
Resilience of Pyrenean Forests after Recurrent Historical Deforestations


Abstract
1. Introduction
2. Study Area
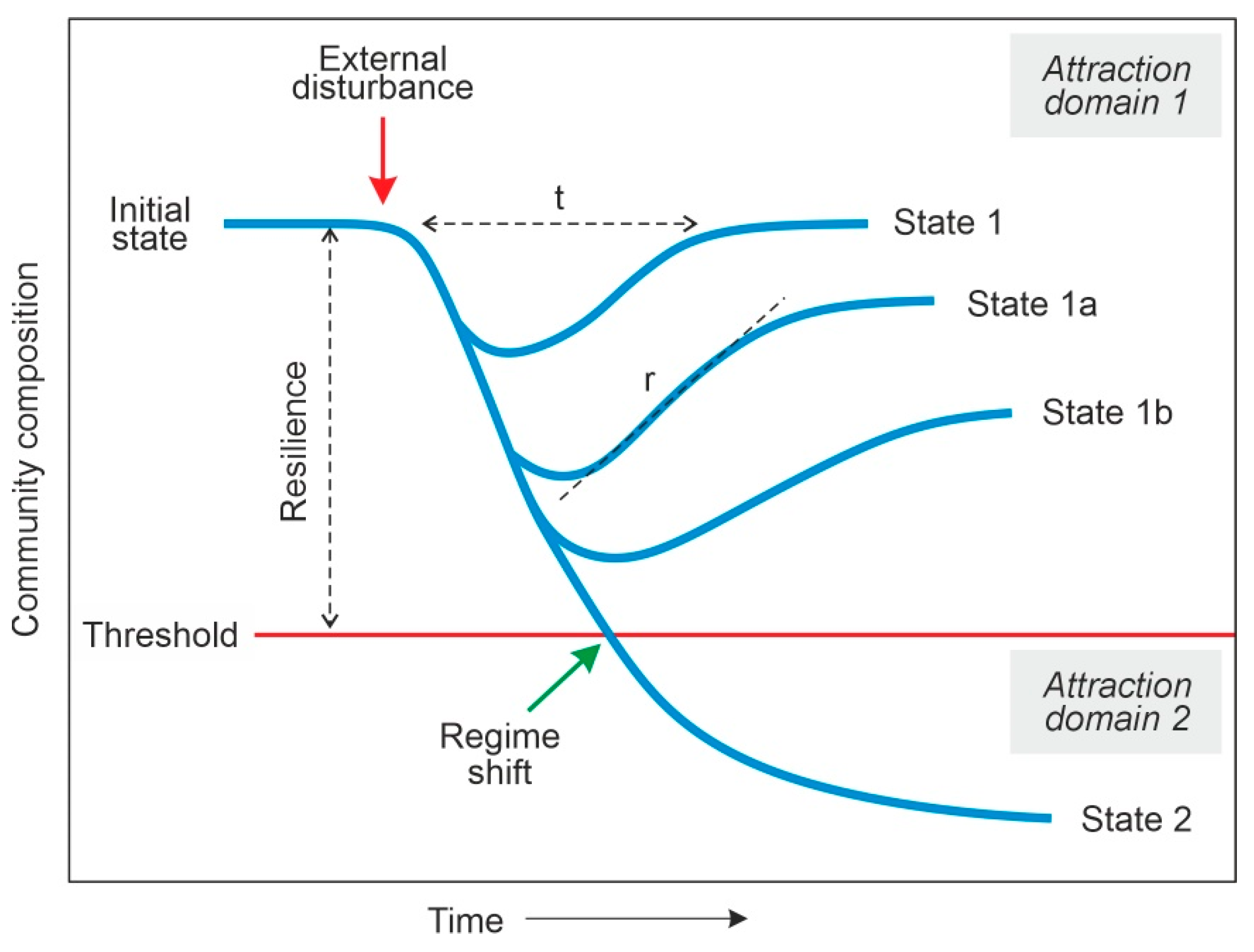
3. Methods
- Bulk resilience. The total forest pollen is a reliable proxy for regional forest cover and fluctuations in this parameter are considered to be indicative of a bulk regime shift if forests are eventually replaced by a different non-forested vegetation type, or bulk forest resilience, if forest cover recovers after each clearing event, regardless of the type of forest that finally establishes.
