
Effect of Pruning Treatment on Growth Characteristics and Metabolites in Eucommia ulmoides Oliver (E. ulmoides)


 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Materials
2.2. Measurement of Growth and Photosynthetic Characteristics
2.2.1. Growth Indices
2.2.2. Photosynthetic Parameters
2.2.3. Chlorophyll Fluorescence Parameters
2.2.4. Photosynthetic Pigments Content
2.3. Determination of Primary Metabolites
2.4. Determination of Secondary Metabolites
2.5. Statistical Analysis
3. Results
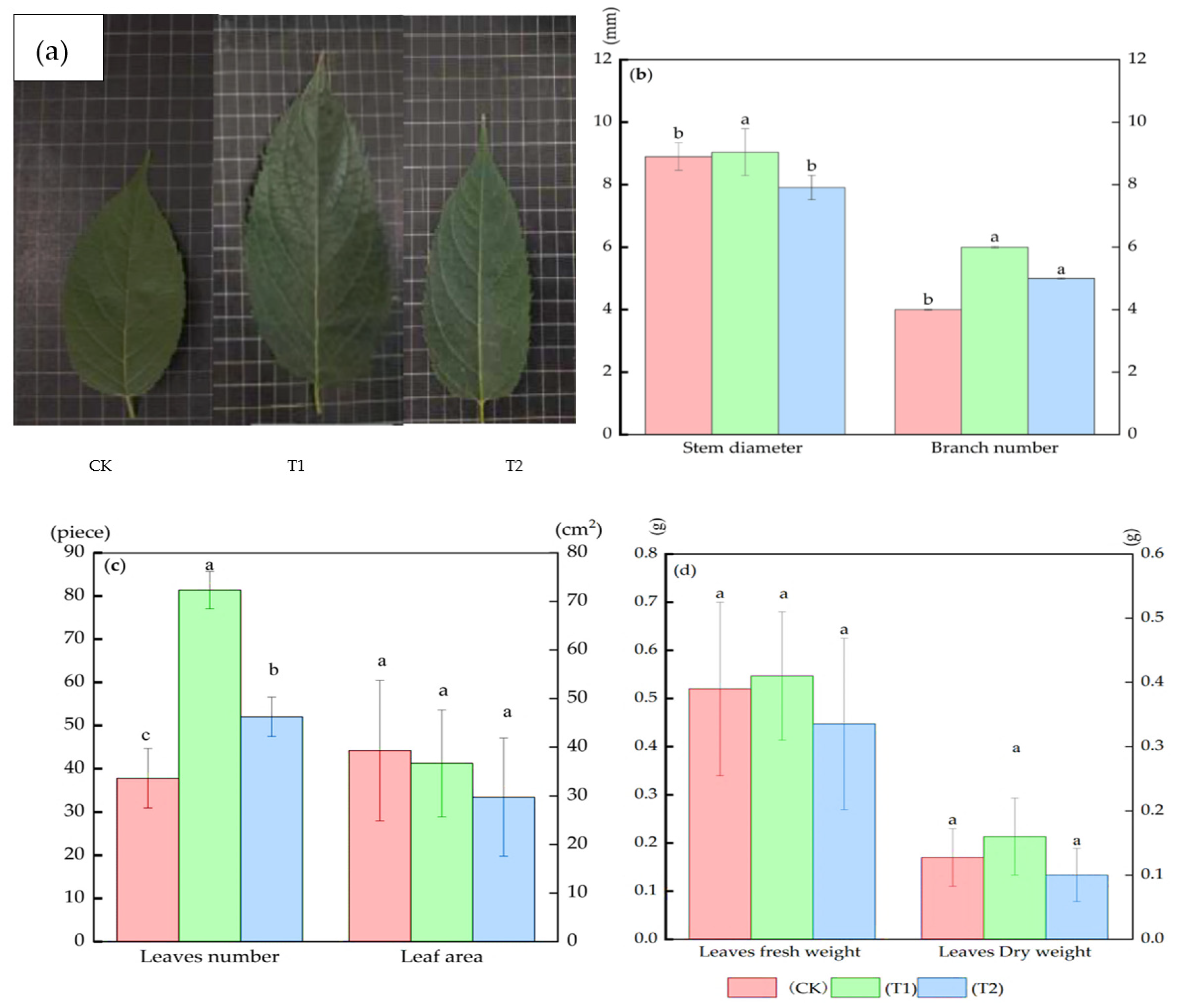
3.1. Growth Indices Changes in Eucommia ulmoides Oliver (E. ulmoides) in Different Treatments
3.2. Photosynthetic Parameters Changes in Eucommia ulmoides Oliver (E. ulmoides) in Different Treatments
3.2.1. Photosynthetic Parameters
3.2.2. Photosynthetic Pigments Contents
3.3. Primary Metabolites Changes in Eucommia ulmoides Oliver (E. ulmoides) in Different Treatments
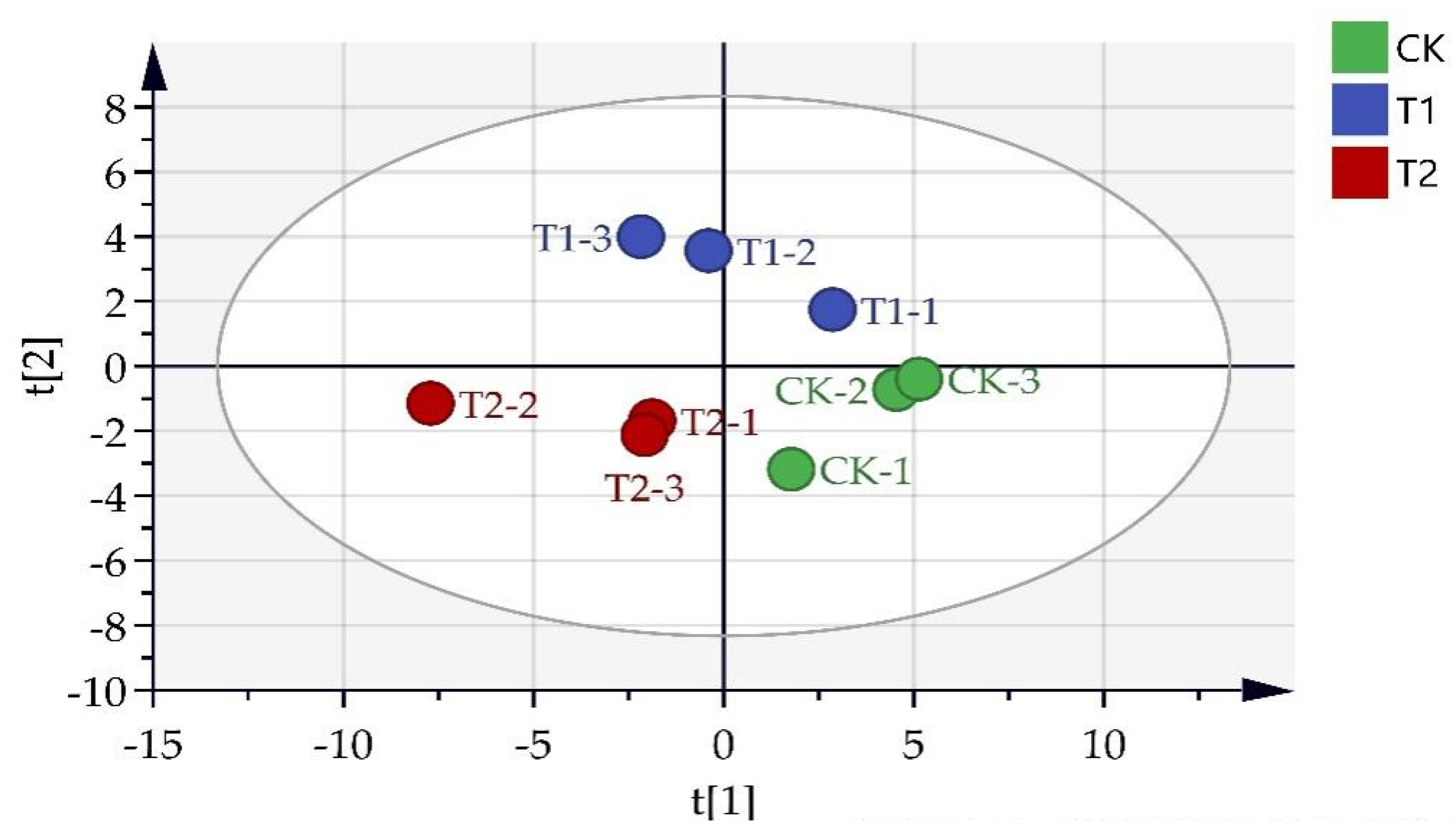
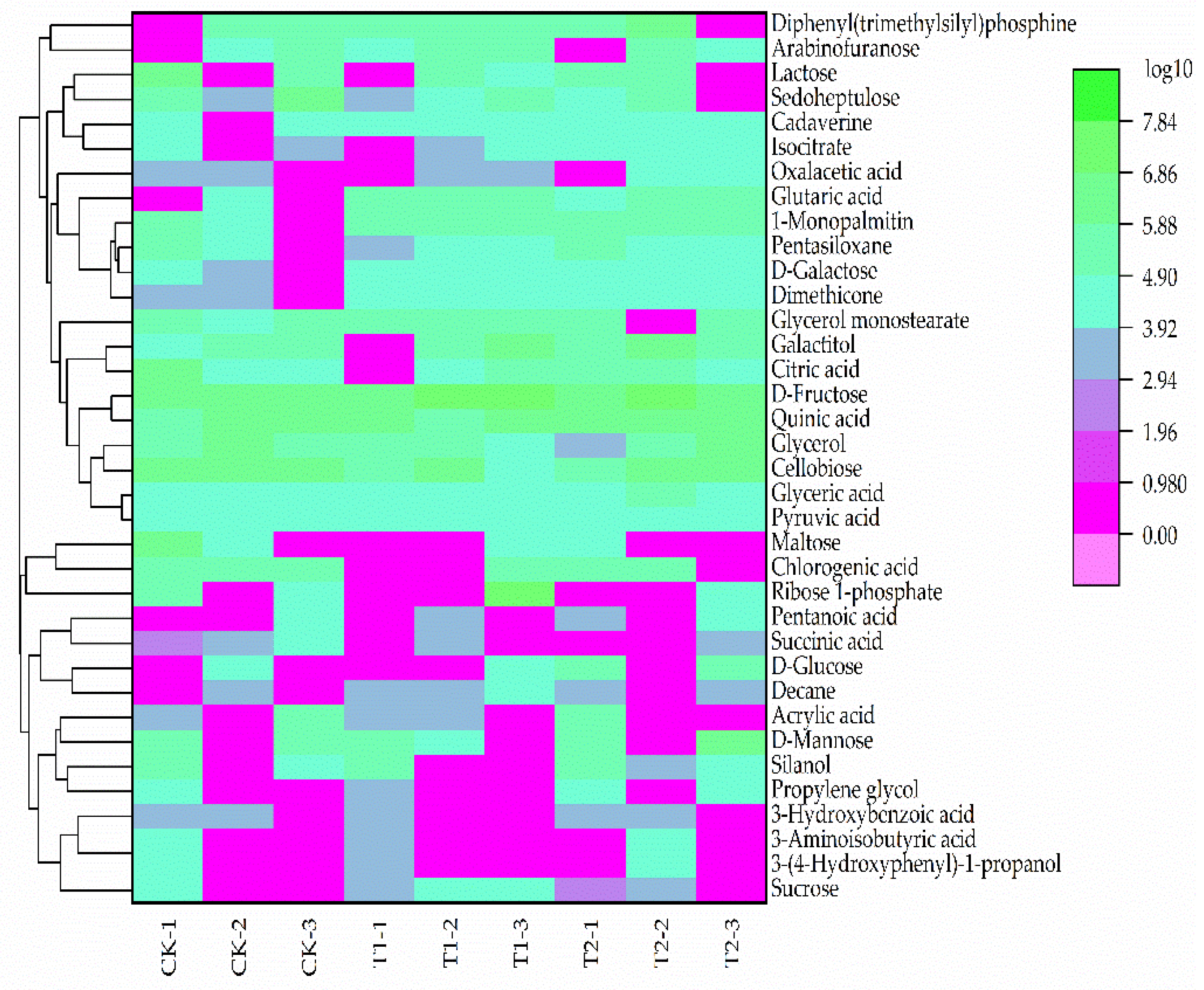
3.3.1. Primary Metabolites Analysis
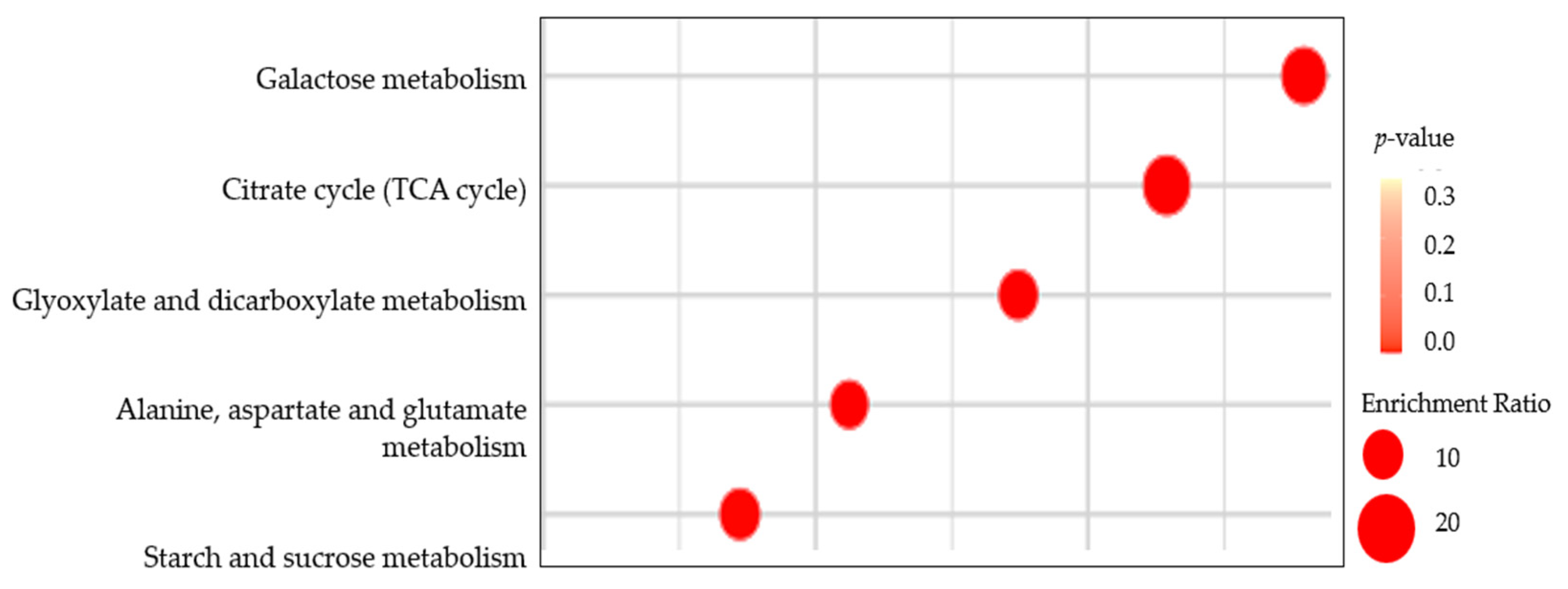
3.3.2. Primary Metabolites Pathways Analysis
3.3.3. Correlations Analysis between Primary Metabolites and Growth Indices, Photosynthetic Parameters in T2 Treatment
3.4. Secondary Metabolites Yield Changes in Eucommia ulmoides Oliver (E. ulmoides) in Different Treatments
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, J.K.; Shi, X.L.; Donkor, P.O.; Ding, L.Q.; Qiu, F. Four new megastigmane glycosides from the leaves of Eucommia ulmoides Oliver. Phytochem. Lett. 2018, 27, 208–213. [Google Scholar] [CrossRef]
- Meng, Y.D.; Du, Q.X.; Du, H.Y.; Wang, Q.; Wang, L.; Du, L.Y.; Liu, P.F. Analysis of chemotypes and their markers in leaves of core collections of Eucommia ulmoides Oliver using metabolomics. Front. Plant Sci. 2023, 13, 1029907. [Google Scholar] [CrossRef]
