
Impact of Crown Closure on Cone Production and Effective Number of Parents in Natural Stands of Taurus Cedar (Cedrus libani A. Rich.)
Abstract
1. Introduction
2. Materials and Methods
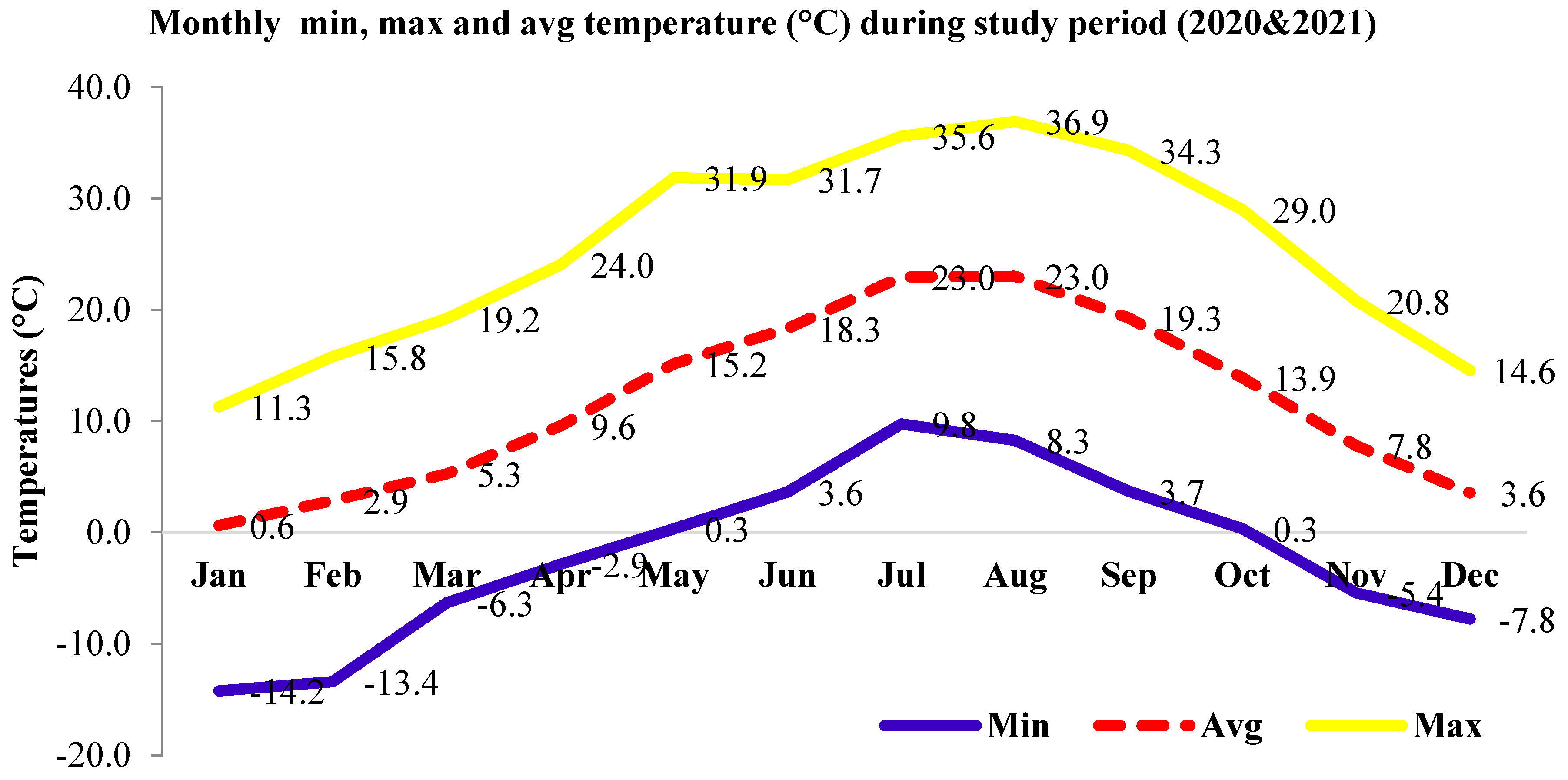