- Mosaic resilience. The three main forest types described above (Cf, Msf, Sdf), are arranged in a characteristic mosaic pattern, which is characterized palynologically by the mixture of pollen from the dominant taxa of each of these forest types. An eventual post-clearing disappearance of the pollen from one or two of the three forest types would imply a drastic mosaic reorganization or the removal of the mosaic itself and its replacement by a single regional forest type (regime shift). In contrast, the post-deforestation continuity of the three forest types with minor variations in their respective pollen percentages is considered to be indicative of the regional persistence of the three forest types with minor reorganizations in their respective patch extent and arrangement (mosaic resilience).
- Community resilience. Intra-community resilience is difficult to evaluate, as it is not possible to identify the particular forest patches acting as pollen sources for the different pollen types. Therefore, the individual pollen components of each forest type present in the pollen record are representative of the assemblage in all regional forest patches together, which defines the metacommunity signal [32]. In this case, a metacommunity-scale regime shift would be recorded by the replacement of the dominant taxa in a given forest type, whereas internally fluctuating relative abundances would be indicative of metacommunity resilience.
4. Results
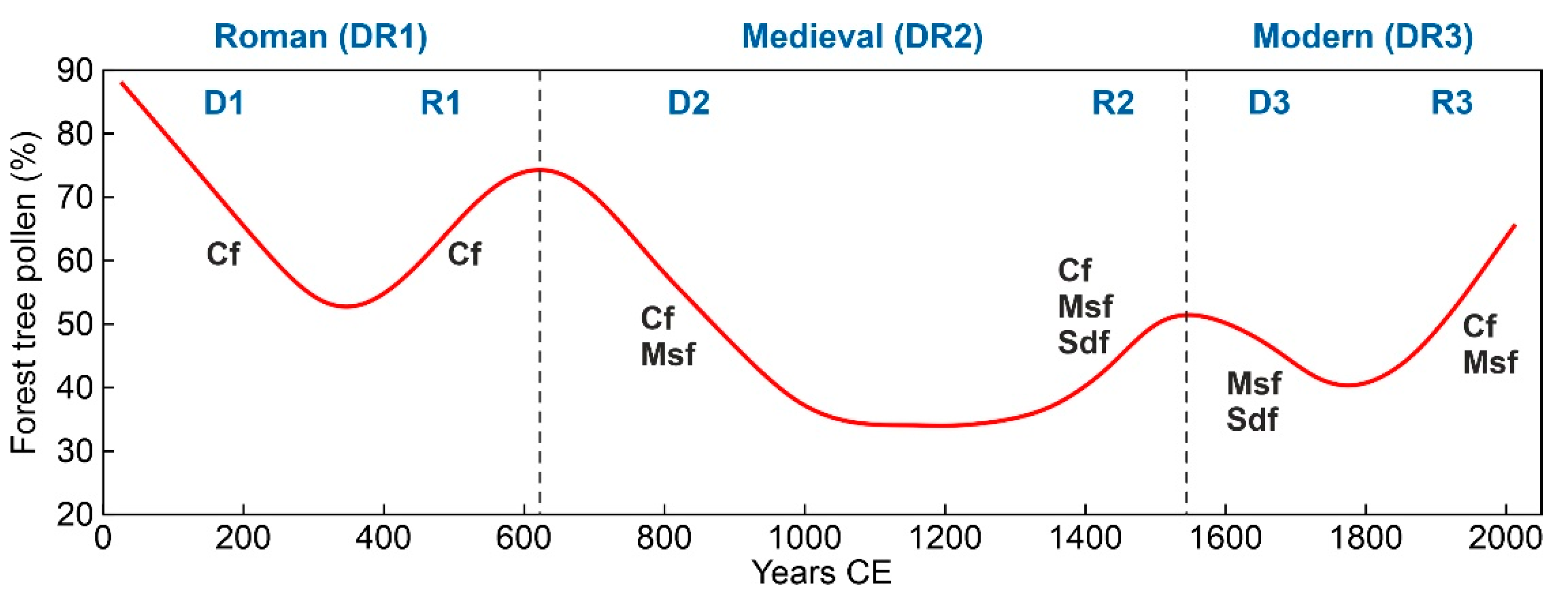
4.1. Bulk Resilience
4.2. Mosaic Resilience
4.3. Community Resilience
5. Discussion and Conclusions
5.1. Comparisons with Other Pyrenean Records
5.2. Applications to Conservation
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Grimm, V.; Schmidt, E.; Wissel, C. On the application of stability concepts in ecology. Ecol. Model. 1992, 63, 143–161. [Google Scholar] [CrossRef]
- Gunderson, L.H. Ecological resilience—In theory and application. Annu. Rev. Ecol. Syst. 2000, 31, 425–439. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and stability of ecological systems. Ann. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Briggs, R.; Carpenter, S.R.; Brock, W.A. Turning back from the brink: Detecting and impeding regime shift in time to advert it. Proc. Natl. Acad. Sci. USA 2009, 106, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Rull, V. Quaternary Ecology, Evolution, and Biogeography; Elsevier: London, UK; Academic Press: London, UK, 2020. [Google Scholar]
- Falk, D.A.; van Mantgem, P.J.; Keeley, J.E.; Gregg, R.M.; Guiterman, C.H.; Tepley, A.J.; Young, D.J.N.; Marshall, L.A. Mechanisms of forest resilience. Forest Ecol. Manag. 2022, 512, 120–129. [Google Scholar] [CrossRef]
- Rull, V. Quaternary palaeoecology and ecological theory. Orsis 1990, 5, 91–111. [Google Scholar]