- Huang, L.; Lyu, Q.; Zheng, W.; Yang, Q.; Cao, G. Traditional application and modern pharma cological research of Eucommia ulmoides Oliver. Chin. Med. 2021, 16, 73. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Mariyama, M.; Hirooka, Y.; Iijima, M. Root pruning is effective in alleviating the inhibition of soybean growth caused by anaerobic stress for a short period. J. Integr. Agric. 2023, 22, 1035–1044. [Google Scholar] [CrossRef]
- Badrulhisham, N.; Othman, N. Knowledge in tree pruning for sustainable practices in urban setting: Improving our quality of life. Procedia-Soc. Behav. Sci. 2016, 234, 210–217. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: Cellular origins and molecular mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef]
- Wen, Y.B.; Jiang, H.L.; Long, S.H.; Chen, S.X.; Peng, Z. Effect of Mounding and Top-Pruning on Survival and Growth of Manchurian Ash Seedlings Planted under the Secondary Forest of the Species. Sci. Silvae Sin. 2015, 51, 104–113. [Google Scholar]
- Nie, J.; Li, Z.; Zhang, Y.; Zhang, D.; Xu, S.; He, N.; Zhan, Z.; Dai, J.; Li, C.; Li, W.; et al. Plant pruning affects photosynthesis and photo assimilate partitioning in relation to the yield formation of field-grown cotton. Ind. Crops Prod. 2021, 173, 114087. [Google Scholar] [CrossRef]
- Mehtan, N.; Jain, A.; Rajkumar, M. Impact of pruning of Diospyros melanoxylon Roxb. (Tendu) bushes on yield and quality of leaves in Maharashtra. J. Pharmacogn. Phytochem. 2020, 9, 1360–1365. [Google Scholar] [CrossRef]