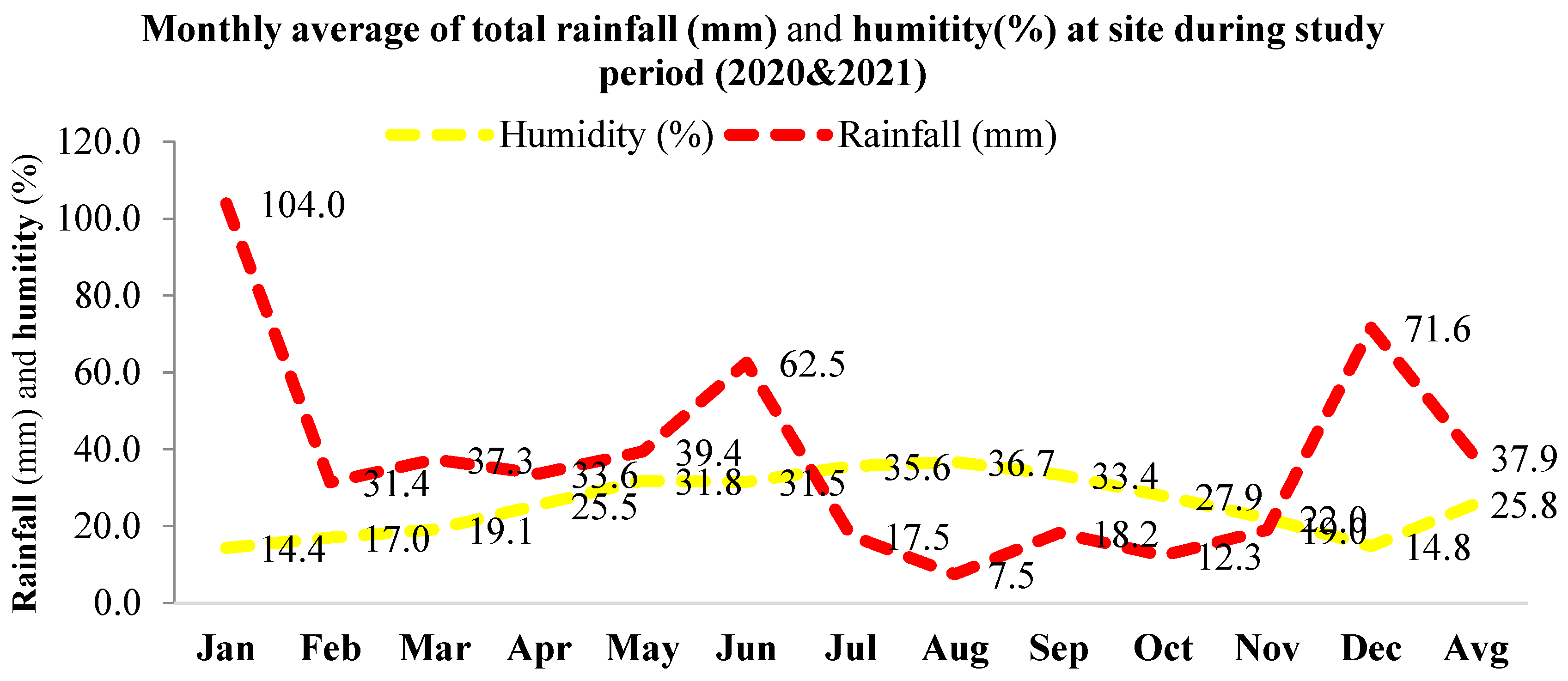
2.1. Study Area
2.2. Data Collection and Analysis
3. Results
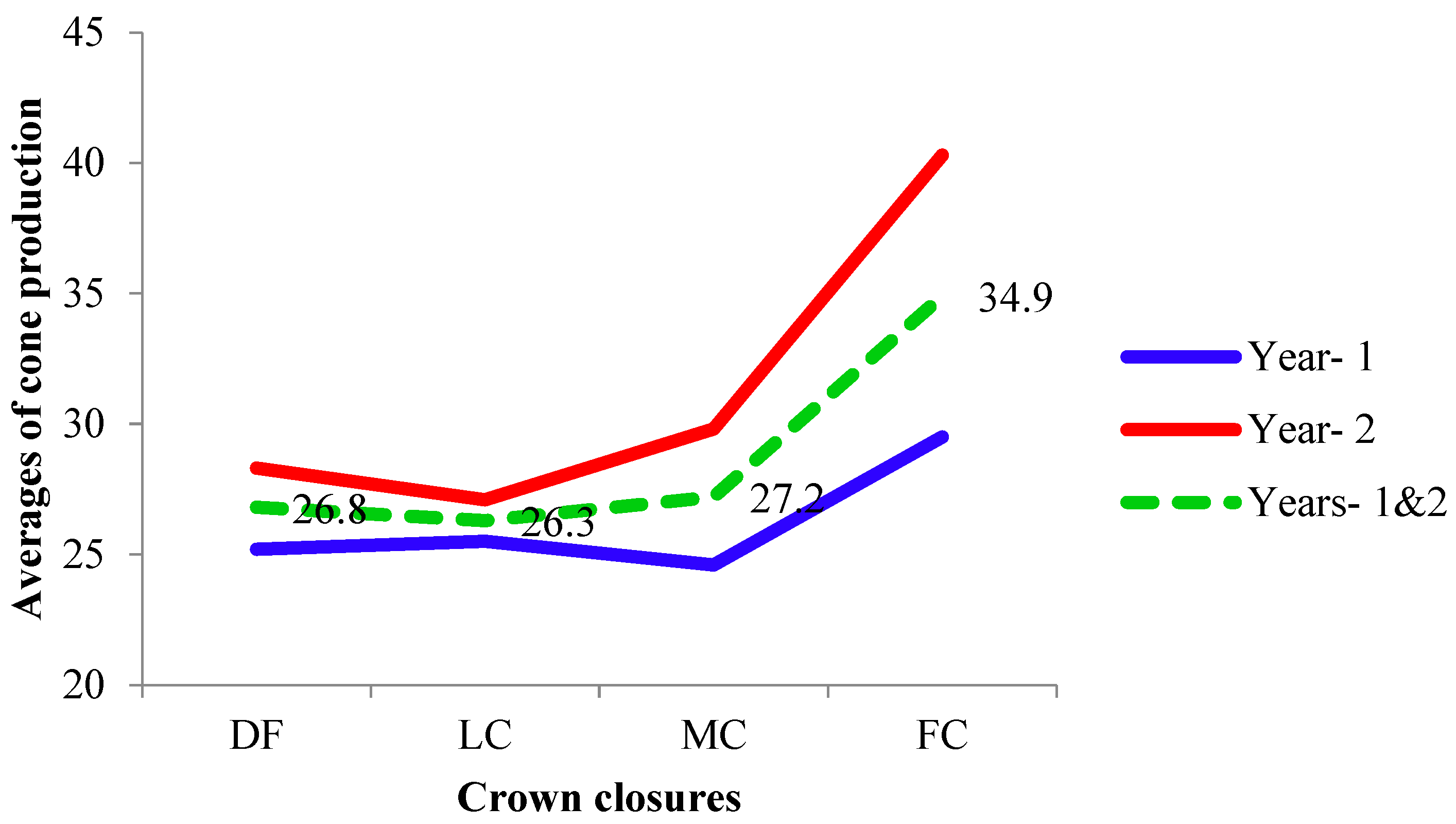
3.1. Cone Production
3.2. Fertility Variation, Effective Number of Parents, and Gene Diversity
