
Evaluation of Fungicides as Protective and Curative Treatments against Canker Disease of Eucalyptus urograndis Caused by Chrysoporthe deuterocubensis in Malaysia

 , ,
, ,
Abstract
:1. Introduction
2. Methods
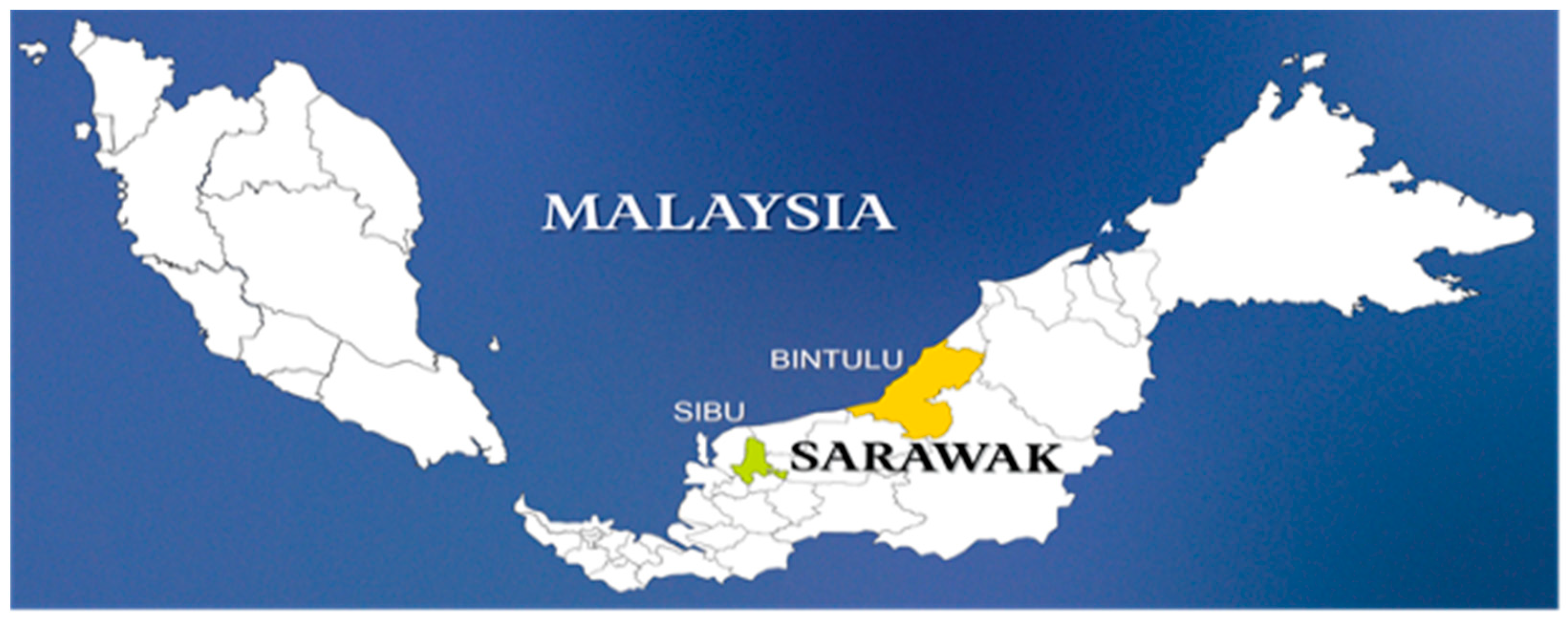
2.1. Disease Symptoms and Sample Collection
2.2. Identification of Isolated Fungal Pathogens
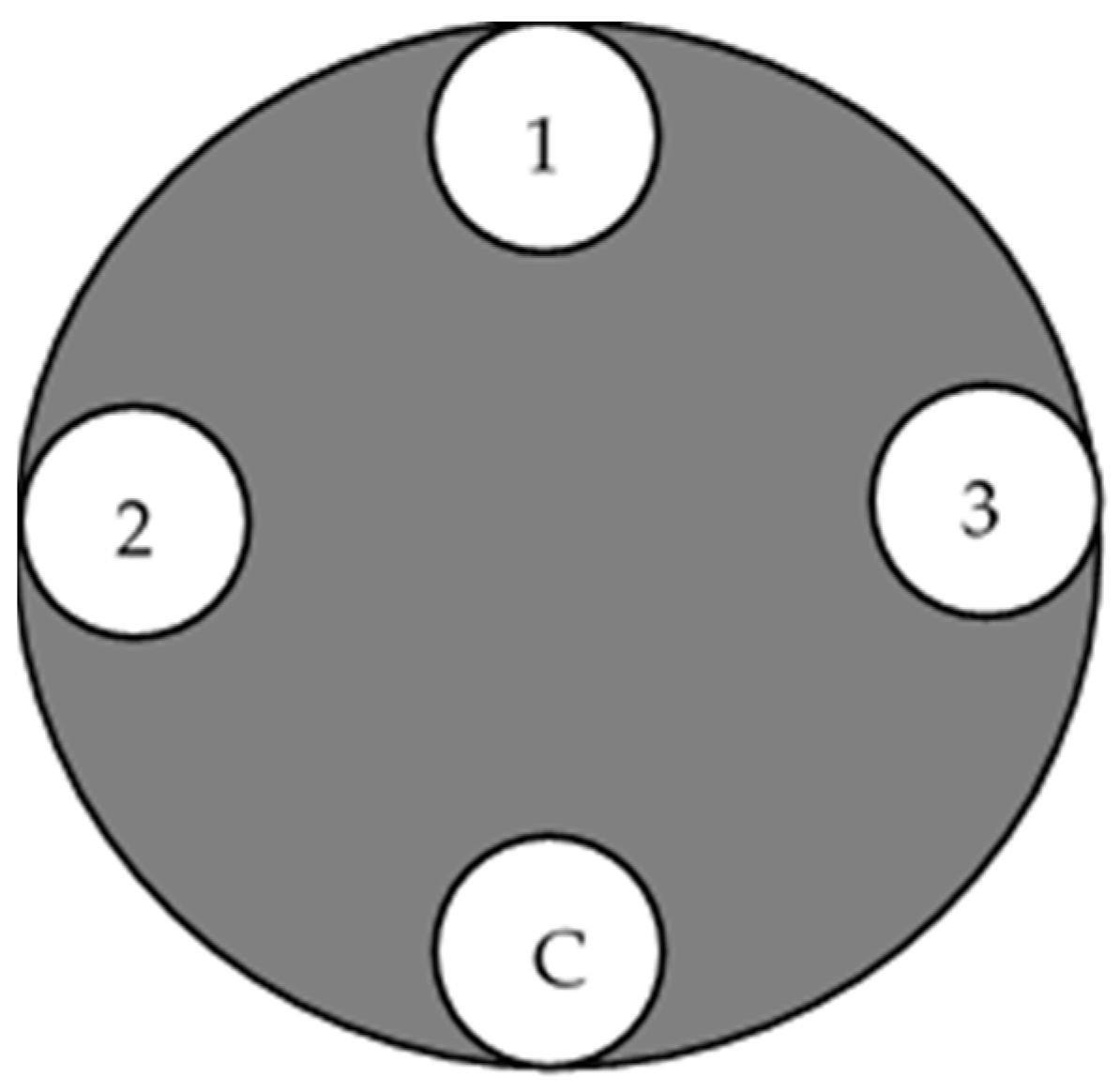
2.3. Pathogenicity Test
2.4. DNA Extraction, PCR, Sequencing, and Phylogenetic Analysis
2.5. In Vitro Evaluation of Chemical Fungicides against Chrysoporthe deuterocubensis
2.6. Field Efficiency of Fungicides Applied for Preventive and Curative Activity against Chrysoporthe deuterocubensis
2.7. Statistical Analysis
3. Results
3.1. Collection of Fungal Isolates and Their General Morphological Characterisation
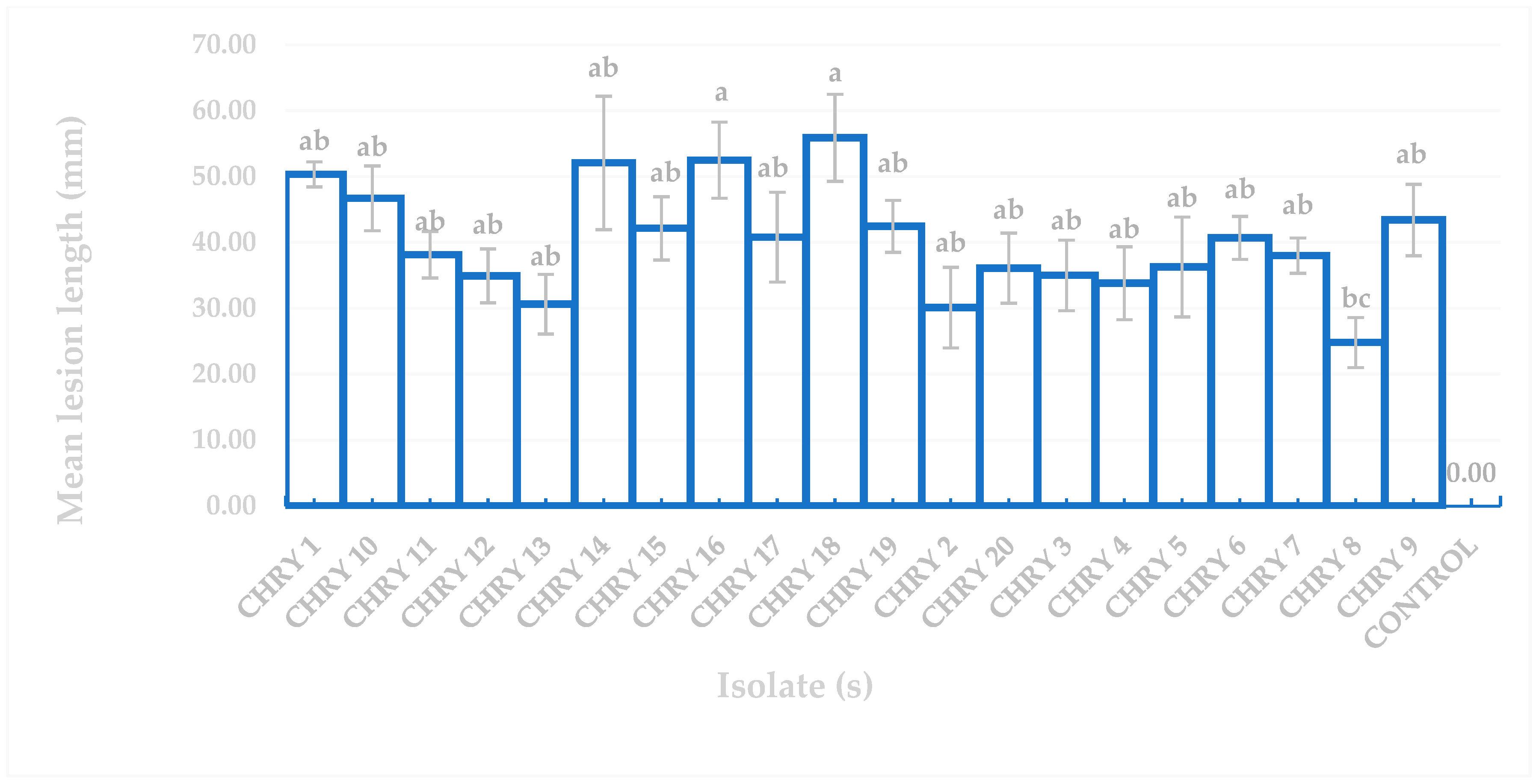
3.2. Pathogenicity Test on Eucalyptus urograndis
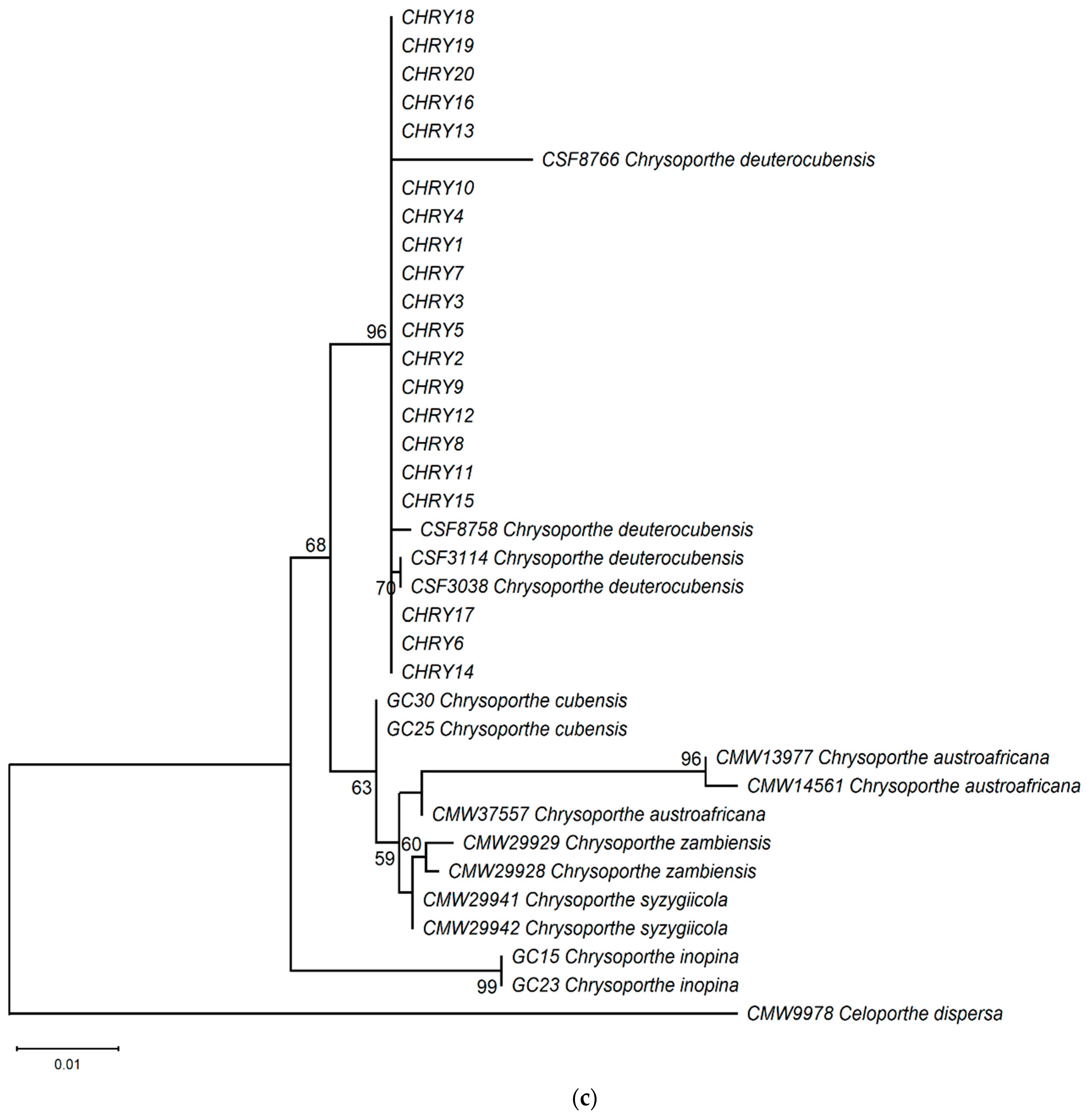
3.3. Molecular Phylogenetic Analyses
3.4. In Vitro Evaluation of Chemical Fungicides against Chrysorpothe deuterocubensis
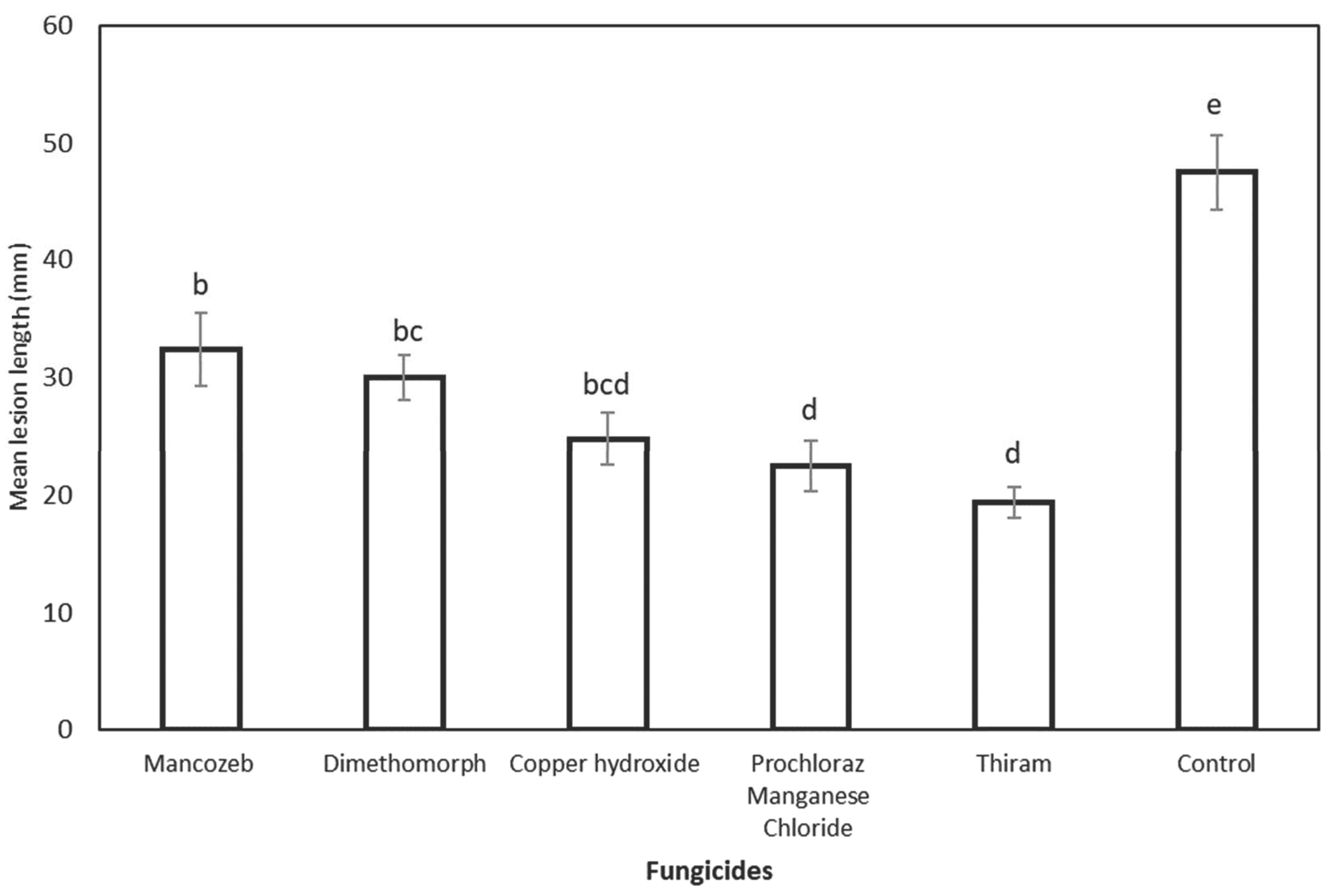
3.5. Field Efficiency of Fungicides Applied for Preventive and Curative Activity against Chrysoporthe deuterocubensis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Science, E.; Hamid, H.A.; Abiri, R. The way forward of Forest Plantation in Malaysia. IOP Conf. Ser. Earth Environ. Sci. 2022, 18, 959. [Google Scholar]
- Ratnasingam, J.; Latib, H.A.; Paramjothy, N.; Liat, L.C.; Nadarajah, M.; Ioras, F. Plantation Forestry in Malaysia: An Evaluation of its Successes and Failures Since the 1970. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 1789–1801. [Google Scholar] [CrossRef]
- Ambrose, A.; Liam, J.; Terhem, R. New and Emerging Disease Threats to Forest Plantations in Sarawak Borneo, Malaysia. In Current and Emerging Challenges in the Diseases of Trees; Bellé, D.C., Ed.; IntechOpen: Rijeka, Croatia, 2022; p. 112. [Google Scholar] [CrossRef]