- Rull, V. Successional patterns of the Gran Sabana (southeastrn Venezuela) vegetation during the last 5000 years, and its responses to climatic fluctuations and fire. J. Biogeogr. 1992, 19, 329–338. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T. What is long term in ecology? Trends Ecol. Evol. 2011, 26, 3–4. [Google Scholar] [CrossRef]
- Davis, M.B. Quaternary history and the stability of forest communities. In Forest Succession, Concepts and Applications; Shugar, D.B., Botkin, D.B., Eds.; Springer: New York, NY, USA, 1981; pp. 132–153. [Google Scholar]
- Davis, M.B. Climatic instability, time lags, and community disequilibrium. In Community Ecology; Diamond, J., Case, T.J., Eds.; Harper & Row: New York, NY, USA, 1984; pp. 269–284. [Google Scholar]
- Svenning, J.-C.; Fløjgaard, C.; Baselga, A. Climate, history and neutrality as drivers of mammal beta diversity in Europe: Insights from multiscale deconstruction. J. Anim. Ecol. 2011, 80, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Rull, V. Community ecology: Diversity and dynamics over time. Comm. Ecol. 2012, 13, 102–116. [Google Scholar] [CrossRef]
- Jackson, S.T.; Blois, J.L. Community ecology in a changing environment: Perspectives from the Quaternary. Proc. Natl. Acad. Sci. USA 2016, 112, 4015–4021. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T. Climatic and anthropogenic drivers of forest succession in the Iberian Pyrenees during the last 500 years: A statistical approach. Forests 2022, 13, 622. [Google Scholar] [CrossRef]
- Trapote, M.C.; Rull, V.; Giralt, S.; Corella, J.P.; Montoya, E.; Vegas-Vilarrúbia, T. High-resolution (sub-decadal) pollen analysis of varved sediments from Lake Montcortès (southern Pyrenean flank): A fine-tuned record of landscape dynamics and human impact during the last 500 years. Rev. Palaeobot. Palynol. 2018, 259, 207–222. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T. Conifer forest dynamics in the Iberian Pyrenees during the Middle Ages. Forests 2021, 12, 1685. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T.; Corella, J.P.; Valero-Garcés, B. Bronze Age to Medieval vegetation dynamics and landscape anthropization in the southern-central Pyrenees. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 571, 110392. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T.; Corella, J.P.; Trapote, M.C.; Montoya, E.; Valero-Garcés, B.L. A unique Pyrenean varved record provides a detailed reconstruction of Mediterranean vegetation over the last three millennia. Quat. Sci. Rev. 2021, 268, 107128. [Google Scholar] [CrossRef]
