Genetic Parameters and Selection Responses for Important Breeding Traits in Liquidambar formosana Based on a Provenance–Family Trial

Abstract
:1. Introduction
2. Materials and Methods
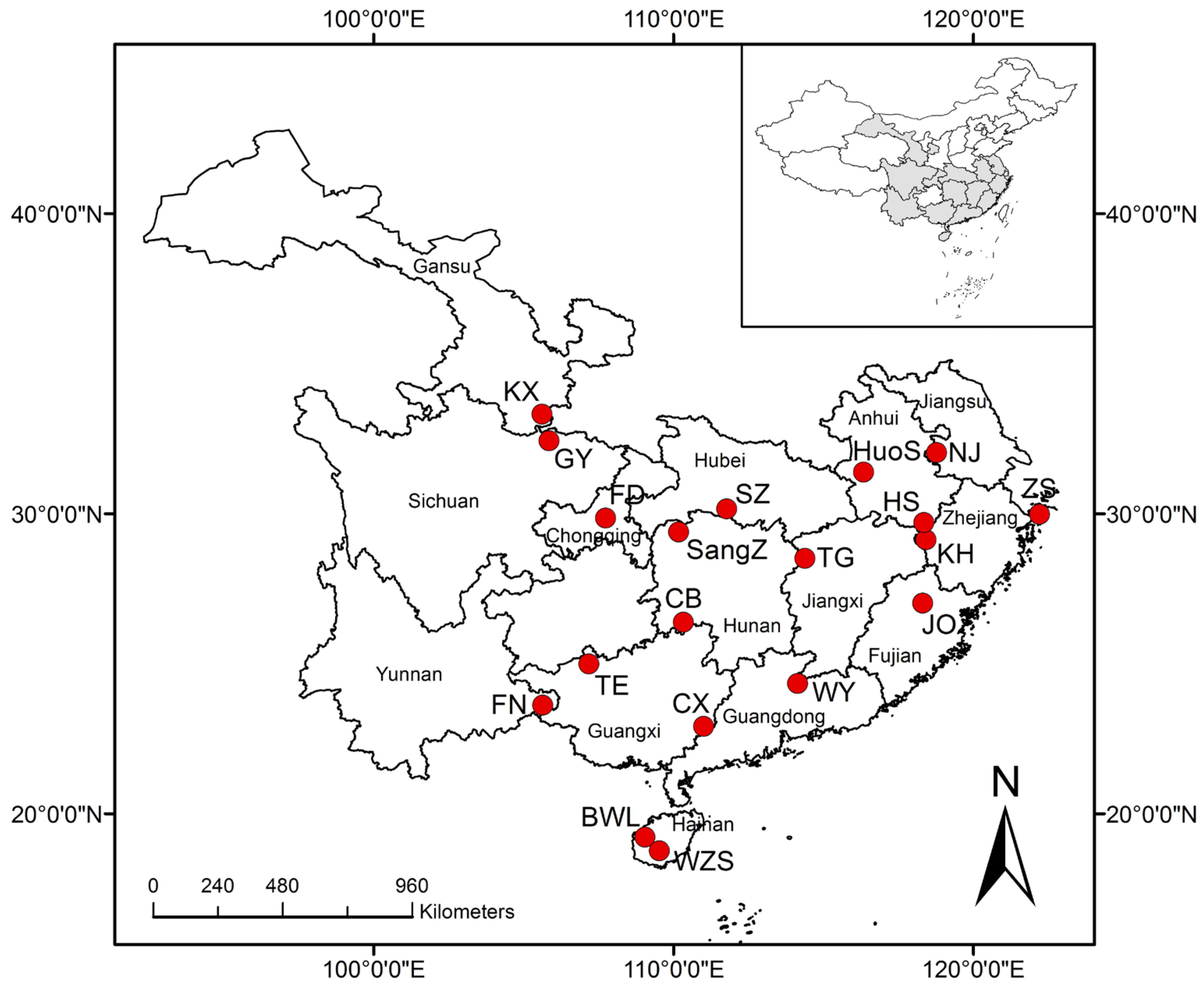
2.1. Genetic Material, Field Trial and Experimental Design
2.2. Shikimic Acid Extraction and Determination
2.3. Other Breeding Trait Measurements
2.4. Statistical Analysis
3. Results
3.1. Trait Variation, Variance Components and Heritability
3.2. Genetic and Phenotypic Correlations between Traits
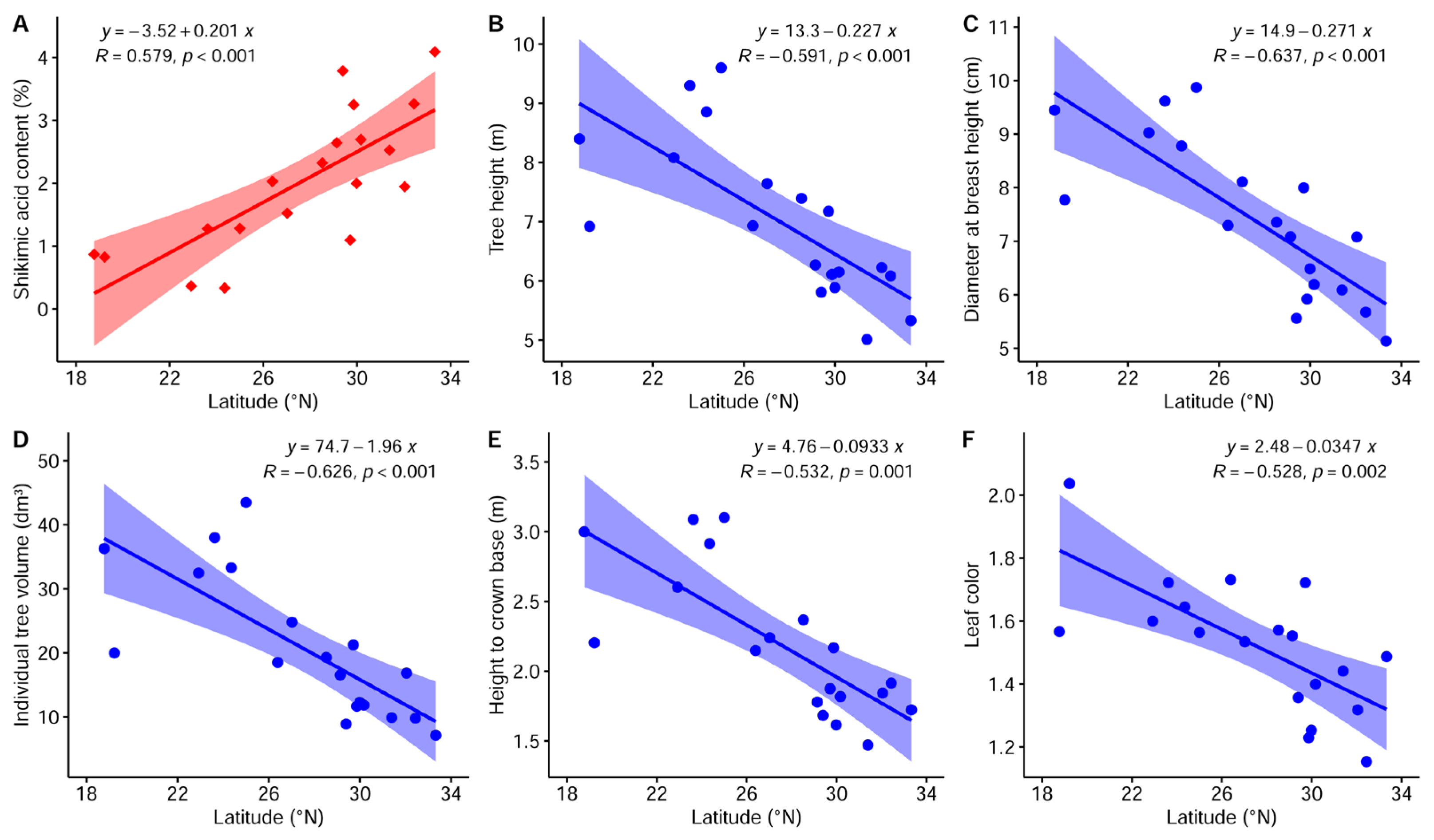
3.3. Geographical Structure of Breeding Traits
3.4. Genetic Gains under Different Selection Scenarios
4. Discussion
4.1. Mean, Variation and Heritability
4.2. Genetic and Phenotypic Correlations between Traits
4.3. Geographical Variation Patterns of Breeding Traits
4.4. Response for Different Selection Scenarios
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sun, R.; Lin, F.; Huang, P.; Zheng, Y. Moderate genetic diversity and genetic differentiation in the relict tree Liquidambar formosana Hance revealed by genic simple sequence repeat markers. Front. Plant Sci. 2016, 7, 1411. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qi, S.; Chen, S.; Li, H.; Zhang, T.; Bao, F.; Zhan, D.; Pang, Z.; Zhang, J.; Zhao, J. Genome-wide identification and expression analysis of late embryogenesis abundant (LEA) genes reveal their potential roles in somatic embryogenesis in hybrid sweetgum (Liquidambar styraciflua × Liquidambar formosana). For. Res. 2023, 3, 12. [Google Scholar] [CrossRef]
- Li, Y.; Gao, Y.; Deng, L.; Lian, H.; Guo, W.; Wu, W.; Xue, B.; Li, B.; Su, Y.; Zhang, H. Volatile profiling and transcriptome sequencing provide insights into the biosynthesis of α-pinene and β-pinene in Liquidambar formosana Hance leaves. Genes 2023, 14, 163. [Google Scholar] [CrossRef] [PubMed]
- Lingbeck, J.M.; O’Bryan, C.A.; Martin, E.M.; Adams, J.P.; Crandall, P.G. Sweetgum: An ancient source of beneficial compounds with modern benefits. Pharmacogn. Rev. 2015, 9, 1–11. [Google Scholar]