- Tarigan, M.; Roux, J.; Wyk, M.V.; Tjahjono, B.; Wingfield, M.J. A new wilt and die-back disease of Acacia mangium associated with A new wilt and die-back disease of Acacia mangium associated with Ceratocystis manginecans and C. acaciivora sp. nov. in Indonesia. S. Afr. J. Bot. 2011, 77, 292–304. [Google Scholar] [CrossRef]
- Ss, L.; Lee, S.S. Observations on the successes and failures of acacia plantations in sabah and sarawak and the way forward. J. Trop. For. Sci. 2018, 30, 468–475. [Google Scholar]
- Zhang, Y.X.; Wang, X.J. Geographical spatial distribution and productivity dynamic change of eucalyptus plantations in China. Sci. Rep. 2021, 11, 19764. [Google Scholar] [CrossRef]
- Naidoo, S.; Külheim, C.; Zwart, L.; Mangwanda, R.; Oates, C.N.; Visser, E.A.; Wilken, F.E.; Mamni, T.B.; Myburg, A.A. Uncovering the defence responses of eucalyptus to pests and pathogens in the genomics age. Tree Physiol. 2014, 34, 931–943. [Google Scholar] [CrossRef]
- Zuhaidi Yahya, A. Planting of Eucalyptus in Malaysia. Acta Sci. Agric. 2020, 4, 139–140. [Google Scholar] [CrossRef]
- De Oliveira Castro, C.A.; dos Santos, G.A.; Takahashi, E.K.; Pires Nunes, A.C.; Souza, G.A.; de Resende, M.D.V. Accelerating Eucalyptus breeding strategies through top grafting applied to young seedlings. Ind. Crops Prod. 2021, 171, 113906. [Google Scholar] [CrossRef]
- Colodette, J.L.; Gomes, C.M.; Gomes, F.J.; Cabral, C.P. The Brazilian wood biomass supply and utilisation focusing on eucalypt. Chem. Biol. Technol. Agric. 2014, 1, 25. [Google Scholar] [CrossRef]
- Rezende, G.D.S.P.; de Resende, M.D.V.; de Assis, T.F. Eucalyptus Breeding for Clonal Forestry. In Challenges and Opportunities for the World’s Forests in the 21st Century; Springer: Berlin/Heidelberg, Germany, 2014; pp. 393–424. [Google Scholar]
- Hunter, G.C.; Crous, P.W.; Carnegie, A.J.; Burgess, T.I.; Wingfield, M.J. Mycosphaerella and Teratosphaeria diseases of Eucalyptus; easily confused and with serious consequences. Fungal Divers. 2011, 50, 145–166. [Google Scholar] [CrossRef]
- Wingfield, M.J. Pathology | Disease Affecting Exotic Plantation Species. Encycl. For. Sci. 2004, 816–822. Available online: https://www.sciencedirect.com/science/article/pii/B0121451607000624 (accessed on 12 July 2023).
