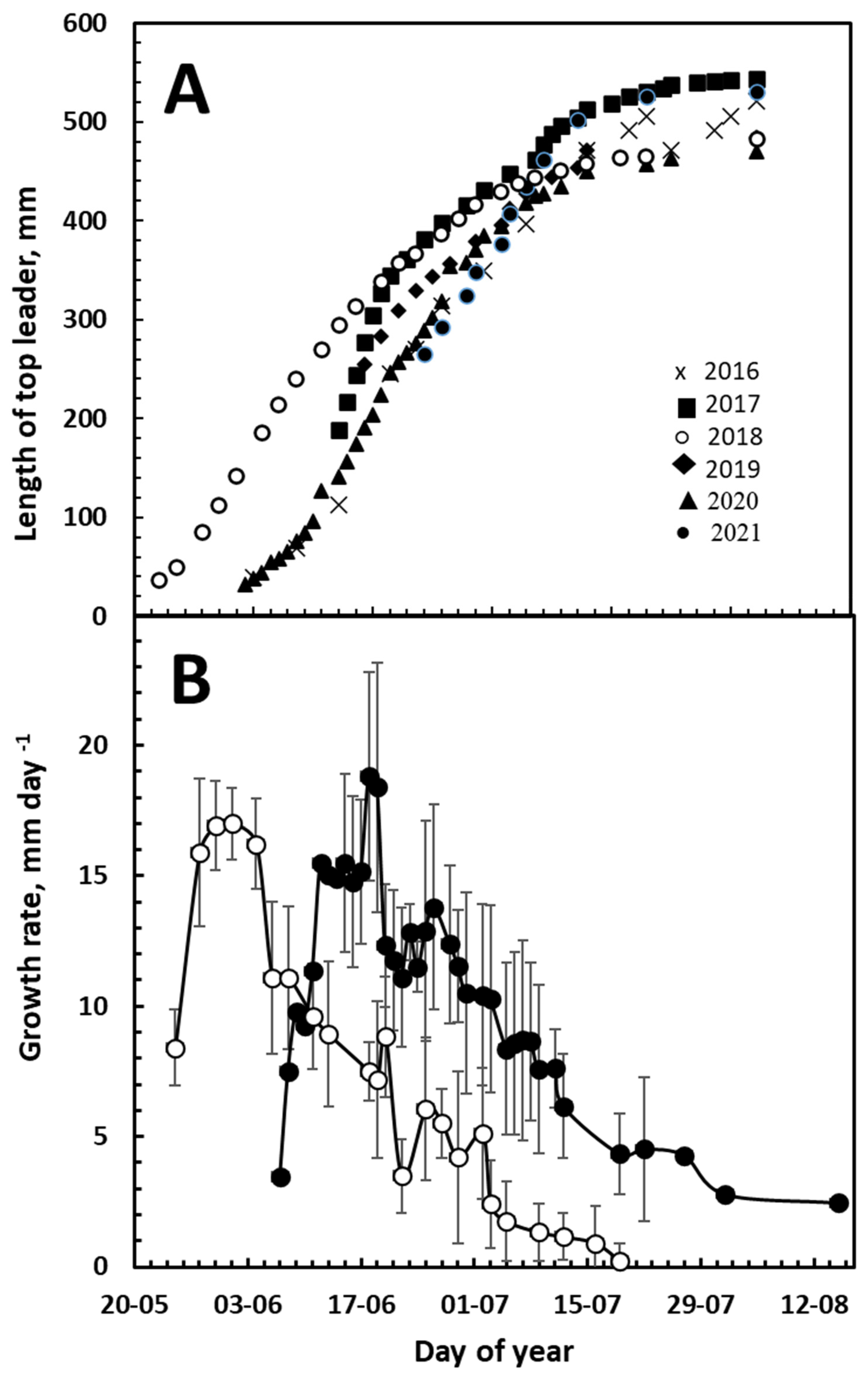
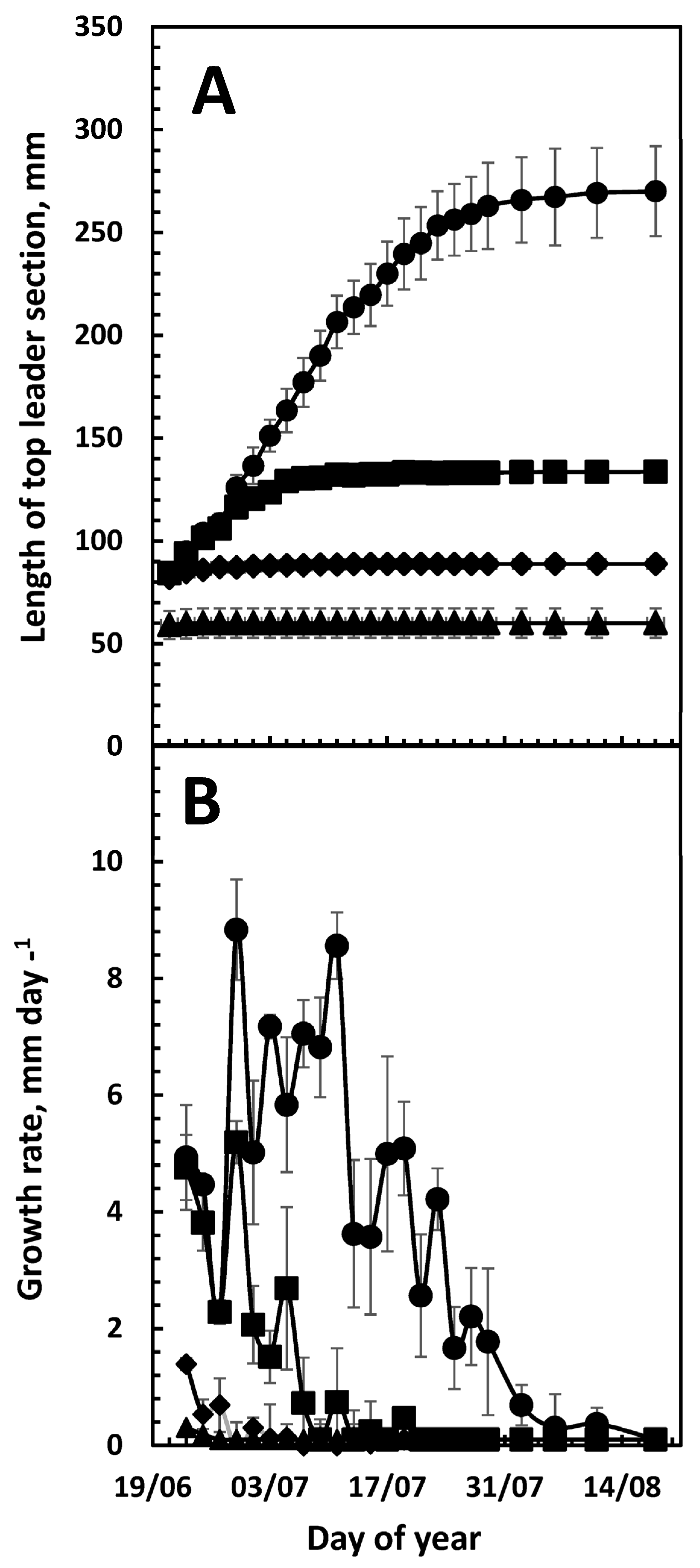
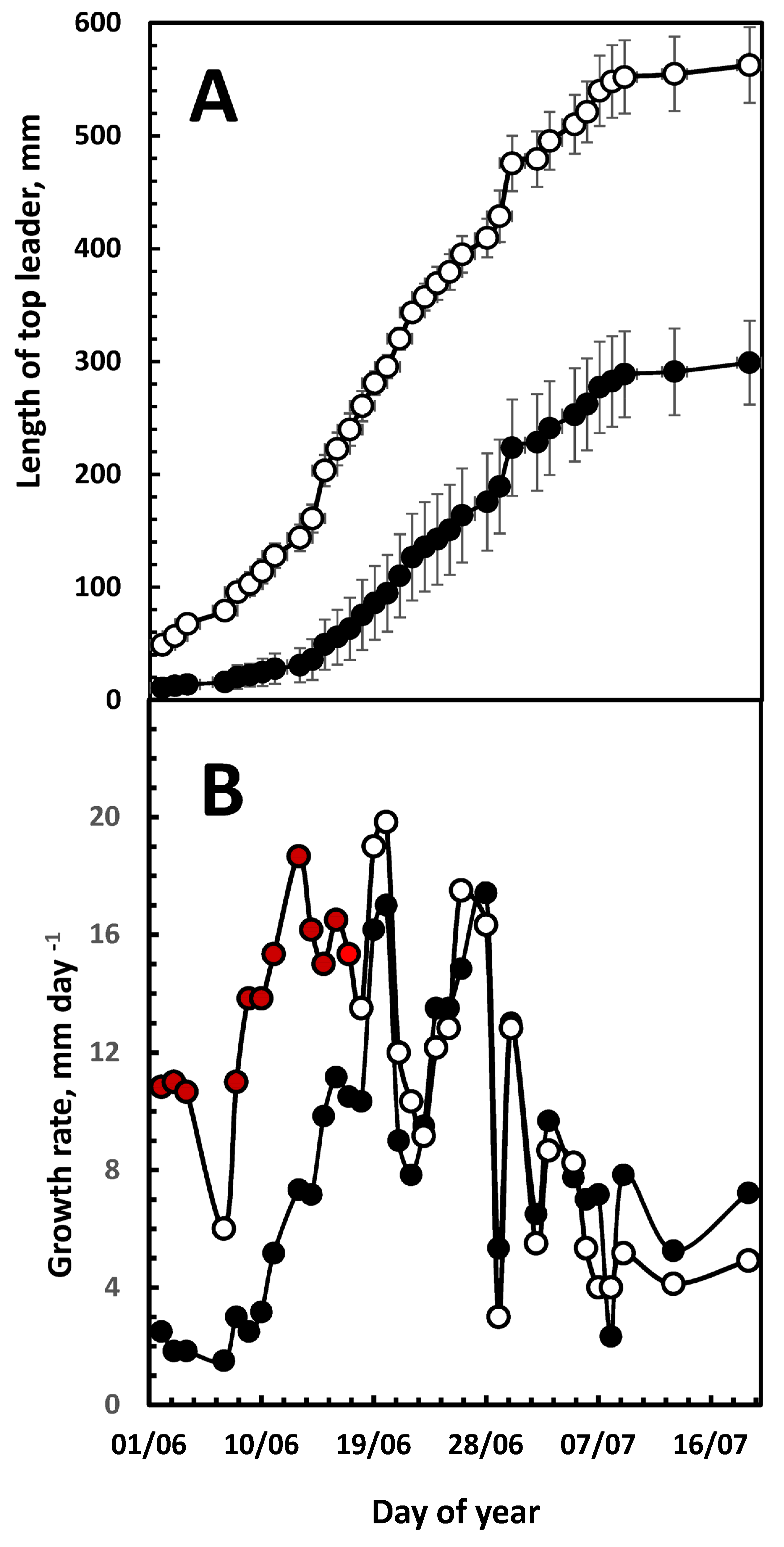
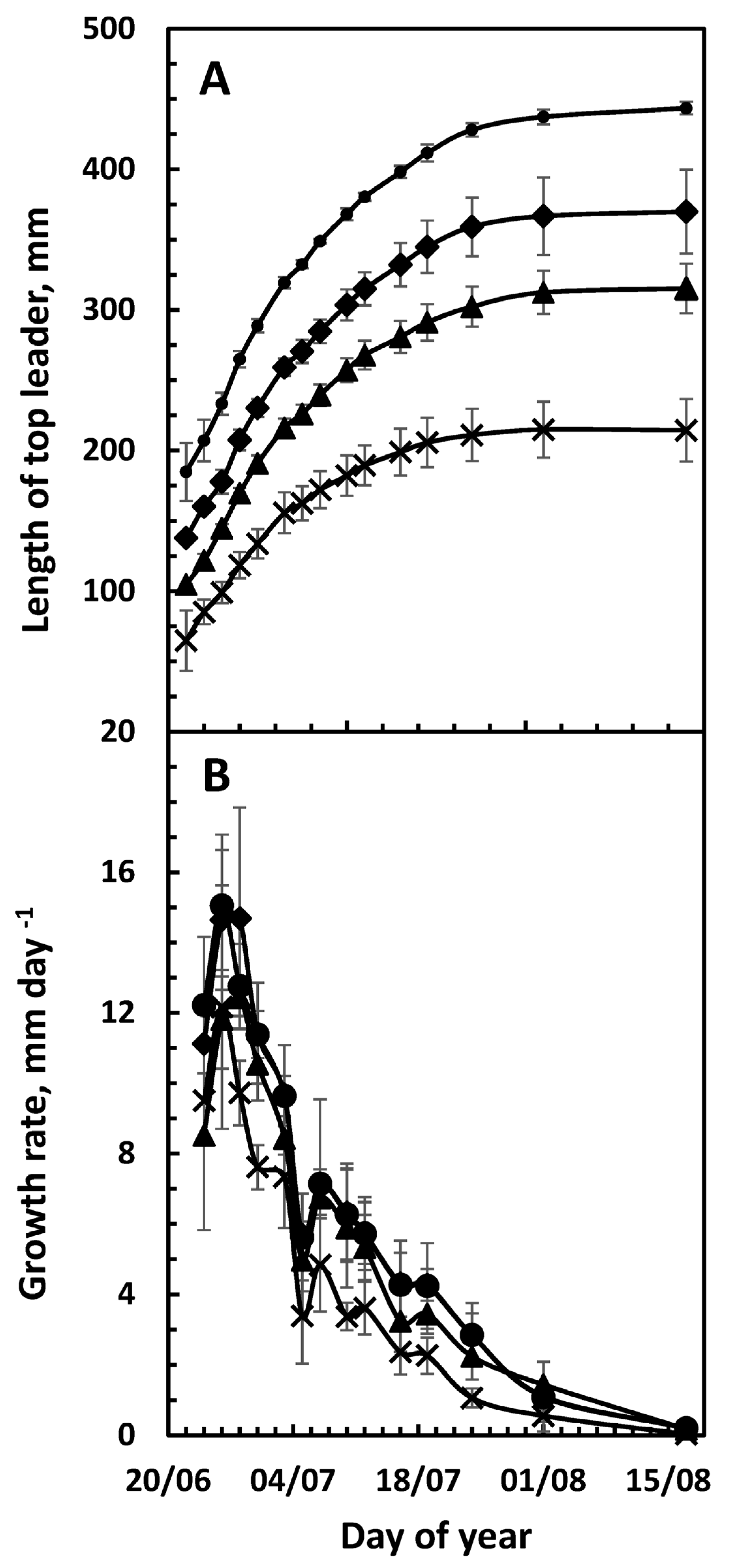
3. Results
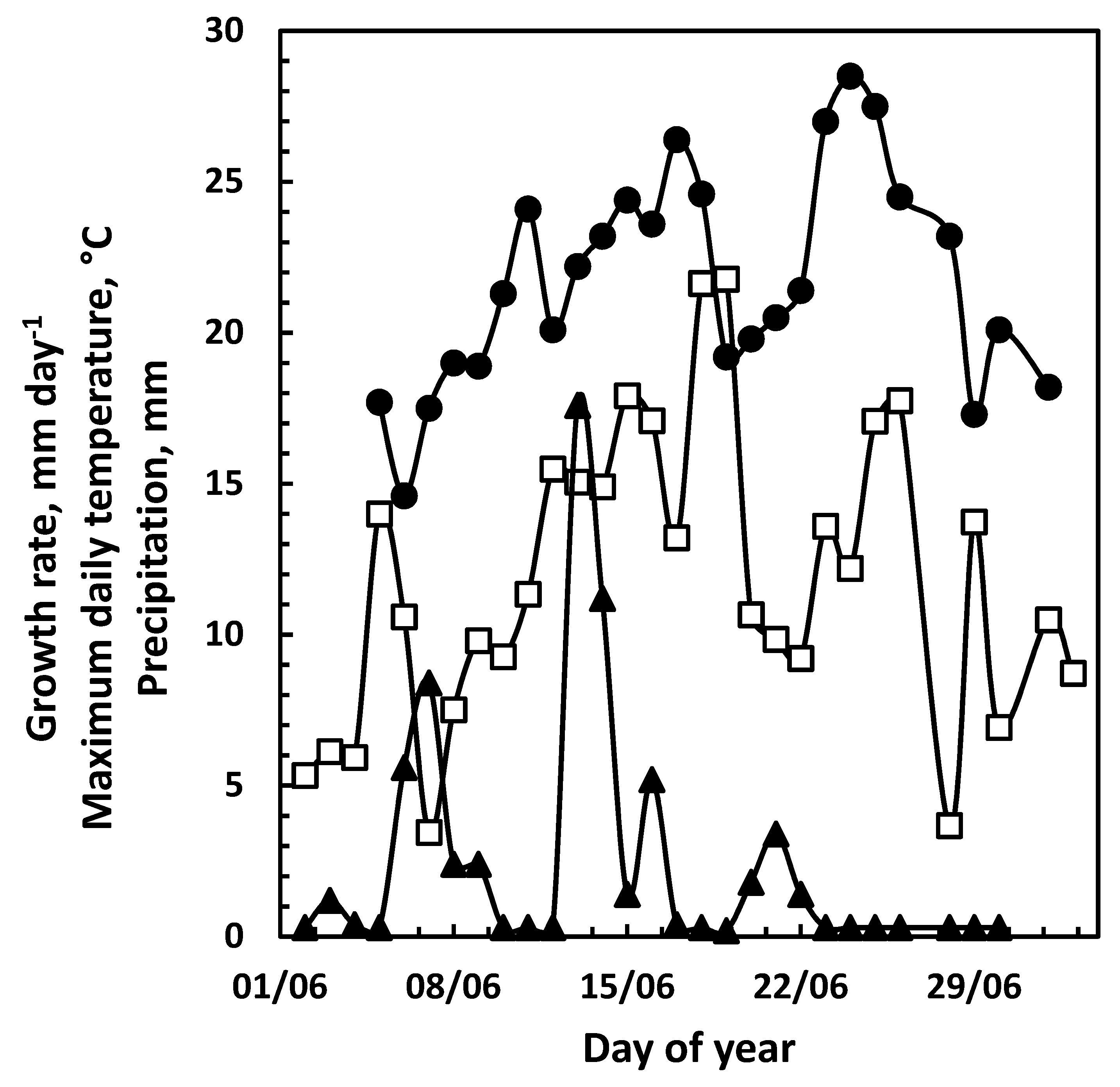
3.1. Top-Leader Elongation
3.2. Daily Growth Rates
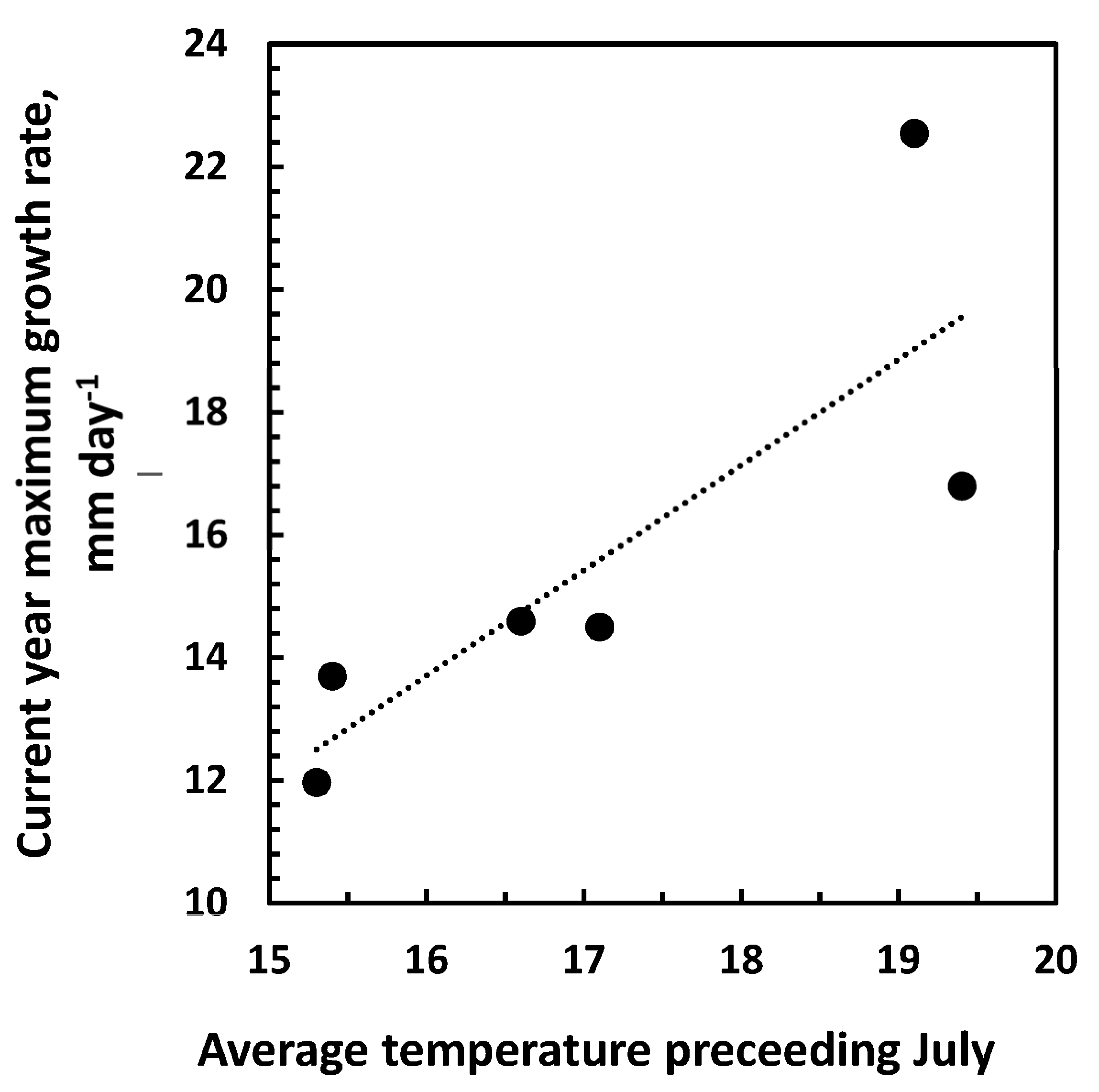
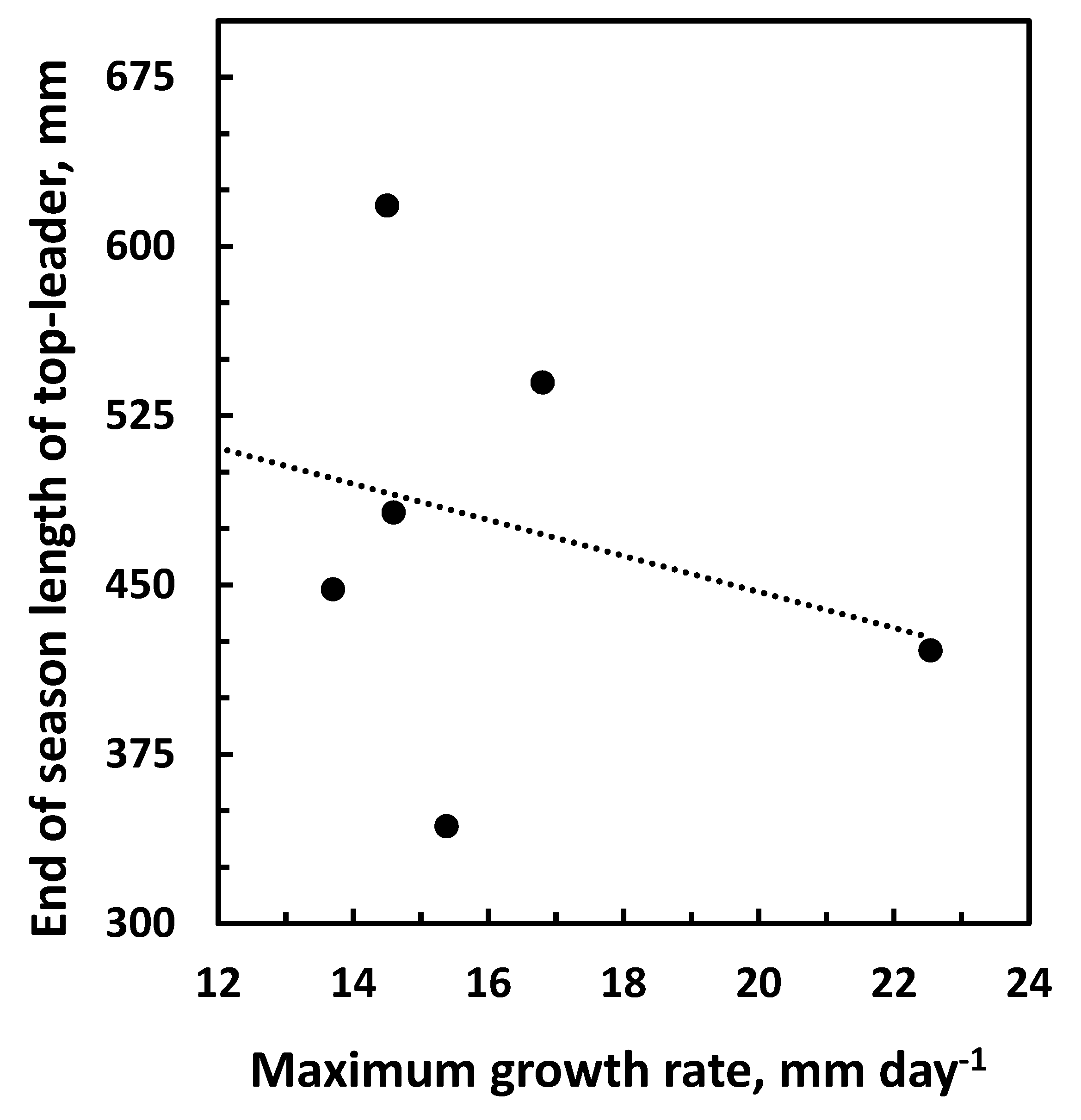
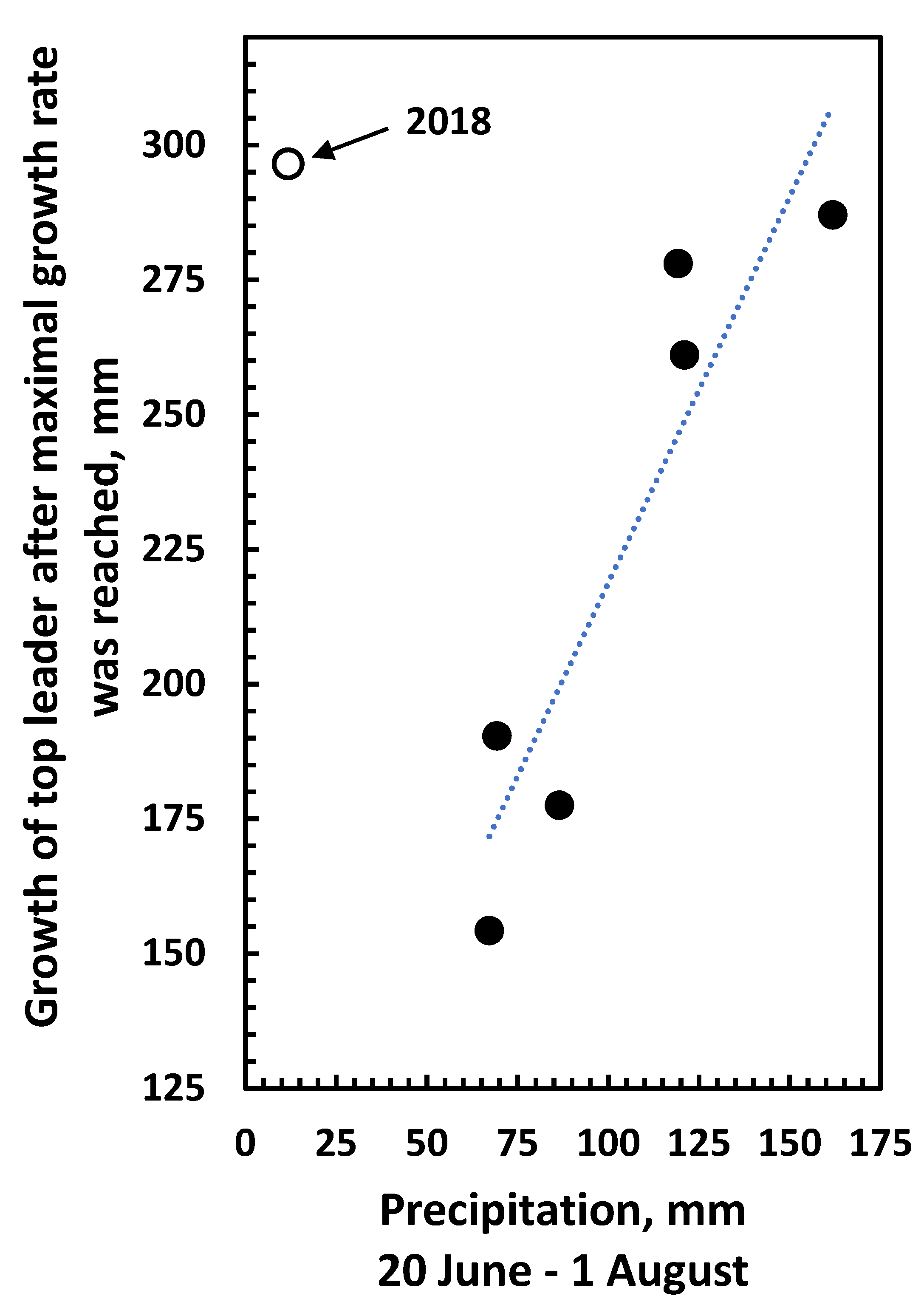
3.3. Growth and Climatic Correlations
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Nielsen, U.B.; Hansen, J.K.; Kromann, H.K. Impact of site and provenance on economic return in Nordmann fir Christmas tree production. Scand. J. For. Res. 2011, 26, 74–89. [Google Scholar] [CrossRef]
- Nielsen, U.B.; Xu, J.; Hansen, O.K. Genetics in and opportunities for improvement of Nordmann fir (Abies nordmanniana(Steven) Spach) Christmas tree production. Tree Genet. Genomes 2020, 16, 66. [Google Scholar] [CrossRef]
- Chen, H.J.; Bollmark, M.; Eliasson, L. Evidence that cytokinin controls bud size and branch form in Norway spruce. Physiol. Plant. 1996, 98, 612–618. [Google Scholar] [CrossRef]
