Decadal Fire Effects on the Structure, Composition, Diversity, and Aboveground Carbon Stocks of a Neotropical Savanna

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
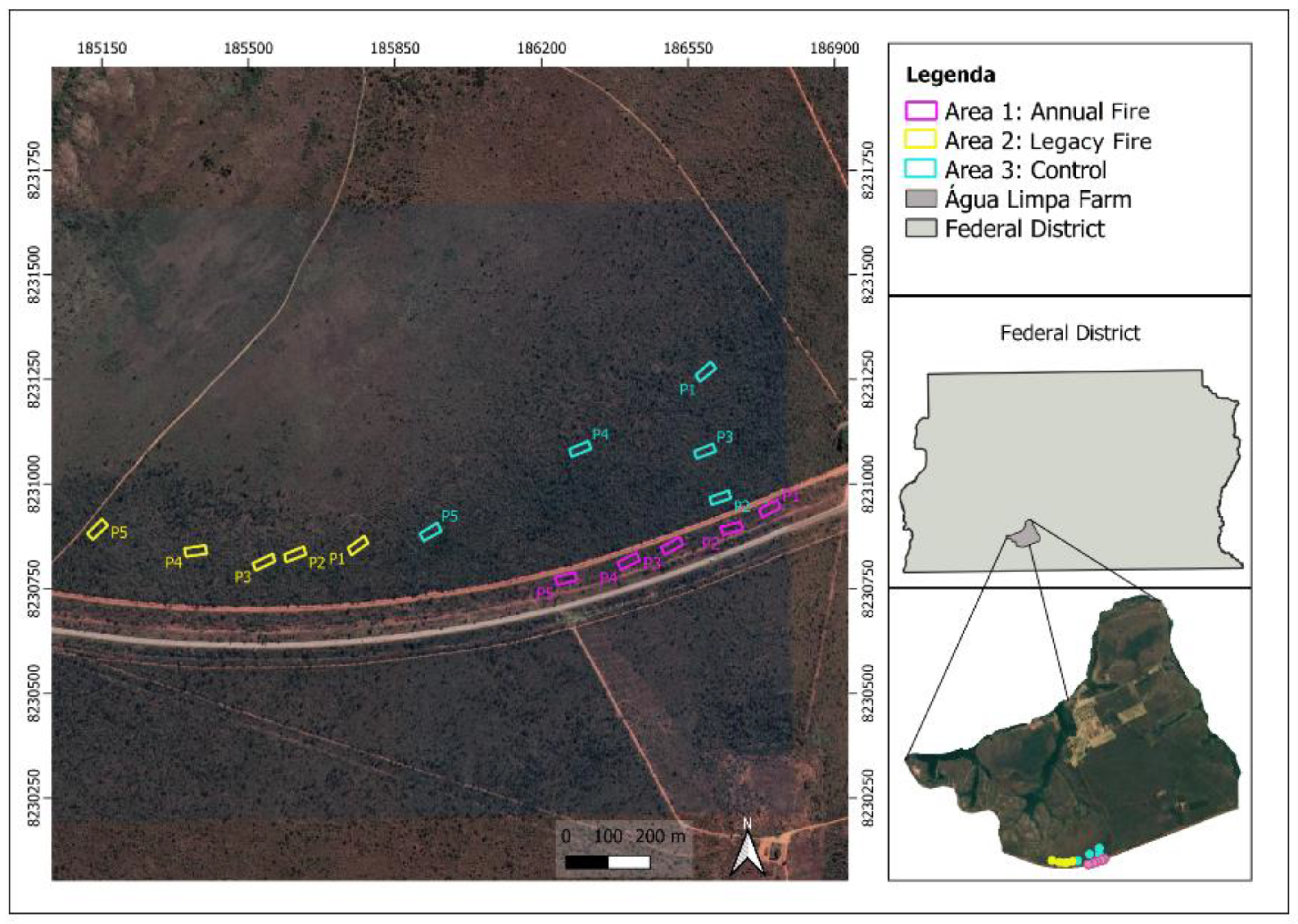
2.1. Study Site
2.2. Sampling Design
2.3. Fire Experiment
2.4. Vegetation Data Collection in the Field
2.5. Aboveground Carbon Stock Estimation
2.6. Statistical Analysis
2.6.1. Floristic Composition and Structure
2.6.2. Carbon Stocks
3. Results
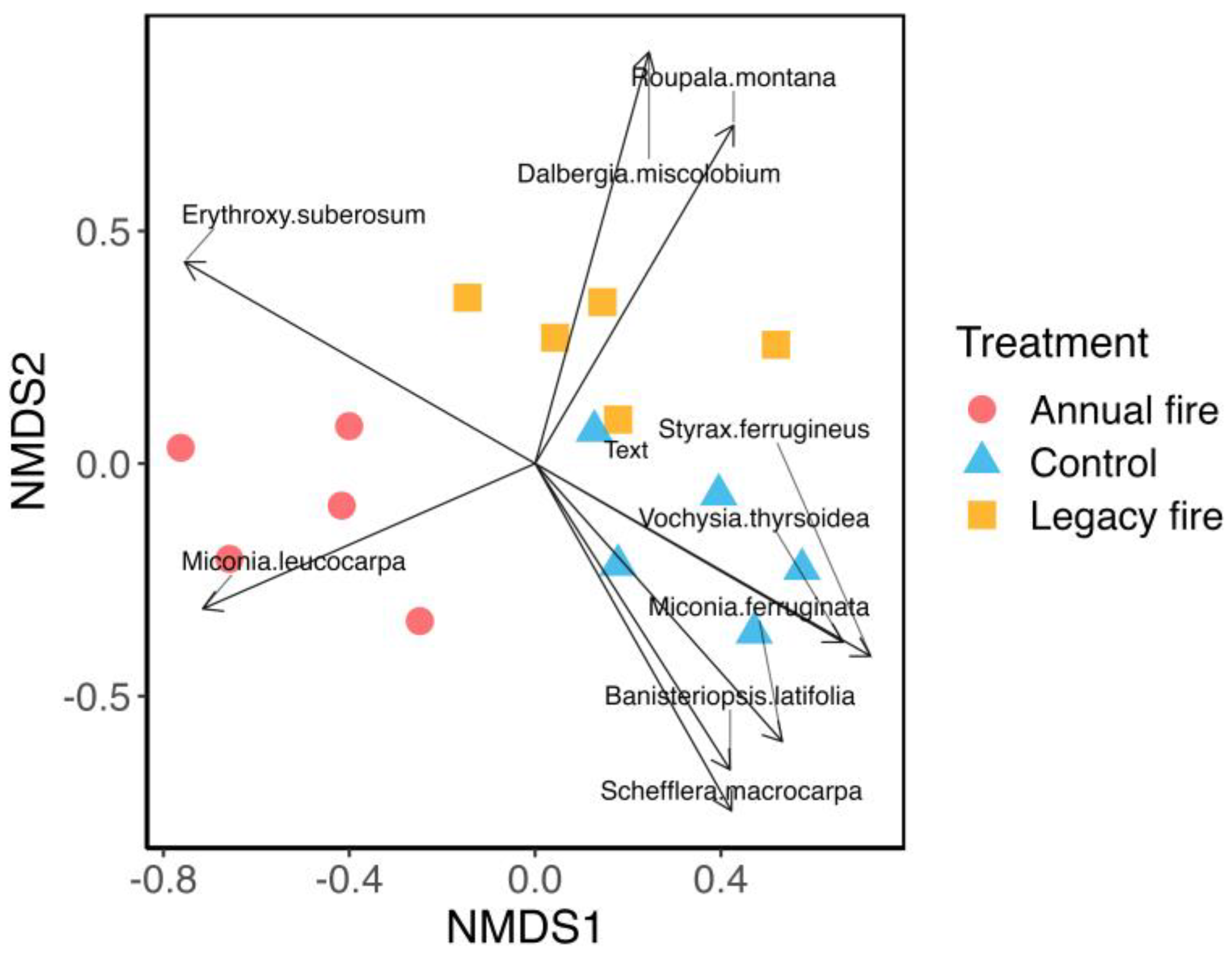
3.1. Species Composition across Treatments
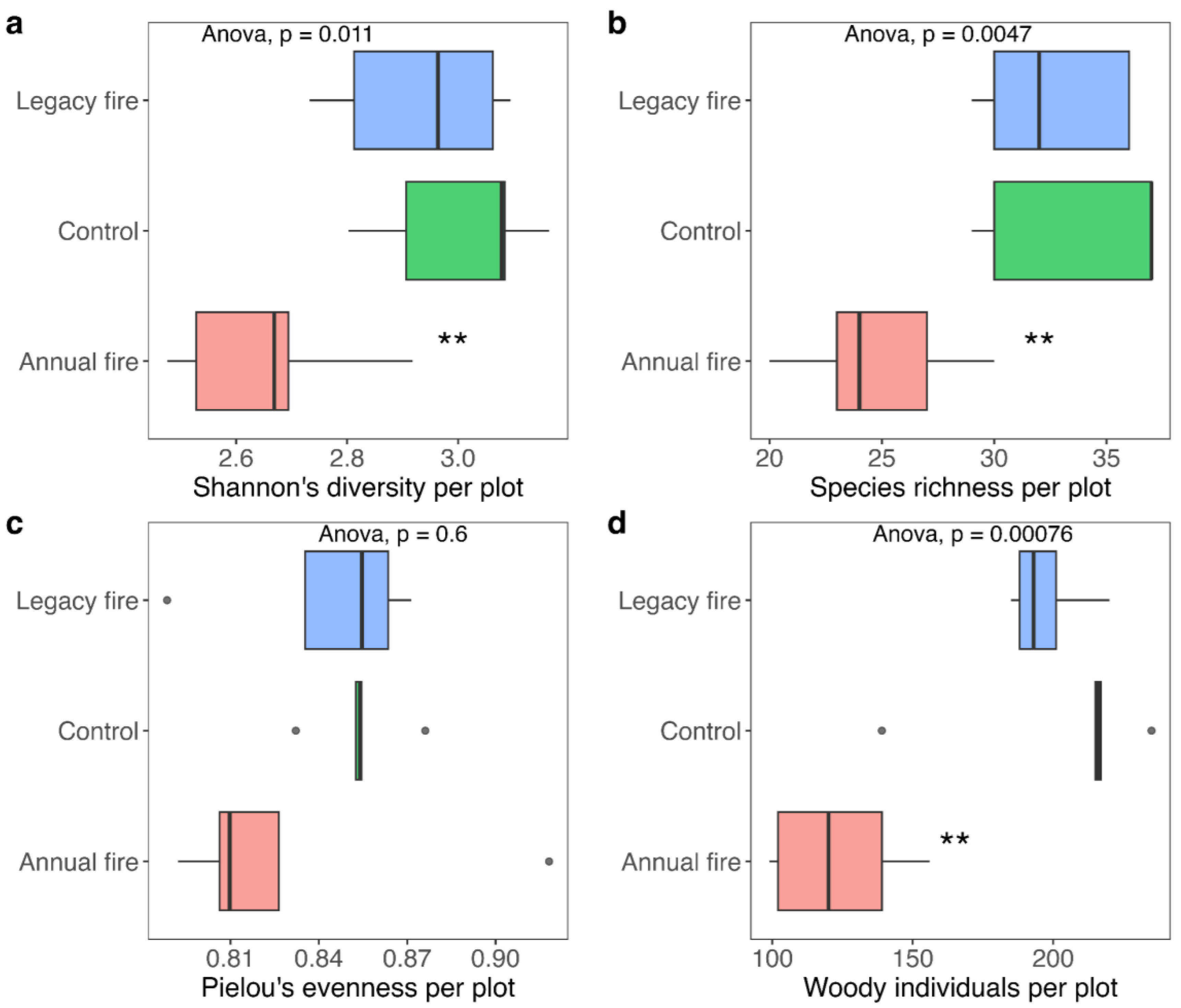
3.2. Diversity Metrics across Treatments
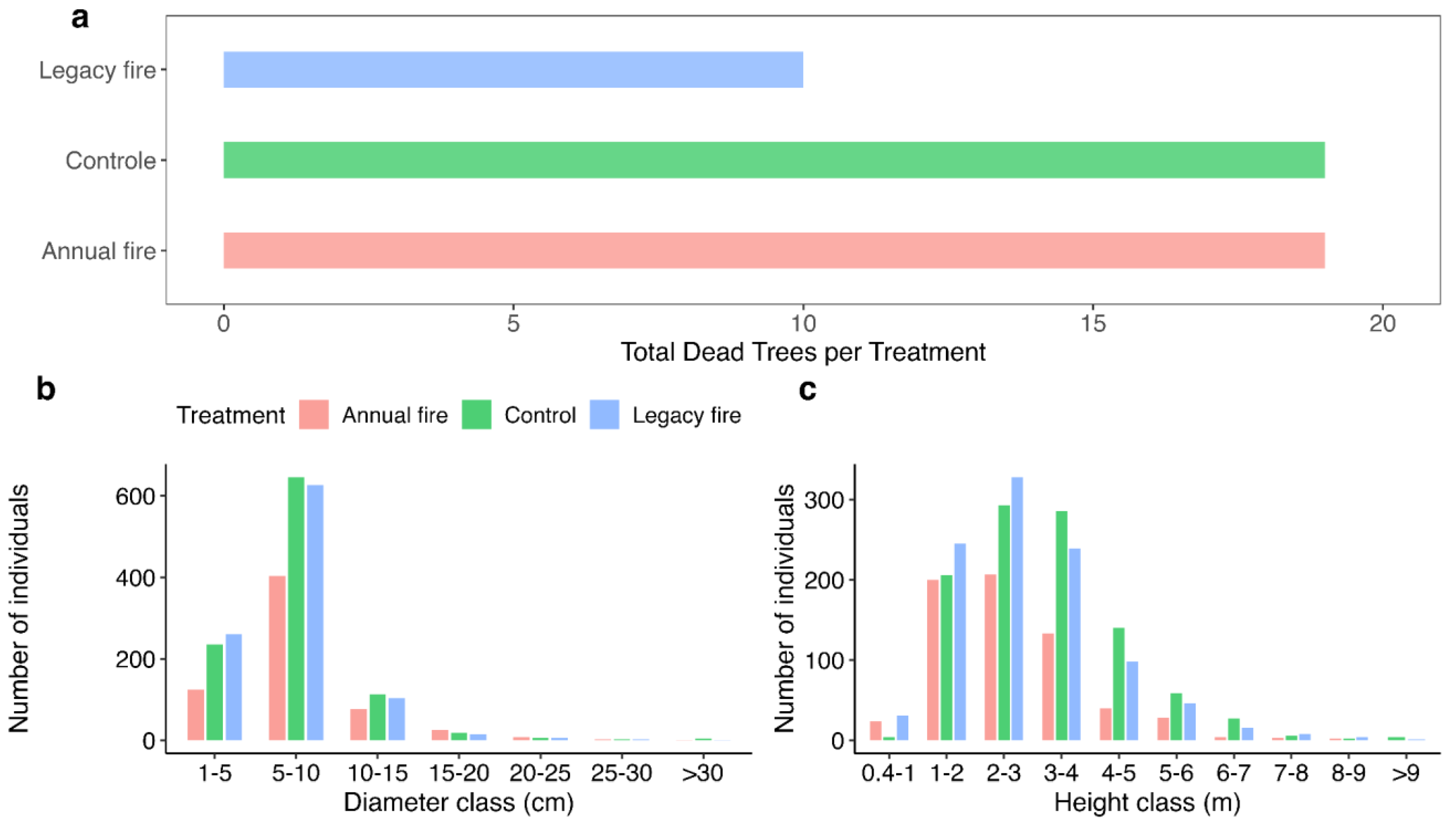
3.3. Structural Differences across Treatments
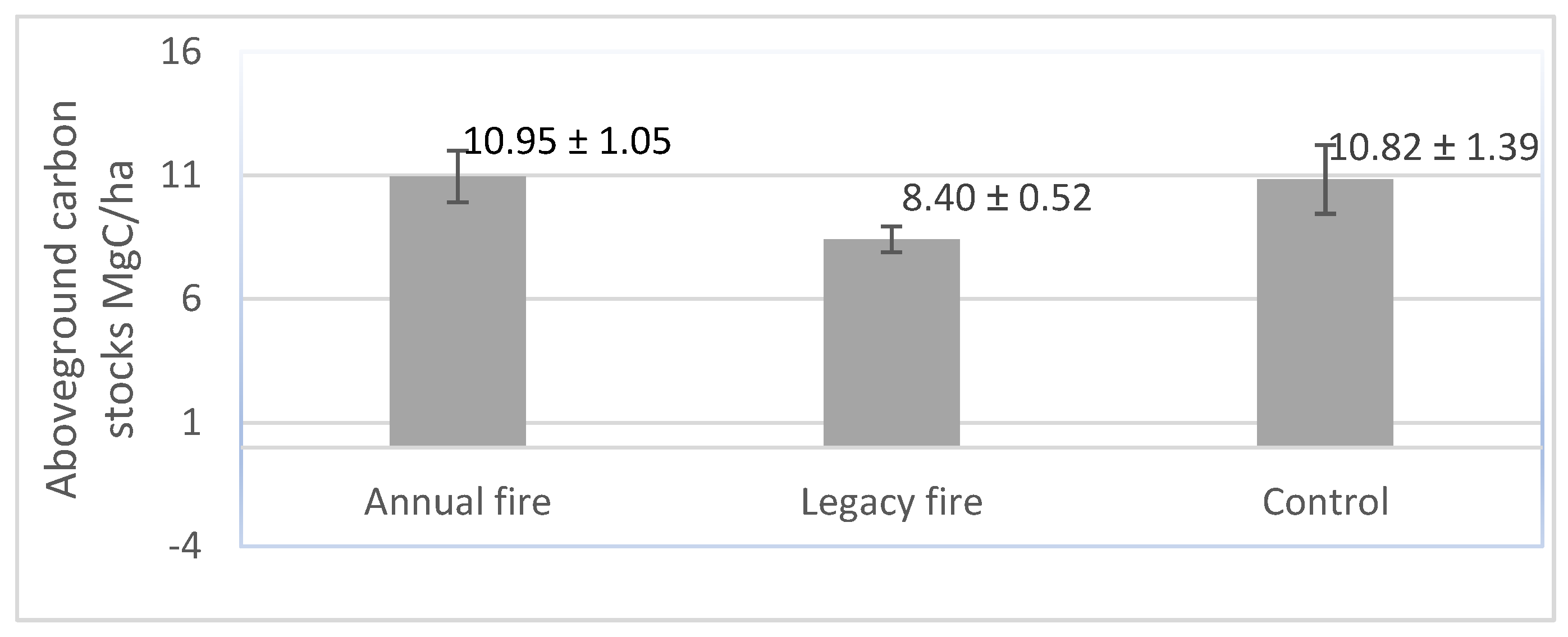
3.4. Aboveground Carbon Stocks across Treatments
4. Discussion
4.1. Relationship between Fire and Cerrado Species Composition
4.2. Fire Effect on Vegetation Diversity
4.3. Fire Effects on Vegetation Structure
4.4. Fire Effects on Aboveground Carbon Stocks across Treatments
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | NMDS1 | NMDS2 | R2 | p-Value |
|---|---|---|---|---|---|
| Agonandra brasiliensis Miers ex Benth. & Hook.f. | Opiliaceae | 0.54628 | 0.8376 | 0.2012 | 0.321 |
| Aspidosperm tomentosum Mart. | Apocynaceae | 0.90325 | 0.42912 | 0.1018 | 0.511 |
| Banisteriopsis latifolia (A.Juss.) B. Gates | Malpighiaceae | 0.53772 | −0.84312 | 0.6093 | 0.001 *** |
| Blepharocalyx salicifolius (Kunth) O. Berg. | Myrtaceae | −0.15560 | 0.98782 | 0.0870 | 0.598 |
| Byrsonima pachyphylla A. Juss. | Malpighiaceae | −0.86522 | −0.50139 | 0.1873 | 0.295 |
| Byrsonima verbascifolia (L.) DC. | Malpighiaceae | −0.91416 | −0.40534 | 0.0769 | 0.654 |
| Bowdichia virgilioides Kunth | Fabaceae | 0.29841 | 0.95444 | 0.1670 | 0.328 |
| Casearia sylvestris Sw | Salicaceae | −0.91062 | −0.41324 | 0.3848 | 0.052 |
| Caryocar brasiliense Cambess. | Caryocaraceae | 0.88254 | −0.47024 | 0.4325 | 0.014 * |
| Connarus suberosus Planch. | Connaraceae | −0.93377 | 0.35786 | 0.0402 | 0.779 |
| Dalbergia miscolobium Benth. | Mimosoideae | 0.26686 | 0.96374 | 0.8432 | 0.001 *** |
| Dalbergia elliptica | Fabaceae | 0.63236 | −0.77468 | 0.2087 | 0.275 |
| Davilla elliptica A.St.-Hil. | Dilleniaceae | −0.80026 | −0.59965 | 0.3734 | 0.035 * |