- Silva, X.; Asiegbu, F.O. Chapter 15—Eucalyptus fungal diseases. In Forest Microbiology; Asiegbu, F.O., Kovalchuk, A., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 313–337. [Google Scholar]
- Chen, S.; Van Wyk, M.; Roux, J.; Wingfield, M.J.; Xie, Y.; Zhou, X. Taxonomy and pathogenicity of Ceratocystis species on Eucalyptus trees in South China, including C. chinaeucensis sp. nov. Fungal Divers. 2013, 58, 267–279. [Google Scholar] [CrossRef]
- Roux, J.; Wingfield, M.J.; Fourie, A.; Noeth, K.; Barnes, I. Ceratocystis wilt on Eucalyptus: First record from South Africa. South. For. J. For. Sci. 2020, 82, 24–31. [Google Scholar] [CrossRef]
- Yong, W.T.L.; Ades, P.K.; Runa, F.A.; Bossinger, G.; Sandhu, K.S.; Potts, B.M.; Tibbits, J.F.G. Genome-wide association study of myrtle rust (Austropuccinia psidii) resistance in Eucalyptus obliqua (subgenus Eucalyptus). Tree Genet. Genomes 2021, 17, 31. [Google Scholar] [CrossRef]
- Carnegie, A.J. First Report of Puccinia psidii (Myrtle Rust) in Eucalyptus Plantations in Australia. Plant Dis. 2015, 99, 161. [Google Scholar] [CrossRef] [PubMed]
- Gryzenhout, M.; Myburg, H.; Van Der Merwe, N.A.; Wingfield, B.D.; Wingfield, M.J. Chrysoporthe, a new genus to accommodate Cryphonectria cubensis. Stud. Mycol. 2004, 50, 119–142. [Google Scholar]
- Pegg, G.S.; Gryzenhout, M.; O’Dwyer, C.; Drenth, A.; Wingfield, M.J. The Eucalyptus canker pathogen Chrysoporthe cubensis discovered in eastern Australia. Australas. Plant Pathol. 2010, 39, 343–349. [Google Scholar] [CrossRef]
- Rauf, M.R.B.A.; McTaggart, A.R.; Marincowitz, S.; Barnes, I.; Japarudin, Y.; Wingfield, M.J. Pathogenicity of Chrysoporthe deuterocubensis and Myrtoporthe bodenii gen. et sp. nov. on Eucalyptus in Sabah, Malaysia. Australas. Plant Pathol. 2020, 49, 53–64. [Google Scholar] [CrossRef]
- Li, G.; Chen, W.; Jie, F.; Chen, S. Selection of tolerant Eucalyptus genotypes to Botryosphaeriaceae species in southern China. J. Plant Pathol. 2022, 104, 527–535. [Google Scholar] [CrossRef]