- Hejnowicz, A.; Obarska, E. Structure and development of vegetative buds, from the lower crown of picea-abies. Ann. Des Sci. For. 1995, 52, 433–447. [Google Scholar] [CrossRef]
- Baxter, S.M.; Cannell, M.G.R. Branch Development on Leaders of Picea-Sitchensis. Can. J. For. Res. 1978, 8, 121–128. [Google Scholar] [CrossRef]
- Sterling, C. Organization of the shoot of pseudotsuga-taxifolia (lamb) britt. I. Structure of the shoot apex. Am. J. Bot. 1946, 33, 742–750. [Google Scholar] [CrossRef]
- Kutschera, U.; Niklas, K.J. Cell division and turgor-driven stem elongation in juvenile plants: A synthesis. Plant Sci. 2013, 207, 45–56. [Google Scholar] [CrossRef]
- Brown, C.L.; Sommer, H.E. Shoot growth and histogenesis of trees possessing diverse patterns of shoot development. Am. J. Bot. 1992, 79, 335–346. [Google Scholar] [CrossRef]
- Ripetti, V.; Escoute, J.; Verdeil, J.L.; Costes, E. Shaping the shoot: The relative contribution of cell number and cell shape to variations in internode length between parent and hybrid apple trees. J. Exp. Bot. 2008, 59, 1399–1407. [Google Scholar] [CrossRef]