| Diospyros burchellii Hiern | Ebenaceae | −0.57408 | 0.8188 | 0.0911 | 0.676 |
| Diospyros lasiocalyx (Mart.) B.Walln. | Ebenaceae | 0.20319 | 0.97914 | 0.5188 | 0.014 * |
| Dimorphandra mollis Benth. | Fabaceae | 0.38512 | −0.92287 | 0.2701 | 0.056 |
| Enterolobium gummiferum (Mart.) J.F.Macr. | Fabaceae | 0.81475 | −0.57982 | 0.0684 | 0.691 |
| Eremanthus glomerulatus Less | Asteraceae | −0.43466 | −0.90059 | 0.2094 | 0.241 |
| Eriotheca pubescens (Mart.& Zucc.) Schott & Endl. | Malvaceae | 0.99989 | 0.01483 | 0.0304 | 0.823 |
| Erythroxylum deciduum A.St.-Hil | Erythroxylaceae | −0.22797 | −0.97367 | 0.0002 | 0.999 |
| Erythroxy suberosum A.St.-Hil | Erythroxylaceae | −0.86718 | 0.498 | 0.7576 | 0.001 *** |
| Erythroxylum tortuosum A.St.-Hil | Erythroxylaceae | 0.76754 | −0.641 | 0.2761 | 0.125 |
| Guapira noxia (Netto) Lundell | Nyctaginaceae | 0.77366 | −0.6336 | 0.4021 | 0.047 * |
| Handroanthus ochraceus (Chm.) Mattos | Bignoniaceae | 0.84702 | 0.53156 | 0.1538 | 0.370 |
| Hancornia speciosa Gomes | Apocynaceae | −0.22933 | −0.97335 | 0.1786 | 0.412 |
| Hymenaea stigonocarpa Mart. Ex Hayne | Fabaceae | 0.32592 | −0.9454 | 0.4944 | 0.026 * |
| Heteropterys byrsonimiifolia A.Juss | Malpighiaceae | 0.45763 | −0.88914 | 0.0534 | 0.768 |
| Kielmeyera coriacea Mart. & Zucc | Callophyllaceae | 0.05631 | 0.99841 | 0.1677 | 0.411 |
| Kielmeyera speciosa A.St.-Hil. | Callophyllaceae | 0.25950 | 0.96574 | 0.4671 | 0.022 * |
| Lafoensia pacari A.St.-Hil. | Loganiaeceae | 0.88586 | −0.46396 | 0.0725 | 0.861 |
| Leptolobium dasycarpum Vogel | Fabaceae | −0.49854 | −0.86686 | 0.1652 | 0.359 |
| Machaerium opacum Vogel | Fabaceae | 0.73150 | −0.68184 | 0.4537 | 0.013 * |
| Miconia albicans (Sw.) Triana | Melastomataceae | 0.60432 | 0.79674 | 0.2459 | 0.162 |
| Miconia leucocarpa DC | Melastomataceae | −0.91636 | −0.40035 | 0.6099 | 0.002 ** |
| Miconia burchelli Triana | Melastomataceae | 0.16307 | 0.98661 | 0.3891 | 0.046 * |
| Miconia speciosa (A.St.-Hil. & Naudin) Naudin | Melastomataceae | 0.13303 | 0.99111 | 0.1687 | 0.519 |
| Miconia ferruginata DC. | Melastomataceae | 0.66501 | −0.74684 | 0.6406 | 0.004 ** |
| Mimosa claussenii Benth | Fabaceae | 0.76611 | −0.64271 | 0.1956 | 0.265 |
| Myrsine guianensis (Aubl.) Kuntze | Primulaceae | −0.04177 | 0.99913 | 0.1592 | 0.425 |