- Zhu, Y.; Guan, Y.J.; Chen, Q.Z.; Yuan, L.H.; Xu, Q.Q.; Zhou, M.L.; Liu, H.; Lin, W.; Zhang, Z.D.; Zhou, Z.L.; et al. Pentacyclic triterpenes from the resin of Liquidambar formosana have anti-angiogenic properties. Phytochemistry 2021, 184, 112676. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Chen, G.; Chen, K.; Chen, X.; Hong, Q.; Kan, J. Assessment of fresh star anise (Illicium verum Hook. f.) drying methods for influencing drying characteristics, color, flavor, volatile oil and shikimic acid. Food Chem. 2021, 342, 128359. [Google Scholar] [CrossRef]
- Cai, M.; Luo, Y.; Chen, J.; Liang, H.; Sun, P. Optimization and comparison of ultrasound-assisted extraction and microwave-assisted extraction of shikimic acid from Chinese star anise. Sep. Purif. Technol. 2014, 133, 375–379. [Google Scholar] [CrossRef]
- Candeias, N.R.; Assoah, B.; Simeonov, S.P. Production and synthetic modifications of shikimic acid. Chem. Rev. 2018, 118, 10458–10550. [Google Scholar] [CrossRef]
- Ghosh, S.; Chisti, Y.; Banerjee, U.C. Production of shikimic acid. Biotechnol. Adv. 2012, 30, 1425–1431. [Google Scholar] [CrossRef]
- Lai, J.L.; Chen, X.L.; Feng, J.H.; Huang, C.N.; Bei, Y.J. First report of Alternaria tenuissima causing leaf spot on star anise (Illicium verum) in China. Plant Dis. 2020, 105, 229. [Google Scholar] [CrossRef]
- Enrich, L.B.; Scheuermann, M.L.; Mohadjer, A.; Matthias, K.R.; Eller, C.F.; Newman, M.S.; Fujinaka, M.; Poon, T. Liquidambar styraciflua: A renewable source of shikimic acid. Tetrahedron Lett. 2008, 49, 2503–2505. [Google Scholar] [CrossRef]
- Martin, E.; Duke, J.; Pelkki, M.; Clausen, E.C.; Carrier, D.J. Sweetgum (Liquidambar styraciflua L.): Extraction of shikimic acid coupled to dilute acid pretreatment. Appl. Biochem. Biotechnol. 2010, 162, 1660–1668. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z. The Research on New Resources of Shikimic Acid and Its Isolation and Purification. Master’s Thesis, Zhejiang University, Hangzhou, China, 2010. [Google Scholar]