- Chen, S.F.; Gryzenhout, M.; Roux, J.; Xie, Y.J.; Wingfield, M.J.; Zhou, X.D. Identification and pathogenicity of Chrysoporthe cubensis on Eucalyptus and Syzygium spp. in South China. Plant Dis. 2010, 94, 1143–1150. [Google Scholar] [CrossRef]
- Soares, T.P.F.; Ferreira, M.A.; Mafia, R.G.; Oliveira, L.S.S.; Hodges, C.S.; Alfenas, A.C. Canker disease caused by Chrysoporthe doradensis and C. cubensis on Eucalyptus sp. and Tibouchina spp. in Brazil. Trop. Plant Pathol. 2018, 43, 314–322. [Google Scholar] [CrossRef]
- Awing, N.H.; Ambrose, A.; Abdu, A.; Hassan, A.; Terhem, R. Characterisation of Chrysoporthe cubensis and Chrysoporthe deuterocubensis, the Stem Canker Diseases of Eucalyptus spp. in a Forest Plantation in Malaysia. Forests 2023, 14, 1660. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Rokas, A.; Charlesworth, D.; Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics. By M. Nei and S. Kumar. Oxford University Press. 2000. ISBN: 0-19-513584-9 (hbk); 0-19-513585-7 (pbk). xiv+ 333 pages. Price: £ 65 (hbk); £ 32.50 (pbk). Genet. Res. 2001, 77, 117–120. [Google Scholar] [CrossRef]
- Malipatil, R.; Yenjerappa, S.T.; Amaresh, Y.S.; Sreedevi, S.C.; Jaiprakash Narayan, R.P. Efficacy of different fungicides by in vitro against Colletotrichum gloeosporioides, the causal agent of mango anthracnose. Int. J. Chem. Stud. 2021, 9, 3408–3412. [Google Scholar] [CrossRef]
- Musdalifa, A.A.; Rosmana, A. The response of different fungicides against Lasiodiplodia pseudotheobromae causing dieback disease of cocoa through in vitro test. IOP Conf. Ser. Earth Environ. Sci. 2021, 807, 22091. [Google Scholar] [CrossRef]
- Ullah, S. In Vitro evaluation of commonly available fungicides against three fungal isolates. Plant Pathol. Q. 2018, 8, 67–77. [Google Scholar] [CrossRef]
- Dahal, N.; Shrestha, R.K. Evaluation of Efficacy of Fungicides Against Fusarium oxysporum f. sp. lentis In Vitro at Lamjung, Nepal. J. Inst. Agric. Anim. Sci. 2018, 35, 105–112. [Google Scholar] [CrossRef]