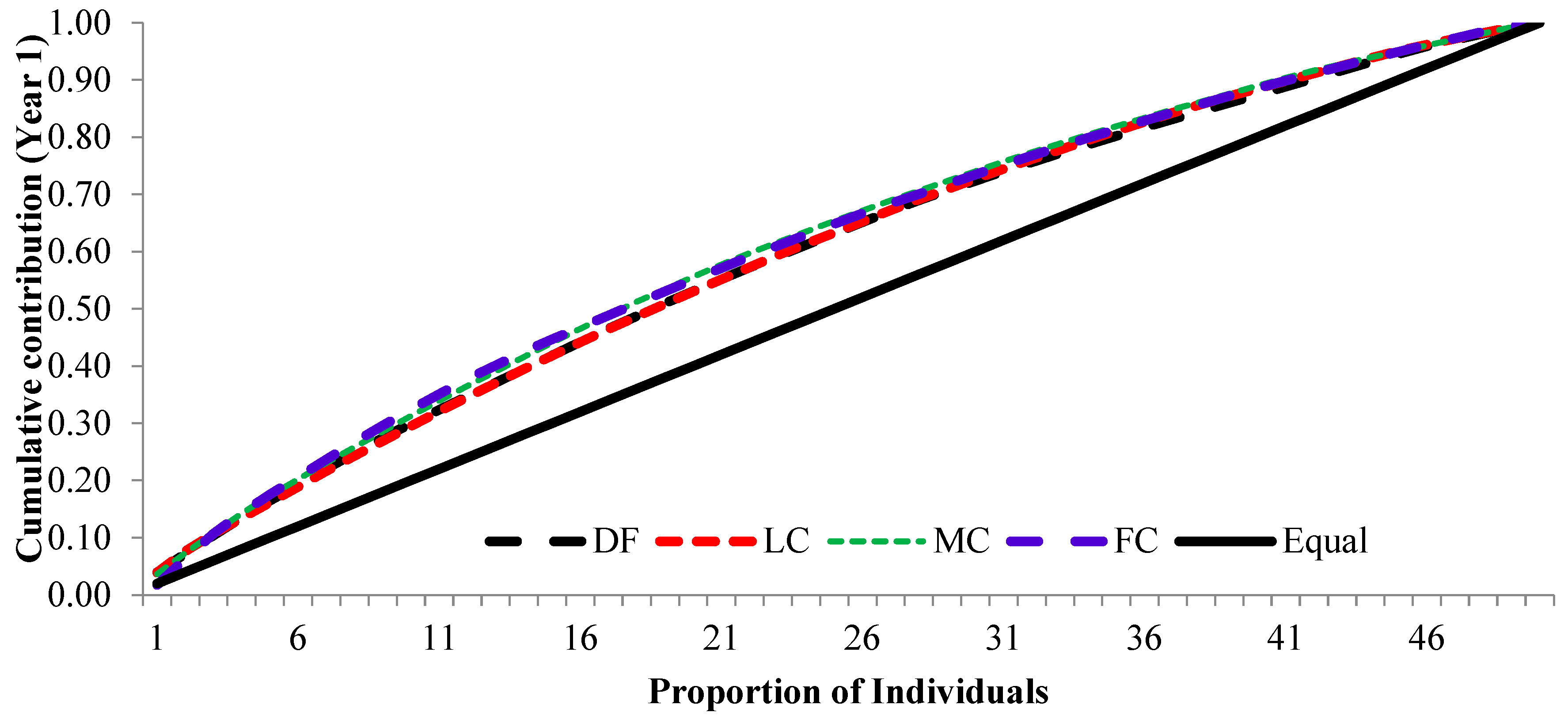
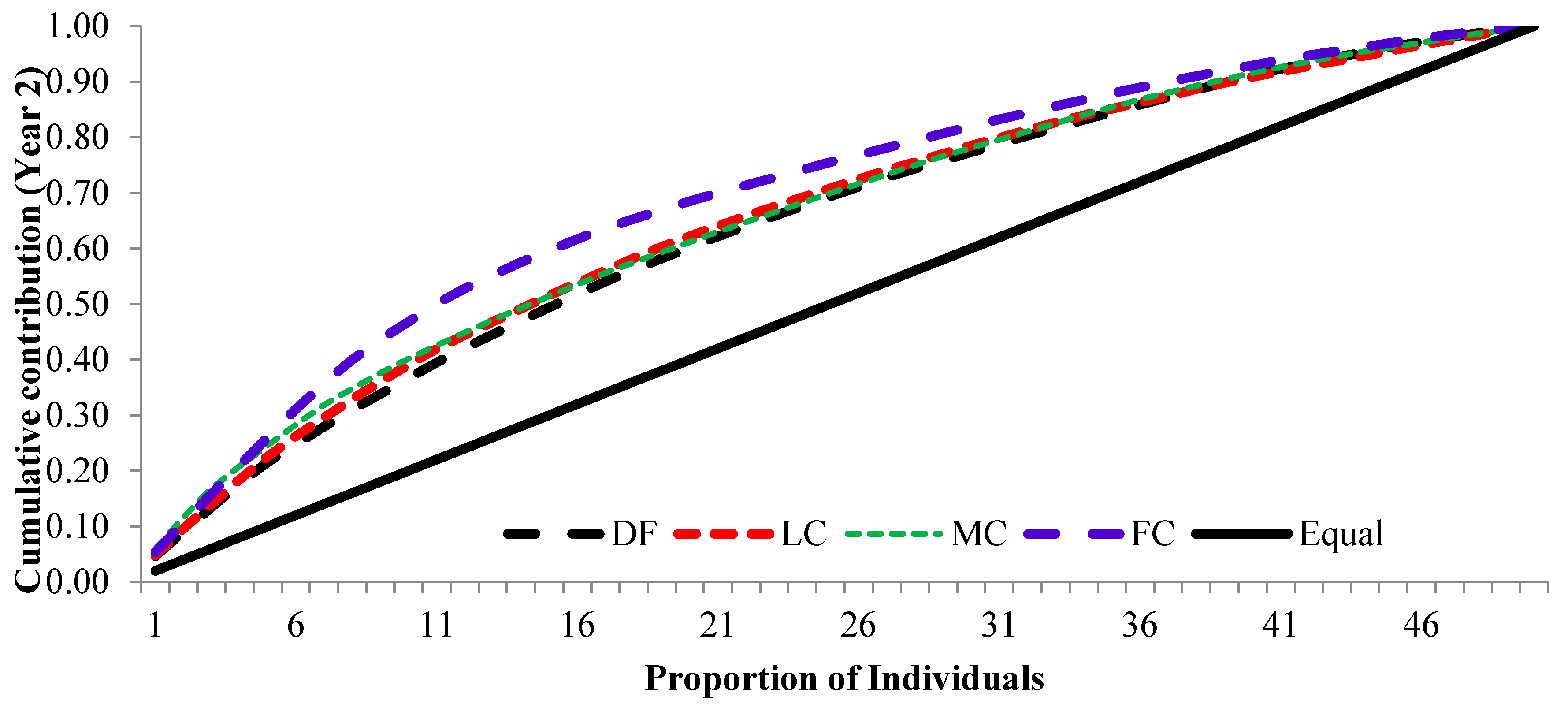
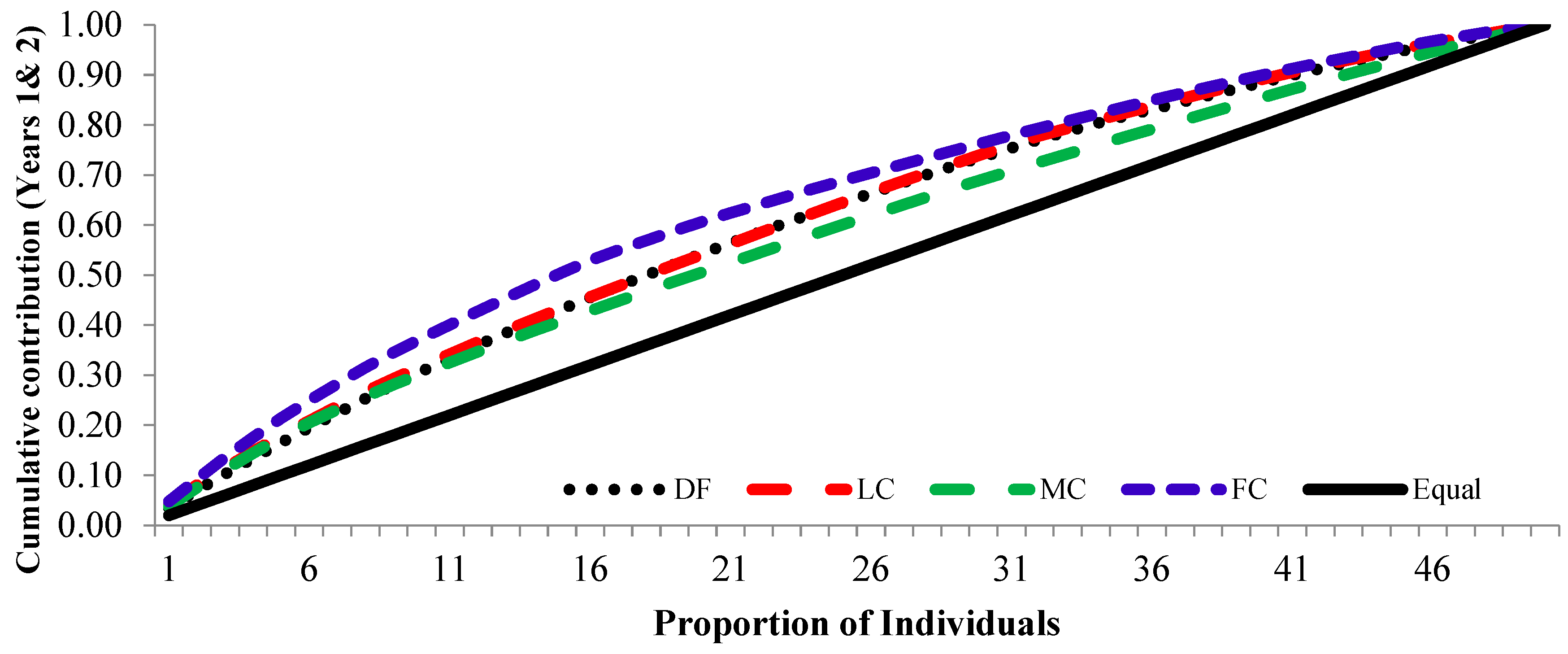
3.3. Cumulative Contribution Curve
4. Discussion
4.1. Cone Production
4.2. Fertility Variation, Effective Number of Parents, and Gene Diversity
4.3. Future Prospect
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boydak, M. Regeneration of Lebanon cedar (Cedrus libani A. Rich.) on karstic lands in Turkey. For. Ecol. Manag. 2003, 178, 231–243. [Google Scholar] [CrossRef]
- Anonymous. Forest Inventory of Turkey, Ankara. 2022. Available online: https://www.ogm.gov.tr/tr/e-kutuphane (accessed on 5 May 2022).