- Hu, W.; Pang, H.; Hu, X.; Wang, X.; Lin, F. Variation and selection of Liquidambar formosana based on a nine-year-old provenance test. J. Cent. South Univ. For. Technol. 2019, 39, 40–46. [Google Scholar]
- Ismael, A.; Klápštĕ, J.; Stovold, G.T.; Fleet, K.; Dungey, H. Genetic variation for economically important traits in Cupressus lusitanica in New Zealand. Front. Plant Sci. 2021, 12, 651729. [Google Scholar] [CrossRef]
- Hu, W.; Pang, H.; Hu, X.; Wang, X.; Zheng, Y. Genetic variation, excellent family and individual selection of 9-year-old Liquidambar formosana. J. Trop. Subtrop. Bot. 2018, 26, 506–514. [Google Scholar]
- Fang, L.; Shi, J.; Li, L.; Wu, X.; Shi, T. Analysis of genetic variation of progeny traits in Liquidambar Formosana. Sci. Silvae Sin. 2003, 39, 148–152. [Google Scholar]
- Zhou, W.; Xia, W.; Chen, Z.; Zhou, J.; Ni, W.; Ren, J. A new Liquidambar formosana cultivar ‘Fulu Zifeng’. Acta Hortic. Sin. 2019, 46, 2921–2922. [Google Scholar]
- Chen, X. Genetic variation and selection of 14-year-old Liquidambar formosana progeny. For. Res. 2015, 28, 183–187. [Google Scholar]
- DuVal, A.; Gezan, S.A.; Mustiga, G.; Stack, C.; Marelli, J.P.; Chaparro, J.; Livingstone, D.; Royaert, S.; Motamayor, J.C. Genetic parameters and the impact of off-types for Theobroma cacao L. in a breeding program in Brazil. Front. Plant Sci. 2017, 8, 2059. [Google Scholar] [CrossRef]
- Bonilla, J.L.S.; Lopes, U.V.; Colmenero, A.Z.; Valencia, B.B.M.; Arrazate, C.H.A.; Gramacho, K.P. Estimation of genetic parameters associated with frosty pod rot (Moniliophthora roreri) and cacao production in Mexico. Tree Genet. Genomes 2021, 17, 24. [Google Scholar] [CrossRef]