- Admasu, W.; Sintayehu, A.; Gezahgne, A.; Terefework, Z. In Vitro bioefficacy of Trichoderma species against two Botryosphaeriaceae fungi causing Eucalyptus stem canker disease in Ethiopia. J. Nat. Pestic. Res. 2023, 4, 100037. [Google Scholar] [CrossRef]
- Bester, W.; Crous, P.W.; Fourie, P.H. Evaluation of fungicides as potential grapevine pruning wound protectants against Botryosphaeria species. Australas. Plant Pathol. 2007, 36, 73–77. [Google Scholar] [CrossRef]
- Halleen, F.; Fourie, P.H.; Crous, P.W. Control of black foot disease in grapevine nurseries. Plant Pathol. 2007, 56, 637–645. [Google Scholar] [CrossRef]
- Yuli, E.; Dayou, J.; Chong, K.P. In Vitro Antifungal Activity of Thiram against Ganoderma boninense. Trans. Sci. Technol. 2020, 7, 159–164. [Google Scholar]
- Pohanish, R.P.T. Sittig’s Handbook of Pesticides and Agricultural Chemicals, 2nd ed.; William Andrew Publishing: Oxford, UK, 2015; pp. 769–838. [Google Scholar]
- Picone, A.L.; Rizzato, M.L.; Lusi, A.R.; Romano, R.M. Stamplike flexible SERS substrate for in-situ rapid detection of thiram residues in fruits and vegetables. Food Chem. 2022, 373, 131570. [Google Scholar] [CrossRef]
- Kaushik, P.; Shakil, N.A.; Kumar, J.; Singh, M.K.; Singh, M.K.; Yadav, S.K. Development of controlled release formulations of thiram employing amphiphilic polymers and their bioefficacy evaluation in seed quality enhancement studies. J. Environ. Sci. Health Part B 2013, 48, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Belcher, E.W.; Waldrip, B.T. Effect of thiram on seed mold and germination of slash pine seed. Proc. Assoc. Off. Seed Anal. 1972, 62, 91–93. [Google Scholar]
- Ivić, D. Curative and Eradicative Effects of Fungicides. In Fungicides; IntechOpen: London, UK, 2010. [Google Scholar]
- Gullino, M.L.; Tinivella, F.; Garibaldi, A.; Kemmitt, G.M.; Bacci, L.; Sheppard, B. History and Role of Mancozeb in Disease Management. Plant Dis. 1963, 94, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Wang, P.; Pu, Z.; Lu, L.; Chen, G.; Hu, X.; Fayyaz, A.; Gai, Y. Effects of mancozeb on citrus rhizosphere bacterial community. Microb. Pathog. 2021, 154, 104845. [Google Scholar] [CrossRef]