- Koski, V.; Antola, J. National Tree Breeding and Seed Production Programme for Turkey 1994–2003; The Research Directorate of Forest Tree Seeds and Tree Breeding: Ankara, Turkey, 1993. Available online: https://ortohum.ogm.gov.tr (accessed on 5 May 2019).
- Odabasi, T. Research on Cone and Seed Characters of Cedrus libani; General Directorate of Forestry: Ankara, Turkey, 1990. Available online: https://www.ogm.gov.tr (accessed on 5 May 2019).
- Griffin, A.R. Clonal variation in radiata pine seed orchards. I. Some flowering, cone and seed production traits. Aust. For. Res. 1982, 12, 295–302. Available online: http://jkv.50megs.com/afr.html (accessed on 5 May 2019).
- Savolainen, O.; Karkkainen, K.; Harju, A.; Nikkanen, T.; Rusanen, M. Fertility variation in Pinus sylvestris: A test of sexual allocation theory. Am. J. Bot. 1993, 80, 1016–1020. [Google Scholar] [CrossRef]
- Bilir, N. Fertility variation in wild rose (Rosa canina) over habitat classes. Int. J. Agric. Biol. 2011, 13, 110–114. [Google Scholar]
- Bilir, N.; Catal, Y.; Tekocak, S.; Cercioglu, M. Fertility variation in endemic populations of Ehrami black pine (Pinus nigra Arnold. subsp. Pallasiana var. pyramidata). J. For. Res. 2017, 28, 683–686. [Google Scholar] [CrossRef]
- Bilir, N.; Kang, K.S. Estimation of fertility variation by strobili and cone productions in Taurus cedar (Cedrus libani A. Rich.) populations. In Proceedings of the IUFRO Forest Tree Breeding Conference, Prague, Czech Republic, 25–29 August 2014; p. 42. Available online: http://www.iufrobreeding2014.org/ (accessed on 5 May 2019).
- Bilir, N.; Kang, K.S. Fertility variation, seed collection and gene diversity in natural stands of Taurus cedar (Cedrus libani). Eur. J. For. Res. 2021, 40, 199–208. [Google Scholar] [CrossRef]
- Bilir, N.; Ozel, H.B. Fertility variation in a natural stand of Taurus cedar (Cedrus libani A. Rich.). In Proceedings of the International Forestry and Environment Symposium (IFES), Trabzon, Turkey, 7–10 November 2017; Available online: http://www.ktu.edu.tr/ifes2017 (accessed on 5 May 2019).
- Yazici, N.; Bilir, N. Aspectual fertility variation and its effect on gene diversity of seeds in natural stands of Taurus cedar (Cedrus libani A. Rich.). Int. J. Genom. 2017, 2017, 2960624. [Google Scholar] [CrossRef] [PubMed]
- Eler, U. Seed Yield in Calabrian Cluster Pine (Pinus brutia Ten.) by Age; Technical Bulletin No. 225; Forest Research Institute: Antalya, Turkey, 1990; p. 78. Available online: https://centralanatolia.ogm.gov.tr (accessed on 5 May 2019).