- Mustiga, G.M.; Gezan, S.A.; Phillips-Mora, W.; Arciniegas-Leal, A.; Mata-Quirós, A.; Motamayor, J.C. Phenotypic description of Theobroma cacao L. for yield and vigor traits from 34 hybrid families in Costa Rica based on the genetic basis of the parental population. Front. Plant Sci. 2018, 9, 808. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Wang, X.; Hu, X.; Lin, F.; Pang, H. Early selection of fine families and individuals of Liquidambar formosana. J. Northeast For. Univ. 2017, 45, 5–11. [Google Scholar]
- Robinson, A.R.; Ukrainetz, N.K.; Kang, K.Y.; Mansfield, S.D. Metabolite profiling of Douglas-fir (Pseudotsuga menziesii) field trials reveals strong environmental and weak genetic variation. New Phytol. 2007, 174, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Nantongo, J.S.; Potts, B.M.; Davies, N.W.; Fitzgerald, H.; Rodemann, T.; O’Reilly-Wapstra, J.M. Additive genetic variation in Pinus radiata bark chemistry and the chemical traits associated with variation in mammalian bark stripping. Heredity 2021, 127, 498–509. [Google Scholar] [CrossRef]
- Külheim, C.; Yeoh, S.H.; Wallis, I.R.; Laffan, S.; Moran, G.F.; Foley, W.J. The molecular basis of quantitative variation in foliar secondary metabolites in Eucalyptus globulus. New Phytol. 2011, 191, 1041–1053. [Google Scholar] [CrossRef]
- Gosney, B.J.; Potts, B.M.; O’Reilly-Wapstra, J.M.; Vaillancourt, R.E.; Fitzgerald, H.; Davies, N.W.; Freeman, J.S. Genetic control of cuticular wax compounds in Eucalyptus globulus. New Phytol. 2016, 209, 202–215. [Google Scholar] [CrossRef]
- Ferrão, L.F.V.; Johnson, T.S.; Benevenuto, J.; Edger, P.P.; Colquhoun, T.A.; Munoz, P.R. Genome-wide association of volatiles reveals candidate loci for blueberry flavor. New Phytol. 2020, 226, 1725–1737. [Google Scholar] [CrossRef]
- Ye, D. Open pollination progeny test and excellent family selection of Liquidambar formosana. J. Cent. South Univ. For. Technol. 2011, 31, 79–82. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kolde, R. Pretty Heatmaps. R Package Version 1.0.12. 2019. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 1 January 2022).