- Vinicius, M.; Bucker, W.; Luiz, E.; Masson, M.V.; Moraes, W.B.; Furtado, E.L. Chemical Control of Eucalyptus Rust: Brazilian Experiences. In Fungicides—Showcases of Integrated Plant Disease Management from Around the World; InTech: London, UK, 2013. [Google Scholar]
- Crous, P.W.; Wingfield, M.J.; Denman, S.; Sadie, A.; Crous, P.W.; Sadie, A.; Wingfield, M.J. Evaluation of fungicides for the control of Botryosphaeria protearum on Protea magnifica in the Western Cape Province of South Africa. Australas. Plant Pathol. 2004, 33, 97. [Google Scholar]
- Tran, T.T.T.; Pham, T.Q.; Barber, P.A.; Nguyen, C.M. Control of Ceratocystis manginecans causing wilt disease on Acacia mangium seedlings. Australas. Plant Pathol. 2018, 47, 579–586. [Google Scholar] [CrossRef]
- Weber, R.W.S.; Børve, J. Infection biology as the basis of integrated control of apple canker (Neonectria ditissima) in Northern Europe. CABI Agric. Biosci. 2021, 2, 5. [Google Scholar] [CrossRef]
- Narciso, J.A.; Ference, C.M.; Ritenour, M.A.; Widmer, W.W. Effect of copper hydroxide sprays for citrus canker control on wild-type Escherichia coli. Lett. Appl. Microbiol. 2012, 54, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Kleczewski, N.M.; Butts-Willmsmeyer, C.; Scanlan, C. Assessing the Curative and Protective Impacts of Select Fungicides for Control of Powdery Mildew of Wheat. Plant Dis. 2020, 104, 1195–1200. [Google Scholar] [CrossRef] [PubMed]
- Prospero, S.; Botella, L.; Santini, A.; Robin, C. Biological control of emerging forest diseases: How can we move from dreams to reality? For. Ecol. Manag. 2021, 496, 119377. [Google Scholar] [CrossRef]
- Rolando, C.A.; Dick, M.A.; Gardner, J.; Bader, M.K.F.; Williams, N.M. Chemical control of two Phytophthora species infecting the canopy of Monterey pine (Pinus radiata). For. Pathol. 2017, 47, e12327. [Google Scholar] [CrossRef]
- Khuzaimah, Z.; Nawi, N.M.; Adam, S.N.; Kalantar, B.; Emeka, O.J.; Ueda, N. Application and Potential of Drone Technology in Oil Palm Plantation: Potential and Limitations. J. Sens. 2022, 2022, 5385505. [Google Scholar] [CrossRef]