- Kang, K.S.; Bila, A.D.; Harju, A.M.; Lindgren, D. Fertility variation in forest tree populations. Forestry 2003, 76, 329–344. [Google Scholar] [CrossRef]
- Bila, A.D.; Lindgren, D. Fertility variation in Milletias thuhlmannii, Brachystegia spiciformis, Brachystegia bohemii and Leucaena leucocephala and its effects on relatedness in seeds. For. Genet. 1998, 5, 119–129. Available online: https://kf.tuzvo.sk/sites/default/files/FG05-2_119-129.pdf (accessed on 5 May 2019).
- Bilir, N.; Kang, K.S.; Lindgren, D. Fertility variation in six populations of Brutian pine (Pinus brutia Ten.) over altitudinal ranges. Euphytica 2005, 141, 163–168. [Google Scholar] [CrossRef]
- Saatcioglu, F. Silviculture-I; Forestry Faculty of Istanbul University Press: Istanbul, Turkey, 1976; Available online: https://cerrahpasa.iuc.edu.tr/ (accessed on 5 May 2019).
- SAS. Statistical Analysis System; SAS Institute, Inc.: Cary, NC, USA, 2004; Available online: https://www.sas.com (accessed on 5 May 2019).
- Rohlf, F.J.; Sokal, R.R. Statistical Tables; Macmillan: New York, NY, USA, 1995; Available online: https://www.scirp.org (accessed on 5 May 2019).
- Kang, K.S.; Lindgren, D. Fertility variation among clones of Korean pine (Pinus koraiensis S. et Z.) and its implications on seed orchard management. For. Genet. 1999, 6, 191–200. Available online: https://kf.tuzvo.sk/sites/default/files/FG06-3_191-200.pdf (accessed on 5 May 2019).
- Kang, K.S. Genetic Gain and Gene Diversity of Seed Orchard Crops. Ph. D Thesis, Swedish University of Agricultural Science, Acta Universitatis Agriculturae Sueciae, Umeå, Sweden, 2001; p. 75. Available online: https://www.upsc.se (accessed on 5 May 2019).
- Shea, K.L. Effects of population structure and cone production on out crossing rates in Engelmann spruce and Subalpine fir. Evolution 1987, 41, 124–136. [Google Scholar] [CrossRef]
- Kamalakannan, R.; Varghese, M.; Park, J.M.; Kwon, S.H.; Song, J.H.; Kang, K.S. Fertility variation and its impact on effective population size in seed stands of Tamarindus indica and Azadirachta indica. Silvae Genet. 2015, 64, 91–99. [Google Scholar] [CrossRef]
- Teodosiu, M.; Botezatu, A.; Ciocîrlan, E.; Mihai, G. Variation of cones production in a Silver fir (Abies alba Mill.) clonal seed orchard. Forests 2023, 14, 17. [Google Scholar] [CrossRef]
- Kang, K.S.; Bilir, N. Seed Orchards (Establishment, Management and Genetics); OGEM-VAK Press: Ankara, Turkey, 2021; Available online: https://www.ogemvak.org.tr/ (accessed on 5 May 2022).
- Kang, K.S.; Lindgren, D. Fertility variation and its effect on the relatedness of seeds in Pinus densiflora, Pinus thunbergii and Pinus koraiensis clonal seed orchards. Silvae Genet. 1998, 47, 196–201. Available online: https://www.degruyter.com (accessed on 5 May 2019).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CCCs * | ||||
|---|---|---|---|---|
| Characteristics | DF | LC | MC | FC |
| H (m) | 20.3 | 21.2 | 20.0 | 21.6 |
| D0 (cm) | 64.1 | 74.9 | 72.1 | 89.4 |
| d1.30 (cm) | 53.9 | 63.4 | 62.4 | 78.2 |
| CD (m) | 7.3 | 8.9 | 8.4 | 10.0 |
| Age (year) | 121 | 131 | 127 | 139 |
| Altitude (m) | 1632 | 1630 | 1628 | 1628 |
| CCCs * | DF | LC | MC | FC | ||||
|---|---|---|---|---|---|---|---|---|