- Song, Y.J.; Sass-Klaassen, U.; Sterck, F.; Goudzwaard, L.; Akhmetzyanov, L.; Poorter, L. Growth of 19 conifer species is highly sensitive to winter warming, spring frost and summer drought. Ann. Bot. 2021, 128, 545–557. [Google Scholar] [CrossRef]
- Junttila, O. Effects of temperature on shoot growth in northern provenances of Pinus sylvestris L. Tree Physiol. 1986, 1, 185–192. [Google Scholar] [CrossRef]
- Salminen, H.; Jalkanen, R.; Lindholm, M. Summer temperature affects the ratio of radial and height growth of Scots pine in northern Finland. Ann. For. Sci. 2009, 66, 810. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Ward, R.C. Seasonal hight growth of conifers. For. Sci. 1957, 3, 61–66. [Google Scholar]
- Takahashi, K. Effects of climatic conditions on shoot elongation of alpine dwarf pine (Pinus pumila) at its upper and lower altitudinal limits in central Japan. Arct. Antarct. Alp. Res. 2003, 35, 1–7. [Google Scholar] [CrossRef]
- Tanabe, T.; Epron, D.; Dannoura, M. A new approach to identify the climatic drivers of leaf production reconstructed from the past yearly variation in annual shoot lengths in an evergreen conifer (Picea mariana). Trees-Struct. Funct. 2022, 36, 179–190. [Google Scholar] [CrossRef]