- Butler, D.; Cullis, B.; Gilmour, A.; Gogel, B.; Thompson, R. ASReml-R Reference Manual Version 4; VSN International Ltd.: Hemel Hempstead, UK, 2017. [Google Scholar]
- Sui, R. Separation of shikimic acid from pine needles. Chem. Eng. Technol. 2008, 31, 469–473. [Google Scholar] [CrossRef]
- Marchiosi, R.; Ferro, A.P.; Ramos, A.V.G.; Baldoqui, D.C.; Constantin, R.P.; Constantin, R.P.; dos Santos, W.D.; Ferrarese-Filho, O. Calophyllum brasiliense Cambess: An alternative and promising source of shikimic acid. Sustain. Chem. Pharm. 2019, 14, 100188. [Google Scholar] [CrossRef]
- Avula, B.; Wang, Y.H.; Smillie, T.J.; Khan, I.A. Determination of Shikimic acid in fruits of illicium species and various other plant samples by LC-UV and LC-ESI-MS. Chromatographia 2009, 69, 307–314. [Google Scholar] [CrossRef]
- Dong, J.; Liang, Z. Analysis on the factor influencing secondary metabolite accumulation in plants. Acta Bot. Boreal-Occident Sin. 2004, 24, 1979–1983. [Google Scholar]
- Bochkov, D.V.; Sysolyatin, S.V.; Kalashnikov, A.I.; Surmacheva, I.A. Shikimic acid: Review of its analytical, isolation, and purification techniques from plant and microbial sources. J. Chem. Biol. 2012, 5, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Hai, H.N.T.; Rimbawanto, A.; Prastyono; Kartikawati, N.K.; Wu, H. Genetic improvement for essential oil yield and quality in Melaleuca cajuputi. Ind. Crops Prod. 2019, 137, 681–686. [Google Scholar]
- Ofori, A.; Padi, F.K.; Ansah, F.O.; Akpetey, A.; Anin-Kwapong, G. Genetic variation for vigour and yield of cocoa (Theobroma cacao L.) clones in Ghana. Sci. Hortic. 2016, 213, 287–293. [Google Scholar] [CrossRef]
- Ghildiyal, V.; Iyiola, E.; Sharma, M.; Apiolaza, L.A.; Altaner, C. Genetic variation in drying collapse and heartwood properties at mid-rotation age of Eucalyptus globoidea. Ind. Crops Prod. 2023, 201, 116891. [Google Scholar] [CrossRef]
- Míguez-Soto, B.; Fernández-López, J. Genetic parameters and predicted selection responses for timber production traits in a Castanea sativa progeny trial: Developing a breeding program. Tree Genet. Genomes 2012, 8, 409–423. [Google Scholar] [CrossRef]
- Zhao, M.; Yan, C.; Wang, W.; Ye, J.; Zhong, Y.; Ke, Z.; Hao, X.; Ke, X.; Ye, L.; Huang, L. Relationship between selection of Pinus massoniana families and Folium Pini. China J. Chin. Mater. Med. 2015, 40, 1699–1704. [Google Scholar]
- Li, C.; Weng, Q.; Chen, J.B.; Li, M.; Zhou, C.; Chen, S.; Zhou, W.; Guo, D.; Lu, C.; Chen, J.C.; et al. Genetic parameters for growth and wood mechanical properties in Eucalyptus cloeziana F. Muell. New For. 2017, 48, 33–49. [Google Scholar] [CrossRef]
- Belaber, E.C.; Gauchat, M.E.; Rodríguez, G.H.; Borralho, N.M.; Cappa, E.P. Estimation of genetic parameters using spatial analysis of Pinus elliottii Engelm. var. elliottii second-generation progeny trials in Argentina. New For. 2019, 50, 605–627. [Google Scholar]