| Years | Year 1 | Year 2 ** | Year 1 | Year 2 | Year 1 | Year 2 | Year 1 | Year 2 |
| 25.2 | 28.3 a | 25.5 | 27.1 a | 24.6 | 29.8 a | 29.5 | 40.3 b | |
| Range | 11–50 | 9–66 | 10–50 | 10–65 | 10–45 | 10–88 | 10–67 | 8–108 |
| CV | 32.8 | 51.4 | 33.5 | 56.0 | 37.5 | 61.1 | 41.0 | 74.6 |
| Source | Variable | df | Sum Square | Mean Square | F | p |
|---|---|---|---|---|---|---|
| Model | year 1 | 3 | 135,077.948 a | 45,025.983 | 509.161 | *** |
| year 2 | 3 | 196,108.588 b | 65,369.529 | 164.498 | *** | |
| Year (Y) | year 1 | 1 | 134,838.434 | 134,838.434 | 1524.774 | *** |
| year 2 | 1 | 191,717.364 | 191,717.364 | 482.444 | *** | |
| CCCs | year 1 | 1 | 238.595 | 238.595 | 2.698 | NS |
| year 2 | 1 | 3735.869 | 3735.869 | 9.401 | ** | |
| Y × CCCs interaction | year 1 | 1 | 0.919 | 0.919 | 0.010 | NS |
| year 2 | 1 | 655.355 | 655.355 | 1.649 | NS | |
| Error | year 1 | 197 | 17,421.052 | 88.432 | ||
| year 2 | 197 | 78,285.412 | 397.388 | |||
| Total | year 1 | 200 | 152,499.000 | |||
| year 2 | 200 | 274,394.000 |
| CCCs (I) | CCCs (J) | Year 1 | Year 2 | ||
|---|---|---|---|---|---|
| Mean Difference (I–J) | Sig. (p) * | Mean Difference (I–J) | Sig. (p) * | ||
| LC | −0.26 | NS | 1.14 | NS | |
| DF | MC | 0.60 | NS | −1.48 | NS |
| FC | −4.28 | NS | −12.00 | p ≤ 0.05 | |
| DF | 0.26 | NS | −1.14 | NS | |
| LC | MC | 0.86 | NS | −2.62 | NS |
| FC | −4.02 | NS | −13.14 | p ≤ 0.05 | |
| DF | −0.60 | NS | 1.48 | NS | |
| MC | LC | −0.86 | NS | 2.62 | NS |
| FC | −4.88 | NS | −10.52 | p ≤ 0.05 | |
| DF | 4.28 | NS | 12.00 | p ≤ 0.05 | |
| FC | LC | 4.02 | NS | 13.14 | p ≤ 0.05 |
| MC | 4.88 | NS | 10.52 | p ≤ 0.05 | |
| CCCs * | DF | LC | MC | FC | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Years | Year 1 | Year 2 | 1 and 2 | Year 1 | Year 2 | 1 and 2 | Year 1 | Year 2 | 1 and 2 | Year 1 | Year 2 | 1 and 2 |
| Ψ | 1.11 | 1.26 | 1.12 | 1.11 | 1.31 | 1.14 | 1.14 | 1.37 | 1.09 | 1.16 | 1.55 | 1.26 |
| Np | 45.23 | 39.71 | 44.60 | 45.04 | 38.25 | 43.73 | 43.95 | 36.62 | 45.79 | 42.92 | 32.35 | 39.60 |
| Nr ** | 0.91 | 0.79 | 0.89 | 0.90 | 0.77 | 0.87 | 0.88 | 0.73 | 0.92 | 0.86 | 0.65 | 0.79 |
| GD | 0.989 | 0.987 | 0.989 | 0.989 | 0.987 | 0.989 | 0.989 | 0.986 | 0.989 | 0.988 | 0.984 | 0.988 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yazici, N.; Bilir, N. Impact of Crown Closure on Cone Production and Effective Number of Parents in Natural Stands of Taurus Cedar (Cedrus libani A. Rich.). Forests 2023, 14, 1130. https://doi.org/10.3390/f14061130
Yazici N, Bilir N. Impact of Crown Closure on Cone Production and Effective Number of Parents in Natural Stands of Taurus Cedar (Cedrus libani A. Rich.). Forests. 2023; 14(6):1130. https://doi.org/10.3390/f14061130
Chicago/Turabian StyleYazici, Nilufer, and Nebi Bilir. 2023. "Impact of Crown Closure on Cone Production and Effective Number of Parents in Natural Stands of Taurus Cedar (Cedrus libani A. Rich.)" Forests 14, no. 6: 1130. https://doi.org/10.3390/f14061130
APA StyleYazici, N., & Bilir, N. (2023). Impact of Crown Closure on Cone Production and Effective Number of Parents in Natural Stands of Taurus Cedar (Cedrus libani A. Rich.). Forests, 14(6), 1130. https://doi.org/10.3390/f14061130