- Morgani, M.B.; Fanzone, M.; Peña, J.E.; Sari, S.; Gallo, A.E.; Tournier, M.G.; Prieto, J.A. Late pruning modifies leaf to fruit ratio and shifts maturity period, affecting berry and wine composition in Vitis vinífera L. cv. ‘Malbec’ in Mendoza, Argentina. Sci. Hortic. 2023, 313, 111861. [Google Scholar] [CrossRef]
- Cinosi, N.; Moriconi, F.; Farinelli, D.; Marchionni, D.; Lodolini, E.M.; Rosati, A.; Famiani, F. Effects of summer pruning on the water status and physiology of olive trees and on fruit characteristics and oil quality. Sci. Hortic. 2024, 324, 112612. [Google Scholar] [CrossRef]
- Danziger, N.; Bernstein, N. Shape Matters: Plant Architecture Affects Chemical Uniformity in Large-Size Medical Cannabis Plants. Plants 2021, 10, 1834. [Google Scholar] [CrossRef] [PubMed]
- Danziger, N.; Bernstein, N. Plant Architecture Manipulation Increases Cannabinoid Standardization in ‘drug-Type’ MedicalCannabis. Ind. Crops Prod. 2021, 167, 113528. [Google Scholar] [CrossRef]
- Jia, X.Y.; Zhou, J.J.; Su, T.T.; Zhou, Y.; Chen, J.B.; Ma, H.B.; Ma, J.L.; Wang, X.F. Effects of different cropping densities on the habitat of artificial Caragana intermedia in desert steppe. Acta Ecol. Sin. 2020, 40, 4126–4136. [Google Scholar]
- Dong, X.Y.; Li, P.H.; Wang, Y.Z.; Liu, C.L.; Wang, S.Q.; Lin, Q. Effects of water stress on Water Utilization Efficiency and Carboxylation Efficiency in the peach leaves of different growth type. J. Irrig. Drain. 2005, 24, 67–69. [Google Scholar]
- Cen, H.Y.; Weng, H.Y.; Yao, J.N.; He, M.B.; Lv, J.W.; Hua, S.J.; Li, H.Y.; He, Y. Chlorophyll fluorescence imaging uncov ers photosynthetic fingerprint of Citrus Huanglongbing. Front. Plant Sci. 2017, 29, 1509. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Sun, D.; Cen, H.; Xu, H.; Weng, H.; Yuan, F.; He, Y. Phenotyping of Arabidopsis drought stress re sponse using kinetic chlorophyll fluorescence and multicolor fluorescence imaging. Front. Plant Sci. 2018, 9, 603. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.X. Effects of Exogenous NO on Physiological Characteristics of Pinus Koreiansis under UV-B Stress. Master’s Thesis, Northeast Fortheast University, Harbin, China, 2011. [Google Scholar]
- Liu, Y.; Liu, J.; Abozeid, A.; Wu, K.X.; Guo, X.R.; Mu, L.Q.; Tang, Z.H. UV-B Radiation Largely Promoted the Transformation of Primary Metabolites to Phenols in Astragalus mongholicus seedlings. Biomolecules 2020, 10, 504. [Google Scholar] [CrossRef]
- Zhang, Q.C. Study on extraction technology of chemical active components from Eucommia ulmoides Oliver leaves. Liaoning Chem. Ind. 2022, 51, 1379–1381. [Google Scholar]
- Zou, K.; Liu, X.; Zhang, D.; Yang, Q.; Fu, S.; Meng, D.; Chang, W.; Li, R.; Yin, H.; Liang, Y. Flavonoid Biosynthesis Is Likely More Susceptible to Elevation and Tree Age Than Other Branch Pathways Involved in Phenylpropanoid Biosynthesis in Ginkgo Leaves. Front. Plant Sci. 2019, 10, 983. [Google Scholar] [CrossRef]
- Yang, H.W.; Chai, Q.; Yin, W.; Hu, F.L.; Qin, A.Z.; Fan, Z.L.; Yu, A.Z.; Zhao, C.; Fan, H. Yield photosynthesis and leaf anatomy of maize in inter and mono-cropping systems at varying plant densities. Crop J. 2022, 10, 893–903. [Google Scholar] [CrossRef]
- Wu, Y.S.; Gong, W.Z.; Wang, Y.M.; Yong, T.W.; Yang, F.; Liu, W.G.; Wu, X.L.; Du, J.B.; Shu, K.; Liu, J.; et al. Leaf area and photosynthesis of newly emerged trifoliolate leaves are regulated by mature leaves in soybean. J. Plant Res. 2018, 131, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Kamble, P.N.; Giri, S.P.; Mane, R.S.; Tiwana, A. Estimation of chlorophyll content in young and adult leaves of some selected plants. Univers. J. Environ. Res. Technol. 2015, 5, 306–310. [Google Scholar]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Wilfried, S.; Rachel, D.R.; Efraim, L. Biosynthesis of plant-derived flavor com pounds. Plant J. 2008, 54, 712–732. [Google Scholar]