- Dong, M.; Fan, Y.; Wu, Z.; Lv, F.; Zhang, J. Age–age correlations and early selection for growth traits in 40 half-sib families of Larix principis-rupprechtii. J. For. Res. 2019, 30, 2111–2117. [Google Scholar] [CrossRef]
- Ding, C.; Hamann, A.; Yang, R.C.; Brouard, J.S. Genetic parameters of growth and adaptive traits in aspen (Populus tremuloides): Implications for tree breeding in a warming world. PLoS ONE 2020, 15, e0229225. [Google Scholar] [CrossRef] [PubMed]
- Díaz, R.; Zas, R.; Fernández-López, J. Genetic variation of Prunus avium in susceptibility to cherry leaf spot (Blumeriella jaapii) in spatially heterogeneous infected seed orchards. Ann. For. Sci. 2007, 64, 21–30. [Google Scholar] [CrossRef]
- Lai, M.; Sun, X.; Chen, D.; Xie, Y.; Zhang, S. Age-related trends in genetic parameters for Larix kaempferi and their implications for early selection. BMC Genet. 2014, 15, S10. [Google Scholar] [CrossRef]
- Mckown, A.D.; Guy, R.D.; Klápště, J.; Geraldes, A.; Friedmann, M.; Cronk, Q.C.B.; El-Kassaby, Y.A.; Mansfield, S.D.; Douglas, C.J. Geographical and environmental gradients shape phenotypic trait variation and genetic structure in Populus trichocarpa. New Phytol. 2014, 201, 1263–1276. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Weng, Y.H.; Krasowski, M.; Yan, G.H.; Fullarton, M. Genetic parameters of growth and stem forking for black spruce progeny tested in New Brunswick, Canada. New For. 2018, 49, 265–277. [Google Scholar] [CrossRef]
- Cao, Y.; Deng, B.; Fang, S.; Shang, X.; Fu, X.; Yang, W. Genotypic variation in tree growth and selected flavonoids in leaves of Cyclocarya paliurus. South. For. 2018, 80, 67–74. [Google Scholar] [CrossRef]
- Kang, J.W.; Lee, H.; Lim, H.; Lee, W.Y. Identification of potential metabolic markers for the selection of a high-yield clone of Quercus acutissima in clonal seed orchard. Forests 2018, 9, 116. [Google Scholar] [CrossRef]
- King, D.J.; Gleadow, R.M.; Woodrow, I.E. The accumulation of terpenoid oils does not incur a growth cost in Eucalyptus polybractea seedlings. Funct. Plant Biol. 2006, 33, 497–505. [Google Scholar] [CrossRef]
- Maeda, H.; Dudareva, N. The shikimate pathway and aromatic amino acid biosynthesis in plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef]
- Wen, C.H.; Lin, S.S.; Chu, F.H. Transcriptome analysis of a subtropical deciduous tree: Autumn leaf senescence gene expression profile of formosan gum. Plant Cell Physiol. 2015, 56, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Chen, H.; Qin, Y.; Yang, Z.; Wang, T.; Wei, B. Physiological basis of Liquidambar formosana leaves during leaf color transformation in autumn. Guihaia 2021, 41, 2061–2068. [Google Scholar]
- Qi, S.; Zhao, R.; Yan, J.; Fan, Y.; Huang, C.; Li, H.; Chen, S.; Zhang, T.; Kong, L.; Zhao, J.; et al. Global transcriptome and coexpression network analyses reveal new insights into somatic embryogenesis in hybrid sweetgum (Liquidambar styraciflua × Liquidambar formosana). Front. Plant Sci. 2021, 12, 751866. [Google Scholar] [CrossRef]