| Neea theifera Oerst | Nyctaginaceae | −0.9057 | −0.42392 | 0.0731 | 0.640 |
| Ouratea hexasperma (A.St.-Hil.) Baill | Ochnaceae. | 0.10205 | 0.99478 | 0.0456 | 0.739 |
| Palicourea rigida Kunth | Rubiaceae | 0.21823 | 0.9759 | 0.3388 | 0.061 |
| Pouteria ramiflora (Mart.) Radlk | Sapotaceae | 0.60027 | −0.79979 | 0.4059 | 0.027 * |
| Piptocarpha rotundifolia (Less.) Baker | Asteraceae | −0.84519 | −0.53447 | 0.2268 | 0.209 |
| Psidium laruotteanum Cambess | Myrtaceae | 0.19464 | 0.98087 | 0.0218 | 0.913 |
| Plenckia populnea Reissek | Celastraceae | −0.71841 | −0.69562 | 0.2406 | 0.224 |
| Pterodon emarginatus Vogel | Fabaceae | 0.79357 | −0.60848 | 0.3462 | 0.072 |
| Qualea grandiflora Mart. | Vochysiaceae | 0.42728 | −0.90412 | 0.3066 | 0.129 |
| Qualea multiflora Mart. | Vochysiaceae | −0.03812 | 0.99927 | 0.1074 | 0.524 |
| Qualea parviflora Mart. | Vochysiaceae | 0.86802 | −0.49653 | 0.2768 | 0.123 |
| Roupala montana Aubl. | Proteaceae | 0.50624 | 0.86239 | 0.7116 | 0.001 *** |
| Rourea induta Planch. | Connaraceae | 0.98941 | 0.14514 | 0.0767 | 0.674 |
| Salacia crassifolia (Mart. ex Schult.) G.Don | Celastraceae | 0.63236 | −0.77468 | 0.2087 | 0.275 |
| Schefflera macrocarpa (Cham. & Schltdl.) Frodin | Araliaceae | 0.49271 | −0.87019 | 0.7362 | 0.001 *** |
| Symplocos rhamnifolia A.DC. | Symplocaceae. | 0.53729 | 0.8434 | 0.2863 | 0.147 |
| Strychnos pseudoquina A. St.-Hil. | Loganiaceae | 0.83442 | −0.55114 | 0.1168 | 0.541 |
| Syagrus comosa Mart. | Arecaceae | 0.05081 | 0.99871 | 0.0976 | 0.609 |
| Styrax ferrugineus Nees. & Mart | Styracaceae | 0.86754 | −0.49737 | 0.6936 | 0.001 *** |
| Stryphnodendron adstringens (Mart.) Coville | Fabaceae | 0.77755 | −0.62883 | 0.1788 | 0.291 |
| Solanum lycocarpum A.St.-Hill | Solanaceae | 0.00000 | 0 | 0.0000 | 1.000 |
| Tachigali subvelutina (Benth.) Oliveira-Filho | Fabaceae | −0.49209 | −0.87054 | 0.1239 | 0.422 |
| Vatairea macrocarpa (Benth.) Ducke | Leguminoseae | 0.35803 | −0.93371 | 0.0222 | 0.900 |
| Vochysia thyrsoidea Pohl | Vochysiaceae | 0.86539 | −0.50109 | 0.5867 | 0.002 ** |
| Vochysia elliptica Mart. | Vochysiaceae | 0.67429 | 0.73847 | 0.4986 | 0.012 * |
References