- Gutièrrez, F.; Linares, R.; Roquè, C.; Zarroca, M.; Rosell, J.; Galve, J.P.; Carbonel, D. Investigating gravitational grabens related to lateral spreading and evaporite dissolution subsidence by means of detailed mapping, trenching, and electrical resistivity tomography (Spanish Pyrenees). Lithosphere 2012, 4, 331–353. [Google Scholar] [CrossRef]
- Rull, V.; Sigró, J.; Vegas-Vilarrúbia, T. Present climate of Lake Montcortès (central Pyrenees): Paleoclimatic relevance and insights on future warming. Geogr. Res. Lett. 2022. [Google Scholar] [CrossRef]
- Vigo, J.; Ninot, J.M. Los Pirineos. In La Vegetación de España; Peinado, M., Rivas-Martínez, F., Eds.; Univ. Alcalá de Henares: Madrid, Spain, 1987; pp. 349–389. [Google Scholar]
- Carreras, J.; Vigo, J.; Ferré, A. Manual dels Hàbitats de Catalunya, vols. I-VIII.; Dep. Medi Amb. Habit. Gen: Catalunya, Barcelona, 2005–2006. [Google Scholar]
- Folch, R. La Vegetació dels Països Catalans; Ketres: Barcelona, Spain, 1981. [Google Scholar]
- Folch, R.; Franquesa, T.; Camarassa, J.M. Història Natural dels Països Catalans, vol. 7: Vegetació; Enciclopèdia Catalana: Barcelona, Spain, 1984. [Google Scholar]
- Masalles, R.M.; Carreras, J.; Farràs, A.; Ninot, J.M.; Camarassa, J.M. Història Natural dels Països Catalans, vol. 6: Plantes Superiors; Enciclopèdia Catalana: Barcelona, Spain, 1988. [Google Scholar]
- Ninot, J.M.; Carrillo, E.; Ferré, A. The Pyrenees. In Vegetation of the Iberian Peninsula; Loidi, J., Ed.; Springer Nature: Cham, Switzerland, 2017; pp. 323–366. [Google Scholar]
- Rull, V.; Sacristan-Soriano, O.; Sánchez-Melsió, A.; Borrego,, C.M.; Vegas-Vilarrúbia, T. Bacterial phylogenetic markers in lake sediments provide direct evidence for historical hemp retting. Quat. Sci. Rev. 2022, 295, 107803. [Google Scholar] [CrossRef]
- Mercadé, A.; Vigo, J.; Rull, V.; Vegas-Vilarrúbia, T.; Garcés, S.; Lara, A.; Cañellas-Boltà, N. Vegetation and landscape around Lake Montcortès (Catalan pre-Pyrenees) as a tool for palaeoecological studies of Lake sediments. Collect. Bot. 2013, 32, 87–101. [Google Scholar] [CrossRef]
- Rull, V.; Trapote, M.C.; Safont, E.; Cañellas-Boltà, N.; Pérez-Zanón, N.; Sigró, J.; Buchaca, T.; Vegas-Vilarrúbia, T. Seasonal patterns of pollen sedimentation in Lake Montcortès (Central Pyrenees) and potential applications to high-resolution paleoecology: A 2-year pilot study. J. Paleolimnol. 2017, 57, 95–108. [Google Scholar] [CrossRef]
- Holyoak, M.; Leibold, M.A.; Holr, R.D. Metacommunities: Spatial Dynamics and Eological Communities; Chicago University Press: Chicago, IL, USA, 2005. [Google Scholar]
- Hammer, Ø. Paleontological Statistics, Version 4.12; Reference Manual; University of Oslo: Oslo, Norway, 2022. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T. Paleontological Data Analysis; Blackwell: London, UK, 2006. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Paleontol. Electr. 2001, 4, 1–9. [Google Scholar]