- Wang, Y.F.; Cufar, K.; Eckstein, D.; Liang, E.Y. Variation of Maximum Tree Height and Annual Shoot Growth of Smith Fir at Various Elevations in the Sygera Mountains, Southeastern Tibetan Plateau. PLoS ONE 2012, 7, e31725. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Deslauriers, A.; Gricar, J.; Seo, J.W.; Rathgeber, C.B.K.; Anfodillo, T.; Morin, H.; Levanic, T.; Oven, P.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- Rubio-Cuadrado, A.; Bravo-Oviedo, A.; Mutke, S.; Del Rio, M. Climate effects on growth differ according to height and diameter along the stem in Pinus pinaster Ait. Iforest-Biogeosci. For. 2018, 11, 237–242. [Google Scholar] [CrossRef]
- Rossi, S.; Isabel, N. The timing of bud break in warming conditions: Variation among seven sympatric conifer species from Eastern Canada. Int. J. Biometeorol. 2017, 61, 1983–1991. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Shoot growth in woody plants. Bot. Rev. 1964, 30, 335–392. [Google Scholar] [CrossRef]
- Ravn, J.; D’Orangeville, L.; Lavigne, M.B.; Taylor, A.R. Phenotypic plasticity enables considerable acclimation to heat and drought in a cold-adapted boreal forest tree species. Front. For. Glob. Chang. 2022, 5, 1075787. [Google Scholar] [CrossRef]
- Pedersen, L.B.; Christensen, C.J.; Ingerslev, M. Leaching versus input of nitrogen, potassium and magnesium in different fertilizer regimens in Christmas tree stands of Abies nordmanniana in Denmark. Scand. J. For. Res. 2006, 21, 130–142. [Google Scholar] [CrossRef]
- Martens, H.J.; Sorensen, S.; Burow, M.; Veierskov, B. Characterization of Top Leader Elongation in Nordmann Fir (Abies nordmanniana). J. Plant Growth Regul. 2019, 38, 1354–1361. [Google Scholar] [CrossRef]
- Landgren, C.; Cregg, B.; Rouse, R.; Kowalski, J. Controlling Leader Growth on Noble and Turkish Fir with S-ABA. Forests 2022, 13, 212. [Google Scholar] [CrossRef]
- Shi, P.J.; Chen, L.; Hui, C.; Grissino-Mayer, H.D. Capture the time when plants reach their maximum body size by using the beta sigmoid growth equation. Ecol. Model. 2016, 320, 177–181. [Google Scholar] [CrossRef]
- Veierskov, B.; Persson, D.; Martens, H.; Pedersen, L. Røde nåle og bare skuldre—Hvorfor opstår det? Nåledrys 2019, 107, 4–9. [Google Scholar]