- Zimbres, B.; Shimbo, J.; Bustamante, M.; Levick, S.; Miranda, S.; Roitman, I.; Silvério, D.; Gomes, L.; Fagg, C.; Alencar, A. A estrutura da vegetação de savana no Cerrado brasileiro permite a estimativa precisa da biomassa acima do solo usando varredura a laser terrestre. For. Ecol. Manag. 2020, 458, 117798. [Google Scholar] [CrossRef]
- Ribeiro, J.F.; Walter, B.M.T. As principais fitofisionomias do bioma Cerrado. In Cerrado: Ecologia e Fóruns; Sano, S.M., Almeida, S.P., Ribeiro, J.F., Eds.; Embrapa Informação Tecnológica: Brasília, Brazil, 2008; pp. 151–212. Available online: https://www.infoteca.cnptia.embrapa.br/infoteca/handle/doc/570911 (accessed on 20 March 2023).
- Terra, M.d.C.N.S.; Prado-Júnior, J.A.D.; de Souza, C.R.; Pinto, L.O.R.; Silveira, E.M.d.O.; Cordeiro, N.G.; Cirne-Silva, T.M.; Mantovani, V.A.; Scolforo, J.R.S.; de Mello, J. MDominância de espécies arbóreas na biomassa e produtividade aérea da savana neotropical. For. Ecol. Manag. 2021, 496, 119430. [Google Scholar] [CrossRef]
- Sano, E.E.; Rodrigues, A.A.; Martins, E.S.; Bettiol, G.M.; Bustamante, M.M.; Bezerra, A.S.; Couto, A.F., Jr.; Vasconcelos, V.; Schüler, J.; Bolfe, E.L. Ecorregiões do Cerrado: Uma estrutura espacial para avaliar e priorizar a diversidade ambiental da savana brasileira para conservação. Rev. Gestão Ambient. 2019, 232, 818–828. Available online: https://pubmed.ncbi.nlm.nih.gov/30529869 (accessed on 15 February 2023).
- Rocha, G.F.; Ferreira, L.G.; Ferreira, N.C.; Ferreira, M.E. Detecção de desmatamento no bioma Cerrado entre 2002 e 2009: Padrões, Tendências e Impactos. Rev. Bras. Cartogr. 2011, 63, 341–349. [Google Scholar] [CrossRef]
- Swann, A.L.S. Plantas e seca num clima em mudança. Curr. Clim. Chang. Rep. 2018, 4, 192–201. [Google Scholar] [CrossRef]
- Strassburg, B.B.; Brooks, T.; Feltran-Barbieri, R.; Iribarrem, A.; Crouzeilles, R.; Loyola, R.; Latawiec, A.E.; Oliveira Filho, F.J.; Scaramuzza, C.A.; Scarano, F.R.; et al. Momento da Verdade Para o Hotspot do Cerrado. Ecol. Evol. Nat. 2017, 1, 99. Available online: https://www.nature.com/articles/s41559-017-0099 (accessed on 27 February 2023). [CrossRef] [PubMed]
- Brasil Ministério do Meio Ambiente. Cadastro Nacional de Unidades de Conservação; MMA: Brasília, Brazil, 2016. Available online: http://www.mma.gov.br/images/arquivo/80112/CNUC_Agosto%20-%20Biomas%201.pdf (accessed on 20 July 2021).