- Chen, Y.; Zhu, C.; Zhao, Y.; Zhang, S.; Wang, W. Transcriptomics Integrated with Changes in Cell Wall Material of Chestnut (Castanea mollissima Blume) during Storage Provides a New Insight into the “Calcification” Process. Foods 2022, 11, 1136. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.X.; Chen, M.H.; Shen, J.G.; Zhang, Y.Z.; Yuan, J.P. Study on components of haematochrome in Wild Zingiber strioatum from Mount Fanjing. Med. Plant 2012, 5, 62–64. [Google Scholar]
- Zhang, H.Q.; Liu, X.W.; Song, B.T.; Nie, B.H.; Zhang, W.; Zhao, Z.Q. Effect of excessive nitrogen on levels of amino acids and sugars, and differential response to postharvest cold storage in potato (Solanum tuberosum L.) tubers. Plant Physiol. Biochem. 2020, 157, 38–46. [Google Scholar] [CrossRef]
- Rivero Meza, S.L.; de Castro Tobaruela, E.; Benedetti Pascoal, G.; Louro Massaretto, I.; Purgatto, E. Post-harvest treatment with methyl jasmonate impacts lipid metabolism in tomato pericarp (Solanum lycopersicum L. cv. Grape) at different ripening stages. Foods 2021, 10, 877. [Google Scholar] [CrossRef]
- Gokavi, N.; Mote, K.; Jayakumar, M.; Raghuramulu, Y.; Surendran, U. The effect of modified pruning and planting systems on growth, yield, labour use efficiency and economics of Arabica coffee. Sci. Hortic. 2021, 276, 109764. [Google Scholar] [CrossRef]