- Chi, N.M.; Thu, P.Q.; Nam, H.B.; Quang, D.Q.; Phong, L.V.; Van, N.D.; Trang, T.T.; Kien, T.T.; Tam, T.T.T.; Dell, B. Management of Phytophthora palmivora disease in Citrus reticulata with chemical fungicides. J. Gen. Plant Pathol. 2020, 86, 494–502. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species | Region | GPS location | Plant Age (Years) | Rotation | Spacing (m) |
|---|---|---|---|---|---|---|
| 1 | Eucalyptus urograndis | Sibu | 2°32′54.60″ N, 112°29′59.97″ E | 7 | 1 | 3 × 3 |
| 2 | Eucalyptus urograndis | Bintulu | 2°27′55.188″ N 112°27′18.396″ E | 7 | 1 | 3 × 3 |
| Isolate Identity | GenBank Accession Number | |
|---|---|---|
| Internal Transcribed Spacer (ITS) | β-tubulin 2(BT2) | |
| CHRY1 | OR234820 | OR262469 |
| CHRY2 | OR234821 | OR262470 |
| CHRY3 | OR234822 | OR262471 |
| CHRY4 | OR234823 | OR262472 |
| CHRY5 | OR234824 | OR262473 |
| CHRY6 | OR234825 | OR262474 |
| CHRY7 | OR234826 | OR262475 |
| CHRY8 | OR234827 | OR262476 |
| CHRY9 | OR234828 | OR262477 |
| CHRY10 | OR234829 | OR262478 |
| CHRY11 | OR234830 | OR262479 |
| CHRY12 | OR234831 | OR262480 |
| CHRY13 | OR234832 | OR262481 |
| CHRY14 | OR234833 | OR262482 |
| CHRY15 | OR234834 | OR262483 |
| CHRY16 | OR234835 | OR262484 |
| CHRY17 | OR234836 | OR262485 |
| CHRY18 | OR234837 | OR262486 |
| CHRY19 | OR234838 | OR262487 |
| CHRY20 | OR234839 | OR262488 |
| Common Name | Chemical Group | Formulation |
|---|---|---|
| Dithiocarbamates | Thiram (Tetramethylthiuram disulfide) | 80% WP |
| Imidazole | Prochloraz manganese chloride | 50% WP |
| Inorganic | Copper hydroxide | 77% WP |
| Carboxylic acid amides | Dimethomorph | 50% wp |
| Dithiocarbamate | Mancozeb | 80% WP |
| Common Name | Chemical Group | Formulation (WP) | Concentration (g/L) * |
|---|---|---|---|
| Dithiocarbamates | Thiram | 80% | 2.0 |
| Imidazole | Prochloraz manganese chloride | 50% | 1.0 |
| Inorganic | Copper hydroxide | 77% | 2.2 |
| Carboxylic acid amides | Dimethomorph | 50% | 0.5 |
| dithiocarbamate | Mancozeb | 80% | 2.0 |
| Fungicide | Concentration (mg/mL) | ||||