- Myers, N.; Mittermeler, R.A.; Mittermeler, C.G.; Da Fonseca, G.A.B.; Kent, J. Hotspots de biodiversidade para prioridades de conservação. Natureza 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Silveira, E.M.O.; Silva, S.H.G.; Acerbi-Júnior, F.W.; Carvalho, M.C.; Carvalho, L.M.T.; Scolforo, J.R.S.; Wulder, M.A. A modelagem florestal aleatória baseada em objetos de biomassa florestal acima do solo supera uma abordagem baseada em pixels em um ambiente tropical montanhoso heterogêneo. Int. J. Appl. Earth Obs. Geoinf. 2019, 78, 175–188. [Google Scholar] [CrossRef]
- Gomes, L.; Miranda, H.S.; Maria, M. Ecologia e Manejo Florestal Como podemos avançar no conhecimento sobre o comportamento e os efeitos do fogo no bioma Cerrado? Ecol. e Manejo Florest. 2018, 417, 281–290. Available online: https://www.sciencedirect.com/science/article/abs/pii/S0378112717321850 (accessed on 15 April 2023). [CrossRef]
- Strassburgo, B.B.N.; Iribarrem, A.; Beyer, H.L.; Cordeiro, C.L.; Crouzeilles, R.; Jakovac, C.C.; Braga Junqueira, A.; Lacerda, E.; Latawiec, A.E.; Balmford, A.; et al. Áreas prioritárias globais para a restauração de ecossistemas. Natureza 2020, 586, 724–729. [Google Scholar] [CrossRef]
- Coutinho, L.M. O fogo na ecologia do Cerrado brasileiro. In Fogo na Biota Tropical: Processos Ecossistêmicos e Desafios Globais; Goldammer, J.G., Ed.; Springer: Berlin, Germany, 1990; pp. 82–105. Available online: https://link.springer.com/chapter/10.1007/978-3-642-75395-4_6 (accessed on 11 March 2023).
- Mistry, J. Savanas Mundiais: Ecologia e Uso Humano; Pearson Education Limited, Prentice Hall: Grã-Bretanha, UK, 2000. [Google Scholar] [CrossRef]
- Pereira, K.M.G.; Cordeiro, N.G.; Terra, M.d.C.N.S.; Pyles, M.V.; Cabacinha, C.D.; de Mello, J.M.; Berg, E.v.D. Status de proteção como determinante dos drivers de estoque de carbono no Cerrado sensu stricto. J. Ecol. Veg. 2020, 13, 361–368. [Google Scholar] [CrossRef]
- Cianciaruso, M.V.; Silva, I.A.; Batalha, M.A. Biomassa aérea de grupos funcionais na camada subterrânea de savanas sob diferentes frequências de fogo. J. Aust. Bot. 2010, 58, 169–174. [Google Scholar] [CrossRef]
- Haridasan, M. Nutrição mineral de plantas nativas do cerrado. Rev. Bras. Fisiol. Veg. 2000, 12, 54–64. [Google Scholar]
- Eiten, G. Cerrado: Caracterização, Ocupação e Perspectivas; UnB/SEMATEC: Brasília, Brazil, 1994. [Google Scholar]
- Medeiros, M.B.; Miranda, H.S. Mortalidade pós-fogo em espécies lenhosas de campo sujas submetidas a três queimadas prescritas anualmente. Acta Bot. Bras. 2005, 19, 493–500. [Google Scholar] [CrossRef]
- Miranda, H.S. Efeitos do Regime de Fogo Sobre a Estrutura de Comunidades do Cerrado: Projeto Fogo; IBAMA: Brasília, Brazil, 2010; p. 144. [Google Scholar]
- Ferraz-Vicentini, K.R.C. História do Fogo no Cerrado. Ph.D. Thesis, Departamento de Ecologia, Universidade de Brasília, Brasília, Brazil, 1999. [Google Scholar]
- Solbrig, O.T.; Young, M.D. Forças motrizes económicas e ecológicas que afectam as savanas tropicais. In As Savanas do Mundo: Forças Motrizes Económicas, Restrições Ecológicas e Opções Políticas para o uso Sustentável da Terra; Young, M.D., Solbrig, O.T., Eds.; Série Homem e Biosfera; The Parthenon Publishing Group: Unesco, Paris, 1993; Volume 12, pp. 3–18. [Google Scholar]
- Pivello, V.R.; Coutinho, L.M. Firetool: Um sistema especialista para uso de fogos prescritos em áreas de conservação do cerrado. For. Ecol. Manag. 1996, 33, 348–356. [Google Scholar] [CrossRef]
- Eloy, L.; Schmidt, I.B.; Borges, S.L.; Ferreira, M.C.; Dos Santos, T.A. O manejo sazonal do fogo por pecuaristas tradicionais evita a propagação de incêndios florestais no Cerrado brasileiro. Ambio 2019, 48, 890–899. [Google Scholar] [CrossRef]
- Bilbau, B.; Mistério, J.; Millán, A.; Berardi, A. Compartilhando múltiplas perspectivas sobre queimadas: Rumo a uma política participativa e intercultural de manejo do fogo na Venezuela, Brasil e Guiana. Fogo 2019, 2, 39. [Google Scholar] [CrossRef]
- Hopkins, B. Processos ecológicos na fronteira floresta-savana. In Natureza e Dinâmica dos Limites Floresta-Savana; Furley, P.A., Proctor, J., Ratter, J.A., Eds.; Chapman & Hall: London, UK, 1992; pp. 21–33. [Google Scholar]
- Rezende, A.V.; Vale, A.T.; Sanquetta, C.R.; Figueiredo Filho, A.; Felfili, J.M. Comparação de modelos matemáticos para estimativa de volume, biomassa e estoque de carbono do tecido lenhoso de um cerrado sensu stricto em Brasília, DF. Sci. For. 2006, 71, 65–76. [Google Scholar]
- Sanquetta, C.R. Métodos de determinação de biomassa florestal. In As Florestas e o Carbono; Sanquetta, C.R., Ed.; Embrapa Florestas: Curitiba, Brazil, 2002; pp. 119–140. [Google Scholar]
- Felfili, J.M.; Carvalho, F.A.; Haidar, R.F. Manual Para o Monitoramento de Parcelas Permanentes nos Biomas Cerrado e Pantanal; Universidade de Brasília: Brasília, Brazil, 2005. [Google Scholar]
- Granado, L.M.A. Estimativa de Biomassa e Combustível em Diferentes Fitofisionomias do Cerrado. Master’s Thesis, Faculdade de Tecnologia, Universidade de Brasília, Brasília, Brazil, 2019; 71p. [Google Scholar]
- Koppen, W. Climatologia: Com um Estudo dos Climas da Terra; Fundo de Cultura Económica: Mexico City, Mexico, 1948; 479p. [Google Scholar]
- Nimer, E. Climatologia do Brasil; IBGE: Rio de Janeiro, Brazil, 1989; 422p. [Google Scholar]
- Haridasan, M. Adaptações nutricionais de plantas nativas do bioma cerrado em solos ácidos. Braz. J. Fisiol Vegetal. 2008, 20, 183–195. [Google Scholar] [CrossRef]
- Santos, G.L.; Pereira, M.G.; Delgado, R.C.; Torres, J.L.R. Regeneração Natural em Ambientes Antropogênicos Devido ao Uso Agrícola no Cerrado, Uberaba, Mg, Brasil. Rev. Biociênc. 2017, 1, 169–263. [Google Scholar] [CrossRef]
- Felfili, J.M.; Rezende, A.V.; Júnior, M.C.D.S.; Silva, M.A. Mudanças na composição florística do cerrado sensu stricto no Brasil ao longo de um período de nove anos. J. Trop. Ecol. 2000, 16, 579–590. [Google Scholar] [CrossRef]
- Azevedo, G.B. Amostragem e Modelagem da Biomassa de Raízes em um Cerrado Sentido Restrito no Distrito Federal. Master’s Thesis, Universidade de Brasília, Brasília, Brazil, 2014. [Google Scholar]