- Beidler, K.V.; Taylor, B.N.; Strand, A.E.; Cooper, E.R.; Schönholz, M.; Pritchard, S.G. Changes in root architecture under elevated concentrations of CO2 and nitrogen reflect alternate soil exploration strategies. New Phytol. 2015, 205, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Baoyin, T.; Sun, J.; Minggagud, H.; Li, X. Plant sizes mediate mowing induced changes in nutrient stoichiometry and allocation of a perennial grass insemiarid grassland. Ecol. Evol. 2018, 8, 3109–3118. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Minggagud, H.; Baoyin, T.; Li, F.Y. Plant production decreases whereas nutrients concentration increases in response to the decrease of mowing stubble height. J. Environ. Manag. 2020, 253, 109745. [Google Scholar] [CrossRef]
- Yuan, J.H.; Li, H.Y.; Yang, Y.F. The compensatory tillering in the forage grass hordeum brevisubulatum after simulated grazing of different severity. Front. Plant Sci. 2020, 11, 792. [Google Scholar] [CrossRef]
- Naliwajski, M.R.; Sklodowska, M. The relationship between carbon and nitrogen metabolism in cucumber leaves acclimated to salt stress. Peer J. 2018, 6, 6043. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Photosynthetic Indexes | CK | T1 | T2 | |
|---|---|---|---|---|
| Photosynthetic Parameters | Pn (μmol/m2s) | 4.83 ± 0.25 c | 7.05 ± 0.26 a | 5.93 ± 0.22 b |
| Gs (mmol/m2s) | 0.041 ± 0.005 c | 0.049 ± 0.004 a | 0.045 ± 0.005 b | |
| Ci (μmolCO2/mol) | 175.79 ± 16.32 a | 137.59 ± 11.73 c | 168.84 ± 20.17 b | |
| Tr (mmol/m2s) | 0.71 ± 0.05 c | 0.78 ± 0.04 a | 0.73 ± 0.07 b | |
| Ewu (μmol/mmol) | 0.68 ± 0.05 b | 0.76 ± 0.04 a | 0.71 ± 0.07 a | |
| EC (mol/m2s) | 0.03 ± 0.003 b | 0.05 ± 0.005 a | 0.04 ± 0.007 ab | |
| Ls (%) | 0.56 ± 0.04 a | 0.65 ± 0.02 a | 0.57 ± 0.05 a | |
| Chlorophyll fluorescence parameters | Fv/Fm | 0.72 ± 0.11 b | 0.78 ± 0.01 a | 0.76 ± 0.01 a |
| Φ(II) | 0.42 ± 0.03 a | 0.42 ± 0.02 a | 0.36 ± 0.02 a | |
| NPQ | 0.89 ± 0.15 b | 1.37 ± 0.08 a | 1.77 ± 0.14 a | |