- Rasmussen, H.N.; Veierskov, B.; Hansen-Moller, J.; Norbaek, R. “Lateral Control”: Phytohormone Relations in the Conifer Treetop and the Short- and Long-Term Effects of Bud Excision in Abies nordmanniana. J. Plant Growth Regul. 2010, 29, 268–279. [Google Scholar] [CrossRef]
- Jacqmard, A.; Gadisseur, I.; Bernier, G. Cell division and morphological changes in the shoot apex of Arabidopsis thaliana during floral transition. Ann. Bot. 2003, 91, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, P.B.; Cassell, S.J.; Adams, P.A. On nature of intercalary growth and cellular differentiation in internodes of avena sativa. Bot. Gaz. 1965, 126, 1–13. [Google Scholar] [CrossRef]
- Hayatgheibi, H.; Haapanen, M.; Lundstromer, J.; Berlin, M.; Karkkainen, K.; Helmersson, A. The Impact of Drought Stress on the Height Growth of Young Norway Spruce Full-Sib and Half-Sib Clonal Trials in Sweden and Finland. Forests 2021, 12, 498. [Google Scholar] [CrossRef]
- Salminen, H.; Jalkanen, R. Intra-annual height increment of Pinus sylvestris at high latitudes in Finland. Tree Physiol. 2007, 27, 1347–1353. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kozlowski, T.T.; Torrie, J.H.; Marshall, P.E. Predictability of shoot length from bud size in Pinus-resinosa. Can. J. For. Res. 1973, 3, 34–38. [Google Scholar] [CrossRef]
- Zlobin, I.E. Linking the growth patterns of coniferous species with their performance under climate aridization. Sci. Total Environ. 2022, 831, 154971. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| °C | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | April | May | June | July | August | September | October | November | December | January | February | March |
| 2016 | 6.0 | 12.7 | 15.8 | 16.1 | 15.8 | 15.8 | 8.9 | 3.9 | 4.8 | 0.1 | 2.1 | 3.7 |
| 2017 | 6.2 | 11.9 | 14.5 | 15.3 | 15.7 | 12.8 | 10.7 | 5.0 | 3.3 | 1.2 | 1.9 | 4.7 |
| 2018 | 8.3 | 14.9 | 16.4 | 19.4 | 17.2 | 13.6 | 9.8 | 5.8 | 4.1 | 2.1 | −0.9 | 0.1 |
| 2019 | 7.5 | 9.4 | 15.8 | 16.6 | 16.9 | 13.0 | 9.0 | 5.8 | 4.6 | 1.4 | 4.3 | 5.1 |
| 2020 | 7.8 | 10.3 | 16.9 | 15.4 | 18.5 | 14.1 | 10.2 | 7.5 | 4.2 | 5.0 | 4.5 | 4.4 |
| 2021 | 5.7 | 10.5 | 17.1 | 19.1 | 15.7 | 14.4 | 10.4 | 6.7 | 1.6 | 0.5 | −0.1 | 3.9 |