- Felfili, J.; Nogueira, P.E.; Silva Júnior, M.C.; Marimon, B.S.; Delitti, W.B.C. Composição florística e fitossociologia do cerrado sentido restrito no município de Água Boa-MT. Acta Bot. Bras. 2002, 16, 103–112. [Google Scholar] [CrossRef]
- Felfili, J.M.; Rezende, R.P. Conceitos e Métodos em Fitossociologia; Comunicações Técnicas Florestais; Universidade de Brasília, Departamento de Engenharia Florestal: Brasília, Brazil, 2003; Volume 5, pp. 1–68. [Google Scholar]
- Durigan, G. Estrutura e diversidade de florestas tropicais. In Ecologia de Florestas Tropicais do Brasil; Martins, S.V., Ed.; Editora UF: Viçosa, Brazil, 2009. [Google Scholar]
- Grupo de Filogenia de Angiospermas (APG IV). Uma atualização da classificação do Grupo de Filogenia de Angiospermas para as ordens e famílias de plantas com flores: APG IV. Rev. Bot. Soc. Linneana 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Forzza, R.C.; Baumgratz, J.F.A.; Bicudo, C.E.M.; Canhos, D.A.L.; Carvalho, A.A.; Coelho, M.A.N.; Costa, A.F.; Costa, D.P.; Hopkins, M.G.; Leitman, P.M.; et al. Nova lista florística brasileira destaca desafios de conservação. BioCiência 2012, 62, 39–45. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. A Teoria Matemática da Comunicação; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Pielou, C.E. Diversidade Ecológica; Wiley Interscience: New York, NY, USA, 1975. [Google Scholar]
- Lamprecht, H. Silvicultura nos Trópicos: Ecossistemas Florestais e Espécies Arbóreas Específicas—Possibilidades e Métodos de Aproveitamento Sustentado; República Federal da Alemanha: Rossdorf, Germany, 1990. [Google Scholar]
- Mueller-Dombois, D.; Ellenberg, H. Objetivos e Métodos de Ecologia do Vegetação; The Blackburn Press: Caldwell, NJ, USA, 2002; 547p. [Google Scholar]
- Kent, M.; Coker, P. Descrição da Vegetação: Uma Abordagem Prática; Imprensa de Belhaven: London, UK, 1992; 363p. [Google Scholar]
- IPCC. Painel Intergovernamental sobre Mudanças Climáticas (IPCC), Programa Nacional de Inventários de Gases de Efeito Estufa; Instituto de Estratégias Ambientais Globais: Hayama, Japan, 2006; Available online: http://www.ipcc-nggip.iges.or.jp/public/2006gl/index.html (accessed on 15 May 2023).
- Dixon, P. Vegan, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Legendre, P. Studying beta diversity: Ecological variation partitioning by multiple regression and canonical analysis. J. Plant Ecol. 2008, 1, 3–8. [Google Scholar] [CrossRef]
- Equipe Principal, R. R: Uma Linguagem e Ambiente Para Computação Estatística (Versão v3.6.2). Fundação R para Computação Estatística. 2019. Available online: https://www.R-project.org/ (accessed on 7 May 2023).
- Klink, C.A.; Machado, R.B. Conservação do Cerrado Brasileiro. Conservar. Biol. 2005, 19, 707–713. [Google Scholar] [CrossRef]
- Dantas, J.S. Relação Solo-Paisagem e Predição da Erodibilidade de Solos Coesos dos Tabuleiros Costeiros no Estado do Maranhão. Tese (Programa de Pós Graduação em Agronomia—Ciência do Solo)—Faculdade de Ciências Agrárias e Veterinárias; Universidade Estadual Paulista “Júlio de Mesquita Filho”: Jaboticabal, Brazil, 2013. [Google Scholar]
- Souchie, F.F.; Pinto, J.R.R.; Lenza, E.; Gomes, L.; Maracahipes-Santos, L.; Silvério, D.V. Estratégias de rebrota pós-fogo da vegetação lenhosa no cerrado brasileiro. Acta Bot. Bras. 2017, 31, 260–266. [Google Scholar] [CrossRef]
- Lopes, E.R.N.; Silva, A.P.P.S.; Peruchi, J.F. Zoneamento de Risco de Incêndio e Queimadas no Município de Sorocaba—São Paulo; Revista do Departamento de Geografia; Universidade de São Paulo: São Paulo, Brazil, 2018; Volume 36. [Google Scholar]
- Goldenberg, R.; Baumgratz, J.F.A.; Souza, M.L.D.E.R. Taxonomia de Melastomataceae no Brasil: Retrospectiva, perspectivas e chave de identificação para os gêneros. Rodriguésia 2012, 63, 145–161. [Google Scholar] [CrossRef]
- Medeiros, M.B. Efeitos do Fogo Nos Padrões de Rebrotamento em Plantas Lenhosas, em Campo sujo, após Queimadas Prescritas; Universidade de Brasília: Brasília, Brazil, 2002. [Google Scholar]
- Hoffmann, W.A.; Moreira, A. O papel do fogo na dinâmica populacional de plantas lenhosas. In Ecologia e História Natural de uma Savana Neotropical: Os cerrados do Brasil. A Imprensa da Universidade de Columbia; Oliveira, P.S., Marquês, R.S., Eds.; Columbia University Press: New York, NY, USA, 2002. [Google Scholar] [CrossRef]
- Felfili, J.M. Dinâmica da regeneração natural na mata de galeria do Gama no Brasil central. For. Ecol. Manag. 1997, 91, 235–245. [Google Scholar] [CrossRef]
- Oliveira, P.T.S.; Leite, M.B.; Mattos, T.; Aproximando-se, M.A.; Scott, R.L.; Oliveira Xavier, R.; Silva Matos, D.M.; Wendland, E. A recarga das águas subterrâneas diminui com o aumento da densidade da vegetação no cerrado brasileiro. Ecohidrologia 2017, 10, 1759. [Google Scholar] [CrossRef]
- Hoffmann, W.A. Incêndio e dinâmica populacional de plantas lenhosas em uma savana neotropical: Projeções de modelos matriciais. Ecologia 1999, 80, 1354–1369. [Google Scholar] [CrossRef]
- do Vale, A.T.; Elias, P.S. Nível de proteção térmica da cascata de quatro espécies lenhosas e a relação da arquitetura da cascata com a transferência de calor. Ciênc. Florest. 2014, 24, 979–987. [Google Scholar] [CrossRef]
- Salgado-Labouriau, M.L.; Ferraz-Vicentini, K.R. Incêndio no Cerrado há 32.000 anos. Pesqui. Atual No Pleistoceno 1994, 11, 85–87. [Google Scholar]
- Lima, J.M.; Castro, A.B.; Lima, A.P.; Magnusson, W.E.; Landeiro, V.L.; Fadini, R.F. Influência do regime de queimadas sobre a riqueza e composição florística de uma savana isolada na Amazônia—PELD Oeste do Pará. Oecologia Aust. 2020, 24, 301–316. [Google Scholar] [CrossRef]
- Miranda, H.S.; Sato, M.; Andrade, S.M.; Haridasan, M.; Moraes, H.C. Queimadas de Cerrado: Caracterização e impactos. In Cerrado: Ecologia e Caracterização; Aguiar, L.M.S., Camargo, A.J.A., Eds.; Embrapa Cerrados: Brasília, Brazil, 2004; pp. 69–123. [Google Scholar]
- Pivello, V.R. Manejo do fogo para conservação biológica no cerrado brasileiro. In Savanas e Florestas Secas; Mistry, J., Berardi, A., Eds.; Routledge: Abingdon-on-Thames, UK, 2017; pp. 141–166. [Google Scholar] [CrossRef]