| qP | 0.74 ± 0.02 a | 0.7 ± 0.048 b | 0.68 ± 0.03 b | |
| YNPQ | 0.27 ± 0.03 b | 0.34 ± 0.02 ab | 0.41 ± 0.03 a | |
| YNO | 0.32 ± 0.02 a | 0.25 ± 0.006 b | 0.23 ± 0.002 b | |
| ETR | 24.62 ± 1.83 a | 24.67 ± 1.4 a | 21.32 ± 1.7 a | |
| Photosynthetic Pigments | CK | T1 | T2 |
|---|---|---|---|
| Chl a (mg/g) | 1.23 ± 0.31 b | 1.06 ± 0.64 b | 2.09 ± 0.05 a |
| Chl b (mg/g) | 0.65 ± 0.08 b | 0.61 ± 0.27 b | 1.03 ± 0.02 a |
| Car (mg/g) | 0.16 ± 0.05 b | 0.14 ± 0.05 b | 0.22 ± 0.003 a |
| Chl (mg/g) | 1.88 ± 0.29 b | 1.67 ± 0.91 b | 3.14 ± 0.07 a |
| Chl a/b | 1.87 ± 0.27 b | 1.63 ± 0.32 b | 2.01 ± 0.01 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Xie, S.; Du, D.; Wei, H.; Zhou, W.; Zhang, Y.; Tang, Z.; Li, D.; Liu, Y. Effect of Pruning Treatment on Growth Characteristics and Metabolites in Eucommia ulmoides Oliver (E. ulmoides). Forests 2023, 14, 2439. https://doi.org/10.3390/f14122439
Yang J, Xie S, Du D, Wei H, Zhou W, Zhang Y, Tang Z, Li D, Liu Y. Effect of Pruning Treatment on Growth Characteristics and Metabolites in Eucommia ulmoides Oliver (E. ulmoides). Forests. 2023; 14(12):2439. https://doi.org/10.3390/f14122439
Chicago/Turabian StyleYang, Jing, Shengnan Xie, Dandan Du, Hongling Wei, Wenling Zhou, Ying Zhang, Zhonghua Tang, Dewen Li, and Ying Liu. 2023. "Effect of Pruning Treatment on Growth Characteristics and Metabolites in Eucommia ulmoides Oliver (E. ulmoides)" Forests 14, no. 12: 2439. https://doi.org/10.3390/f14122439
APA StyleYang, J., Xie, S., Du, D., Wei, H., Zhou, W., Zhang, Y., Tang, Z., Li, D., & Liu, Y. (2023). Effect of Pruning Treatment on Growth Characteristics and Metabolites in Eucommia ulmoides Oliver (E. ulmoides). Forests, 14(12), 2439. https://doi.org/10.3390/f14122439