|---|---|---|---|---|---|
| 0.125 | 0.25 | 0.50 | 1.00 | 2.00 | |
| RGI (%) | |||||
| Thiram | 0.0 b ± 0.0 | 0.0 c ± 0.0 | 0.0 c ± 0.0 | 24.5 c ± 8.8 | 42.3 c ± 12.8 |
| Prochloraz manganese chloride | 91.1 a ± 4.4 | 92.5 a ± 4.7 | 97.0 a ± 1.5 | 100.0 a ± 0.0 | 100.0 a ± 0.0 |
| Copper hydroxide | 0.0 b ± 0.0 | 0.0 c ± 0.0 | 0.0 c ± 0.0 | 0.0 d ± 0.0 | 5.7 d ± 11,6 |
| Dimethomorph | 3.2 b ± 9.7 | 18.8 b ± 22.1 | 28.6 b ± 11.1 | 31.9 b ± 15.7 | 57.6 b ± 14.8 |
| Mancozeb | 0.0 b ± 0.0 | 0.0 c ± 0.0 | 0.0 c ± 0.0 | 0.0 d ± 0.0 | 0.0 d ± 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ambrose, A.; Peter Remun, A.B.; Shamsul Kamar, N.A.; Mohamad Pazi, A.M.; Wan Azhar, W.M.A.; Awing, N.H.; Liam, J.; Terhem, R. Evaluation of Fungicides as Protective and Curative Treatments against Canker Disease of Eucalyptus urograndis Caused by Chrysoporthe deuterocubensis in Malaysia. Forests 2023, 14, 2337. https://doi.org/10.3390/f14122337
Ambrose A, Peter Remun AB, Shamsul Kamar NA, Mohamad Pazi AM, Wan Azhar WMA, Awing NH, Liam J, Terhem R. Evaluation of Fungicides as Protective and Curative Treatments against Canker Disease of Eucalyptus urograndis Caused by Chrysoporthe deuterocubensis in Malaysia. Forests. 2023; 14(12):2337. https://doi.org/10.3390/f14122337
Chicago/Turabian StyleAmbrose, Annya, Attlee Banyang Peter Remun, Nura Adilla Shamsul Kamar, Ahmad Mustapha Mohamad Pazi, Wan Muhammad Azrul Wan Azhar, Norida Hanim Awing, Jack Liam, and Razak Terhem. 2023. "Evaluation of Fungicides as Protective and Curative Treatments against Canker Disease of Eucalyptus urograndis Caused by Chrysoporthe deuterocubensis in Malaysia" Forests 14, no. 12: 2337. https://doi.org/10.3390/f14122337
APA StyleAmbrose, A., Peter Remun, A. B., Shamsul Kamar, N. A., Mohamad Pazi, A. M., Wan Azhar, W. M. A., Awing, N. H., Liam, J., & Terhem, R. (2023). Evaluation of Fungicides as Protective and Curative Treatments against Canker Disease of Eucalyptus urograndis Caused by Chrysoporthe deuterocubensis in Malaysia. Forests, 14(12), 2337. https://doi.org/10.3390/f14122337