- Martín-Chivelet, J.; Muñoz-García, M.B.; Edwards, L.; Turrero, M.J.; Ortega, A.I. Land surface temperature changes in northern Iberia since 4000 yr BP, based on δ13C of speleothems. Glob. Planet. Chang. 2011, 77, 1–12. [Google Scholar] [CrossRef]
- Rull, V. Microrefugia. J. Biogeogr. 2009, 36, 481–484. [Google Scholar] [CrossRef]
- Hanski, I. Metappulation Ecology; Oxford University Press: Oxford, NY, USA, 1999. [Google Scholar]
- Alexander, H.M.; Foster, B.L.; Ballantyne, F.; Collins, C.D.; Antonovics, J.; Holt, R.D. Metapopulations and metacommunities: Combining spatial and temporal perspectives in plant ecology. J. Ecol. 2012, 100, 88–103. [Google Scholar] [CrossRef]
- García-Ruiz, J.M.; Tomás-Faci, G.; Diarte-Blasco, P.; Montes, L.; Domingo, R.; Sebastián, M.; Lasanta, T.; González-Sampériz, P.; López-Moreno, J.I.; Arnaez, J.; et al. Transhumance and long-term deforestation in the subalpine belt of the central Spanish Pyrenees: An interdisciplinary approach. Catena 2020, 195, 104744. [Google Scholar] [CrossRef]
- González-Sampériz, P.; Aranbarri, J.; Pérez-Sanz, A.; Gil-Romera, G.; Moreno, A.; Leunda, M.; SEvilla-Callejo, M.; Corella, J.P.; Morellón, M.; Oliva, B.; et al. Environmental and climate change in the southern Central Pyrenees since the Last Glacial Maximum: A review from lake records. Catena 2017, 149, 668–688. [Google Scholar] [CrossRef]
- González-Sampériz, P.; Montes, L.; Aranbarri, J.; Leunda, M.; Domingo, R.; Laborda, R.; Sanjuan, Y.; Gil-Romera, G.; Lasanta, T.; García-Ruiz, J.M. Escenarios, tiempo e indicadores paleoambientales para la identificación del Antropoceno en el paisaje vegetal del Pirineo central (NE Iberia). Geogr. Res. Lett. 2019, 45, 167–193. [Google Scholar]
- Rull, V.; Cañellas-Boltà, N.; Vegas-Vilarrúbia, T. Late-Holocene forest resilience in the central Pyrenean highlands as deduced from pollen analysis Lake Sant Maurici sediments. Holocene 2021, 31, 1797–1803. [Google Scholar] [CrossRef]
- Büntgen, U.; Krusic, P.J.; Verstege, A.; Sangüesa-Barreda, G.; Wagner, S.; Camarero, J.J.; Ljungqvist, F.C.; Zorita, E.; Oppenheimer, C.; Konter, O.; et al. New tree-ring evidence from the Pyrenees reveals western Mediterranean climate variability since Medieval times. J. Clim. 2017, 30, 5295–5318. [Google Scholar] [CrossRef]
- Vegas-Vilarrúbia, T.; Corella, J.P.; Sigró, J.; Rull, V.; Dorado-Liñán, I.; Valero-Garcés, B.L.; Gutiérrez, E. Regional precipitation trends since 1500 CE, as reconstructed from calcite sublayers of a varved Mediterranean lake record (central Pyrenees). Sci. Total Environ. 2022, 826, 153773. [Google Scholar] [CrossRef] [PubMed]