| 2022 | 6.3 | 11.8 | 15.6 | 17.1 | 18.7 | 13.3 | 11.7 | 7.4 | 1.1 | 3.5 | 3.7 | 3.6 |
| Maximum Growth Rate | Length of Top-Leader on Day of Maximal Growth Rate | |||
|---|---|---|---|---|
| Year | Date | mm Day−1 | mm | As % of Final Length |
| 2016 | 28 June | 12.0 a | 233 a | 45 |
| 2017 | 19 June | 16.8 b | 279 a | 52 |
| 2018 | 4 June | 14.6 a | 210 a,b | 39 |
| 2019 | 20 June | 13.7 a | 258 a | 58 |
| 2020 | 20 June | 22.5 c | 243 a | 58 |
| 2021 | 27 June | 14.5 a | 340 c | 55 |
| 2022 | 28 June | 13.2 a | 204 b | 55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veierskov, B. Top-Leader Growth in Nordmann Fir (Abies nordmanniana). Forests 2023, 14, 1214. https://doi.org/10.3390/f14061214
Veierskov B. Top-Leader Growth in Nordmann Fir (Abies nordmanniana). Forests. 2023; 14(6):1214. https://doi.org/10.3390/f14061214
Chicago/Turabian StyleVeierskov, Bjarke. 2023. "Top-Leader Growth in Nordmann Fir (Abies nordmanniana)" Forests 14, no. 6: 1214. https://doi.org/10.3390/f14061214
APA StyleVeierskov, B. (2023). Top-Leader Growth in Nordmann Fir (Abies nordmanniana). Forests, 14(6), 1214. https://doi.org/10.3390/f14061214