- Zhao, X.; Xia, D.; Zeng, F.; Yao, S.; Shang, Y.; Zhang, G.; Wang, Y.; Zhang, T.; Zhan, Y. Provenances by sites interaction of growth traits and provenance selection of Fraxinus mandshurica. Sci. Silvae Sin. 2015, 51, 140–147. [Google Scholar]
- Liu, Y.; Xu, H.; Shang, F.; Jiao, H.; Zhang, L.; Luo, J.; Teng, W.; Liu, G. Variation and zoning of 16-year-old Betula platyphylla provenance. Sci. Silvae Sin. 2016, 52, 48–56. [Google Scholar]
- Hu, X.; Wu, F.; Sun, X.; Chen, H.; Yin, A.; Ji, K. Joint analysis of growth and wood property of 38-year-old Pinus massoniana from 55 provenance. J. Nanjing For. Univ. 2022, 46, 203–212. [Google Scholar]
- He, Q.; Fang, R.; Li, W.; Xia, Y.; Zhang, Y.; Shi, C.; Yang, S. Geographical variation of growth traits of Liquidambar formosana seedlings from different provenances. J. Plant Resour. Environ. 2019, 28, 88–95. [Google Scholar]
- Zhang, P.; Jin, G.; Zhou, Z.; Yu, L.; Fan, H. Provenance difference and geographic variation pattern for seedling trait of Schima superba. For. Res. 2004, 17, 192–198. [Google Scholar]
- Guerra, F.P.; Richards, J.H.; Fiehn, O.; Famula, R.; Stanton, B.J.; Shuren, R.; Sykes, R.; Davis, M.F.; Neale, D.B. Analysis of the genetic variation in growth, ecophysiology, and chemical and metabolomic composition of wood of Populus trichocarpa provenances. Tree Genet. Genomes 2016, 12, 6. [Google Scholar] [CrossRef]
- Chen, Z.Q.; Karlsson, B.; Mörling, T.; Olsson, L.; Mellerowicz, E.J.; Wu, H.X.; Lundqvist, S.O.; Gil, M.R.G. Genetic analysis of fiber dimensions and their correlation with stem diameter and solid-wood properties in Norway spruce. Tree Genet. Genomes 2016, 12, 123. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Provenance | Code | No. of Families | Longitude (°E) | Latitude (°N) | Altitude (m) | Tmean (°C) | Prec |
|---|---|---|---|---|---|---|---|
| Bawangling, Hainan | BWL | 20 | 109.04 | 19.22 | 1390 | 21.3 | 1657 |
| Cenxi, Guangxi | CX | 16 | 110.99 | 22.92 | 180 | 21.4 | 1450 |
| Chengbu, Hunan | CB | 17 | 110.32 | 26.39 | 1009 | 17.6 | 1600 |
| Fengdu, Chongqing | FD | 24 | 107.73 | 29.86 | 1044 | 18.3 | 1123 |
| Funing, Yunnan | FN | 24 | 105.63 | 23.63 | 679 | 19.8 | 1103 |
| Guangyuan, Sichuan | GY | 21 | 105.84 | 32.44 | 1225 | 16.1 | 950 |
| Huangshan, Anhui | HS | 23 | 118.34 | 29.72 | 1864 | 17.9 | 1697 |
| Huoshan, Anhui | HuoS | 17 | 116.33 | 31.39 | 406 | 15.2 | 1400 |
| Jianou, Fujian | JO | 23 | 118.31 | 27.02 | 608 | 19.3 | 1600 |
| Kaihua, Zhejiang | KH | 21 | 118.42 | 29.14 | 507 | 16.4 | 1830 |
| Kangxian, Gansu | KX | 18 | 105.61 | 33.33 | 1184 | 11.5 | 757 |
| Nanjing, Jiangsu | NJ | 20 | 118.77 | 32.04 | 17 | 16.2 | 1106 |
| Sangzhi, Hunan | SangZ | 23 | 110.16 | 29.40 | 1092 | 16.3 | 1447 |