- Steffen, W.; Rockström, J.; Richardson, K.; Lenton, T.M.; Folke, C.; Liverman, D.; Summerhayes, C.P.; Barnosky, A.D.; Cornell, S.E.; Crucifix, M.; et al. Trajectories of the earth System in the Anthropocene. Proc. Natl. Acad. Sci. USA 2018, 115, 8252–8259. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dates (CE) | Total Forest Pollen (%) | Time (yr) | Rates (% d−1) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DR Cycle | Id | Md | Fd | Ip | Mp | Fp | Dp | Rp | Np | Dt | Rt | St | Dr | Rr | Sr |
| DR1 | 30 | 320 | 620 | 88 | 38 | 84 | 50 | 46 | −4 | 290 | 300 | 1.0 | 1.7 | 1.5 | 0.9 |
| DR2 | 620 | 1050–1300 | 1550 | 84 | 23 | 65 | 61 | 42 | −19 | 430 | 250 | 0.6 | 1.4 | 1.7 | 1.2 |
| DR3 | 1550 | 1780 | 2000 | 65 | 27 | 73 | 38 | 46 | 8 | 230 | 220 | 1.0 | 1.7 | 2.1 | 1.3 |
| Roman | Medieval | Modern | |||||
|---|---|---|---|---|---|---|---|
| Forest type | Overall | D1 | R1 | D2 | R2 | D3 | R3 |
| Cf | 0.823 ** | 0.862 ** | 0.798 ** | 0.890 ** | 0.749 ** | 0.338 | 0.759 ** |
| Msf | 0.479 ** | 0.056 | 0.651 | 0.540 ** | 0.723 ** | 0.748 ** | 0.670 ** |
| Sdf | 0.083 | 0.424 | 0.449 | −0.321 | 0.716 ** | 0.771 ** | 0.218 |
| Rf | 0.101 | −0.008 | 0.455 | 0.127 | −0.377 | 0.204 | 0.058 |
| Roman | Medieval | Modern | |||||
|---|---|---|---|---|---|---|---|
| Tree Taxa | Overall | D1 | R1 | D2 | R2 | D3 | R3 |
| Pinus | 0.824 ** | 0.856 ** | 0.803 * | 0.889 ** | 0.748 ** | 0.328 | 0.759 ** |
| Abies | 0.456 ** | 0.728 * | −0.306 | 0.292 | 0.223 | 0.178 | 0.000 |
| Quercus (e) | 0.479 ** | 0.056 | 0.651 | 0.541 ** | 0.723 ** | 0.748 ** | 0.670 ** |
| Quercus (d) | 0.030 | 0.303 | 0.280 | −0.570 ** | 0.726 ** | 0.500 ** | 0.339 * |
| Betula | −0.031 | 0.425 | 0.691 | 0.206 | 0.536 ** | 0.489 ** | −0.211 |
| Corylus | 0.213 ** | 0.528 | 0.360 | 0.033 | 0.304 | 0.392 * | 0.193 |
| Fagus | 0.211 ** | 0.165 | −0.308 | 0.174 | 0.193 | 0.179 | 0.095 |
| Alnus | 0.172 * | 0.142 | 0.447 | −0.048 | −0.133 | 0.050 | 0.179 |
| Salix | 0.047 | −0.246 | 0.329 | 0.250 | 0.062 | 0.190 | −0.288 |
| Ulmus | 0.170 * | 0.000 | 0.000 | 0.000 | −0.628 ** | 0.179 | 0.309 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rull, V.; Vegas-Vilarrúbia, T. Resilience of Pyrenean Forests after Recurrent Historical Deforestations. Forests 2023, 14, 567. https://doi.org/10.3390/f14030567
Rull V, Vegas-Vilarrúbia T. Resilience of Pyrenean Forests after Recurrent Historical Deforestations. Forests. 2023; 14(3):567. https://doi.org/10.3390/f14030567
Chicago/Turabian StyleRull, Valentí, and Teresa Vegas-Vilarrúbia. 2023. "Resilience of Pyrenean Forests after Recurrent Historical Deforestations" Forests 14, no. 3: 567. https://doi.org/10.3390/f14030567
APA StyleRull, V., & Vegas-Vilarrúbia, T. (2023). Resilience of Pyrenean Forests after Recurrent Historical Deforestations. Forests, 14(3), 567. https://doi.org/10.3390/f14030567