- Pivello, V.R. O Uso do Fogo no Cerrado e nas Florestas Amazônicas do Brasil: Passado e Presente. Ecol. Do Fogo 2011, 7, 24–39. [Google Scholar] [CrossRef]
- Bowman, D.M.; Murphy, B.P.; Boer, M.M.; A Bradstock, R.; Cary, G.J.; A Cochrane, M.; Fensham, R.J.; A Krawchuk, M.; Price, O.F.; Williams, R.J. Gestão de incêndios florestais, mudanças climáticas e o risco de perdas catastróficas de carbono. Front. Ecol. Environ. 2013, 11, 66–68. [Google Scholar] [CrossRef]
- Bond, W.J.; Archibald, S. Enfrentando a complexidade: Escolhas de políticas de incêndio em parques de savana sul-africanos. Int. J. Wildland Fire 2003, 12, 381–389. [Google Scholar] [CrossRef]
- Batista, E.K.L.; Russell-smith, J.; França, H.; Figueira, J.E.C. Uma avaliação dos regimes contemporâneos de incêndios na savana no Parque Nacional da Canastra, Brasil: Resultados das políticas de supressão de incêndios. J. Meio Ambiente Gerenciar. 2018, 205, 40–49. [Google Scholar]
- Fidelis, A.; Alvarado, S.T.; Barradas, A.C.S.; Pivello, V.R. O ano de 2017: Megaincêndios e gestão no Cerrado. Fire 2018, 1, 49. [Google Scholar] [CrossRef]
- Ramos, P.C.M. Sistema Nacional de Prevenção e Combate aos Incêndios Florestais; IPEF: Piracicaba, Brazil, 1995; pp. 29–58. [Google Scholar]
- Durigan, G. Fogo zero: Não é possível nem desejável no Cerrado do Brasil. Flora 2020, 268, 15161. [Google Scholar] [CrossRef]
- Abreu, R.C.R.; Hoffmann, W.A.; Vasconcelos, H.L.; Pilon, N.A.; Rossatto, D.R.; Durigan, G. O custo da biodiversidade do sequestro de carbono na savana tropical. Sci. Adv. 2017, 3, 1701284. [Google Scholar] [CrossRef] [PubMed]
- Durigan, G.; Leitão Filho, H.F.; Rodrigues, R.R. Fitossociologia e estenose de uma vegetação de cerrado frequentemente queimada no Sudeste do Brasil. Flora 1994, 189, 153–160. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Cummings, D.L.; Ward, D.E. Relações entre fogo, biomassa e dinâmica de nutrientes ao longo de um gradiente de vegetação no Cerrado brasileiro. J. Ecol. 1994, 82, 519–531. [Google Scholar] [CrossRef]
- Koch, A.; Brierley, C.; Maslin, M.M.; Lewis, S.L. Impactos no sistema terrestre de chegada europeia e da Grande Morte nas Américas após 1492. Quat. Ciênc. 2019, 207, 13–36. [Google Scholar]
- Almeida, M.A. Modelagem da Propagação do Fogo Como Ferramenta de Auxílio à Tomada de Decisão no Combate e Prevenção de Incêndios no Parque Nacional das Emas, GO; Instituto Nacional de Pesquisas Espaciais-INPE: São José dos Campos, Brazil, 2012. [Google Scholar]
- Lopes, J.F.; de Andrade, E.M.; de Oliveira Lobato, F.A.; de Queiroz Palácio, H.A.; Arraes, F.D. Deposição e acomodação de serapilheira em área da Caatinga. Rev. Agro@ Mbiente-Line 2009, 2, 72–79. [Google Scholar]
- Moreira, A.G. Efeitos da proteção contra fogo na estrutura das savanas do Brasil Central. Rev. Biogeogr. 2000, 27, 1021–1029. [Google Scholar] [CrossRef]
- Fiedler, N.C.; Azevedo, I.N.; Rezende, A.V.; Medeiros, M.B.; Venturoli, F. Efeito de incêndios florestais na estrutura e composição florística de uma área de cerrado sentido estrito na Fazenda Água Limpa-DF. Rev. Árvore 2004, 28, 129–138. [Google Scholar] [CrossRef]
- Líbano, A.M.; Felfili, M.J. Mudanças temporais na composição florística e na diversidade de um cerrado sensu stricto do Brasil Central em um período de 18 anos. Acta Bot. Bras. 2006, 20, 927–936. [Google Scholar] [CrossRef]
- Roitman, I.; Bustamante, M.M.C.; Haidar, R.F.; Shimbo, J.Z.; Abdala, G.C.; Eiten, G.; Fagg, C.W.; Felfili, M.C.; Felfili, J.M.; Jacobson, T.K.B.; et al. Otimizando estimativas de biomassa de florestas de savana em diferentes escalas espaciais no Cerrado brasileiro: Reavaliando equações alométricas e influências ambientais. PLoS ONE 2018, 13, e0196742. [Google Scholar] [CrossRef]
- Ribeiro, M.C.; Metzger, J.P.; Martensen, A.C.; Ponzoni, F.J.; Hirota, M.M. A Mata Atlântica Brasileira: Quanto resta e como está distribuída a floresta remanescente? Implicações para a conservação. Conserv. Biol. 2009, 142, 1141–1153. [Google Scholar] [CrossRef]
- Bernhardt-Römermann, M.; Baeten, L.; Craven, D.; De Frenne, P.; Hédl, R.; Lenoir, J.; Bert, D.; Brunet, J.; Chudomelová, M.; Decocq, G.; et al. Os impulsionadores das mudanças temporais na diversidade de plantas das florestas temperadas variação entre as escalas espaciais. Glob. Chang. Biol. 2015, 21, 3726–3737. [Google Scholar] [CrossRef]
- Altomare, M.; Vasconcelos, H.L.; Raymundo, D.; Lopes, S.; Vale, V.; Prado-Junior, J. Avaliação da resiliência ao fogo do componente arbóreo da savana através de uma abordagem funcional. Acta Oecol. 2021, 111, 103728. [Google Scholar] [CrossRef]
- Sato, N.M. Mortalidade de Plantas do Cerrado Submetidas a Diferentes Regimes de Queima. Ph.D. Thesis, Universidade de Brasília, Brasília, Brazil, 1996. [Google Scholar]
- Ramos-Neto, M.B.; Pivello, V.R. Incêndios de raios em um Parque Nacional do Cerrado Brasileiro: Repensando estratégias de manejo. Gestão Ambient. 2000, 26, 675–684. [Google Scholar] [CrossRef]
- Bond, W.J.; Keeley, J.E. O fogo como um ‘herbívoro’ global: A ecologia e a evolução dos ecossistemas inflamáveis. Tendências Em Ecol. E Evolução 2005, 20, 387–394. [Google Scholar] [CrossRef]
- Pausas, J.G.; Poorter, L. Espessura da casca e regime de fogo. Funct. Ecol. 2015, 29, 315–327. [Google Scholar] [CrossRef]
- Araújo, F.D.; Tng, D.Y.; Apgaua, D.M.; Coelho, P.A.; Pereira, D.G.; Santos, R.M. Regeneração vegetal pós-incêndio em transição florestal-savana fechada. For. Ecol. Manag. 2017, 400, 77–84. [Google Scholar] [CrossRef]
- Souza, C.R.; Coelho De Souza, F.; Maia, V.A.; Aguiar-Campos, N.; Coelho, P.A.; Farrapo, C.L.; Santos, A.B.M.; Araújo, F.C.; Gianasi, F.M.; Paula, G.G.P.; et al. Estrutura e diversidade das florestas tropicais: Uma comparação de escolhas metodológicas. Methods Ecol. Evol. 2021, 12, 2017–2027. [Google Scholar] [CrossRef]
- Lucas, R.; MacArthur, A. Incêndio florestal na Austrália; Serviço de Publicação do Governo Australiano: Canberra, Australia, 1978. [Google Scholar]
- Maracahipes, L.; Marimon, B.S.; Lenza, E.; Marimon-Júnior, B.H.; De Oliveira, E.A.; Mews, H.A.; Gomes, L.; Feldpausch, T.R. Dinâmica pós-fogo da vegetação lenhosa em florestas sazonalmente inundadas (impucas) na zona de transição Cerrado-Floresta Amazônica. Flora 2014, 209, 260–270. [Google Scholar] [CrossRef]
- Silveira, F.A.O.; Ordóñez-Parra, C.A.; Moura, L.C.; Schmidt, I.B.; Andersen, A.N.; Bond, W.; Buisson, E.; Durigan, G.; Fidelis, A.; Oliveira, R.S.; et al. A disparidade de conscientização do bioma é RUIM para a conservação e restauração de ecossistemas tropicais. J. Appl. Ecol. 2021, 59, 1967–1975. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J. Incêndios florestais e mudanças globais. Front. Em Ecol. E Meio Ambiente 2021, 19, 387–395. [Google Scholar] [CrossRef]
- Hagmann, R.; Hessburg, P.; Salter, R.; Merschel, A.; Reilly, M. Os incêndios florestais contemporâneos degradam ainda mais a resistência e a resiliência das florestas excluídas do fogo Ecologia e Gestão Florestal. For. Ecol. Manag. 2022, 506, 119975. [Google Scholar] [CrossRef]
- Hessburgo, P.F.; Miller, C.L.; Povak, N.A.; Taylor, A.H.; Higuera, P.E.; Prichard, S.J.; Norte, D.; Collins, B.M.; Hurteau, M.D.; Larson, A.J.; et al. O clima, o meio ambiente e a história de perturbações governam a resiliência das florestas do oeste da América do Norte. Front. Ecol. Evol. 2019, 7, 239. [Google Scholar] [CrossRef]
- Hessburgo, P.F.; Espiões, T.A.; Perry, D.A.; Skinner, C.N.; Taylor, A.H.; Marrom, P.M.; Stephens, S.L.; Larson, A.J.; Churchill, D.J.; Povak, N.A.; et al. Tamm Review: Gestão de florestas em regime de incêndio de gravidade mista em Oregon, Washington e norte da Califórnia. For. Ecol. Manag. 2016, 366, 221–250. [Google Scholar] [CrossRef]
- Flores, C.; Limites, D.L.; Rubi, D.E. O fogo prescrito beneficia a vegetação das zonas húmidas? Zonas Úmidas 2011, 31, 35–44. [Google Scholar] [CrossRef]
- Elogne, A.G.; Piponiot, C.; Zo-Bi, I.C.; Amani, B.H.; Van der Meersch, V.; Hérault, B. Life after fire—Long-term responses of 20 timber species in semi-deciduous forests of West Africa. For. Ecol. Manag. 2023, 538, 120977. [Google Scholar] [CrossRef]
- Lal, R. Sequestro de carbono. Transações Filosóficas Da R. Soc. B 2008, 363, 815–830. [Google Scholar] [CrossRef]
- Grace, J.; Jose, J.S.; Meir, P.; Miranda, H.S.; Montes, R.A. Produtividade e fluxos de carbono de savanas tropicais. Rev. Biogeogr. 2006, 33, 387–400. [Google Scholar] [CrossRef]
- Bustamante, M.M.C.; Oliveira, E.L. Impacto das atividades agrícolas, florestais e pecuárias nos recursos naturais. In Savanas: Desafios e Estratégias Para o Equilíbrio Entre Sociedade, Agronegócio e Recursos Naturais; Faleiro, F.G., Farias Neto, A.L., Eds.; Embrapa Cerrados: Planaltina, Brazil, 2008. [Google Scholar]
- Houghton, R.A.; Hall, F.; Goetz, S.J. Importância da biomassa no ciclo global do carbono. Biogeosciences 2009, 114, 935. [Google Scholar] [CrossRef]
- Paiva, A.O.; Rezende, A.V.; Pereira, R.S. Estoque de carbono no cerrado sensu stricto no Distrito Federal. Rev. Árvore 2011, 3, 527–538. [Google Scholar] [CrossRef]
- Bastin, J.-F.; Berrahmouni, N.; Grainger, A.; Maniatis, D.; Mollicone, D.; Moura, R.; Patriarca, C.; Picard, N.; Pardal, B.; Abraão, E.M.; et al. A extensão da floresta em biomas de sequeiro. Ciência 2017, 356, 635–638. [Google Scholar]
- Zhou, Y.; Singh, J.; Butnor, J.R.; Coetsee, C.; Boucher, P.B.; Caso, M.F.; Hockridge, E.G.; Davies, A.B.; Staver, A.C. Aumentos limitados nos estoques de carbono da savana ao longo de décadas de supressão de incêndios. Natureza 2022, 603, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Poorter, L.; Van Der Sande, M.T.; Thompson, J.; Arets, E.J.M.M.; Alarcón, A.; Álvarez-Sánchez, J.; Ascarrunz, N.; Balvanera, P.; Barajas-guzmán, G.; Boit, A.; et al. Diversity enhances carbon storage in tropical forests. Glob. Ecol. Biogeogr. 2015, 24, 1314–1328. [Google Scholar] [CrossRef]
- Piponiot, C.; Derroire, L.G.; Descroix, L.; Mazzei, E.; Rutishauser, P.; Sist, B. Hérault. Avaliando a recuperação do volume de madeira após perturbação em florestas tropicais—Uma nova estrutura de modelagem. Ecol. Model. 2018, 384, 353–369. [Google Scholar] [CrossRef]
- Chaves, J.; Andalo, C.; Marrom, S.; Cairns, M.A.; Câmaras, J.Q.; Éamus, D.; Folster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Alometria de árvores e estimativa aprimorada de estoques e equilíbrio de carbono em florestas tropicais. Ecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Nogueira, E.M.; Nelson, B.W.; Fearnside, P.M.; França, M.B.; Oliveira, A.C.A. Altura das árvores no “arco do desmatamento” do Brasil: árvores mais baixas no sul e sudoeste da Amazônia implicam menor biomassa. For. Ecol. Manag. 2008, 255, 2963–2972. [Google Scholar] [CrossRef]
- Alves, R.J.V.; Kolbek, J. A vegetação do campo rupestre pode ser delimitada floristicamente com base em gêneros de plantas vasculares? Planta Ecol. 2010, 207, 67–79. [Google Scholar] [CrossRef]
- Morandi, P.S.; Marimon, B.S.; Marimon-Junior, B.H.; Ratter, J.A.; Feldpausch, T.R.; Colli, G.R.; Munhoz, C.B.R.; Júnior, M.C.d.S.; Lima, E.d.S.; Haidar, R.F.; et al. Diversidade de árvores e biomassa acima do solo no bioma Cerrado da América do Sul e suas implicações para a conservação. Biodivers. Conserv. 2018, 29, 1519–1536. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neri, S.C.M.; Bomfim, B.; Pereira, R.S.; dos Santos, P.V.; Tetto, A.F. Decadal Fire Effects on the Structure, Composition, Diversity, and Aboveground Carbon Stocks of a Neotropical Savanna. Forests 2023, 14, 2294. https://doi.org/10.3390/f14122294
Neri SCM, Bomfim B, Pereira RS, dos Santos PV, Tetto AF. Decadal Fire Effects on the Structure, Composition, Diversity, and Aboveground Carbon Stocks of a Neotropical Savanna. Forests. 2023; 14(12):2294. https://doi.org/10.3390/f14122294
Chicago/Turabian StyleNeri, Sarah Cristine Martins, Barbara Bomfim, Reginaldo Sérgio Pereira, Pâmela Virgilio dos Santos, and Alexandre França Tetto. 2023. "Decadal Fire Effects on the Structure, Composition, Diversity, and Aboveground Carbon Stocks of a Neotropical Savanna" Forests 14, no. 12: 2294. https://doi.org/10.3390/f14122294
APA StyleNeri, S. C. M., Bomfim, B., Pereira, R. S., dos Santos, P. V., & Tetto, A. F. (2023). Decadal Fire Effects on the Structure, Composition, Diversity, and Aboveground Carbon Stocks of a Neotropical Savanna. Forests, 14(12), 2294. https://doi.org/10.3390/f14122294