| Songzi, Hubei | SZ | 21 | 111.77 | 30.17 | 145 | 16.5 | 1200 |
| Tiane, Guangxi | TE | 17 | 107.17 | 25.00 | 322 | 19.7 | 1078 |
| Tonggu, Jiangxi | TG | 14 | 114.37 | 28.52 | 433 | 16.4 | 1771 |
| Wengyuan, Guangdong | WY | 14 | 114.13 | 24.35 | 141 | 20.6 | 1778 |
| Wuzhishan, Hainan | WZS | 23 | 109.52 | 18.78 | 1867 | 22.4 | 1690 |
| Zhoushan, Zhejiang | ZS | 31 | 122.21 | 29.99 | 3 | 16.7 | 1222 |
| Trait | N | Mean ± SD | Minimum | Maximum | CVP (%) | CVG (%) | F Value | |
|---|---|---|---|---|---|---|---|---|
| Provenance | Family | |||||||
| SAC (%) | 967 | 2.03 ± 2.10 | 0.0 | 9.2 | 104.06 | 65.12 | 20.43 ** | 1.37 ** |
| HT (m) | 3342 | 6.30 ± 2.23 | 1.3 | 14.1 | 34.90 | 18.81 | 66.87 ** | 1.92 ** |
| DBH (cm) | 3339 | 6.46 ± 2.46 | 1.0 | 18.4 | 38.08 | 18.76 | 57.83 ** | 1.77 ** |
| VOL (dm3) | 3339 | 15.32 ± 15.64 | 0.1 | 164.8 | 101.65 | 42.28 | 59.14 ** | 1.55 ** |
| HCB (m) | 3336 | 2.17 ± 1.17 | 0.1 | 9.0 | 52.76 | 28.75 | 51.75 ** | 1.82 ** |
| LC | 2626 | 1.69 ± 0.95 | 1.0 | 5.0 | 32.69 | 17.59 | 15.82 ** | 1.56 ** |
| SSD | 3338 | 3.04 ± 1.15 | 1.0 | 5.0 | 21.34 | 10.40 | 15.28 ** | 1.57 ** |
| Trait | |||||||
|---|---|---|---|---|---|---|---|
| SAC | 1.1055 | 0.4369 | 2.9206 | 4.4630 | 0.53 (0.11) | 0.31 (0.10) | 0.39 (0.15) |
| HT | 1.3711 | 0.3516 | 3.1203 | 4.8431 | 0.57 (0.09) | 0.25 (0.04) | 0.29 (0.06) |
| DBH | 1.4477 | 0.3666 | 4.2281 | 6.0424 | 0.51 (0.09) | 0.21 (0.04) | 0.24 (0.05) |
| VOL | 61.7009 | 10.4925 | 170.4696 | 242.6630 | 0.52 (0.09) | 0.16 (0.03) | 0.17 (0.04) |
| HCB | 0.2931 | 0.0970 | 0.9165 | 1.3067 | 0.49 (0.09) | 0.24 (0.03) | 0.30 (0.05) |
| LC | 0.0014 | 0.0013 | 0.0150 | 0.0177 | 0.22 (0.07) | 0.20 (0.04) | 0.29 (0.06) |
| SSD | 0.0011 | 0.0009 | 0.0138 | 0.0158 | 0.19 (0.06) | 0.17 (0.03) | 0.24 (0.05) |
| Trait | SAC | HT | DBH | VOL | HCB | LC | SSD |
|---|---|---|---|---|---|---|---|
| SAC | −0.739 ** | −0.816 ** | −0.734 ** | −0.494 ** | −0.940 ** | 0.102 | |
| HT | −0.367 ** | 0.942 ** | 0.977 ** | 0.816 ** | 0.731 ** | −0.177 | |
| DBH | −0.401 ** | 0.844 ** | 0.982 ** | 0.705 ** | 0.835 ** | 0.147 | |
| VOL | −0.358 ** | 0.870 ** | 0.932 ** | 0.772 ** | 0.766 ** | 0.106 | |
| HCB | −0.226 ** | 0.543 ** | 0.347 ** | 0.400 ** | 0.339 * | −0.212 | |
| LC | −0.368 ** | 0.177 ** | 0.167 ** | 0.128 ** | 0.123 ** | 0.090 | |
| SSD | 0.032 | −0.408 ** | −0.293 ** | −0.293 ** | −0.143 ** | −0.039 |
| Trait | Top 5% Provenance | Top 15% Provenance | Top 5% Family | Top 15% Family | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | ΔG (%) | Mean | ΔG (%) | Mean | ΔG (%) | Mean | ΔG (%) | |
| SAC | 4.09 | 103.94 | 3.71 | 85.14 | 5.35 | 163.57 | 4.42 | 117.71 |
| HT | 8.51 | 34.25 | 8.29 | 30.83 | 8.31 | 32.52 | 7.69 | 22.71 |
| DBH | 8.47 | 30.92 | 8.29 | 28.27 | 8.52 | 33.15 | 7.96 | 24.37 |
| VOL | 30.44 | 96.33 | 28.62 | 84.59 | 33.22 | 121.51 | 27.58 | 83.90 |
| HCB | 3.38 | 54.47 | 3.15 | 44.04 | 3.69 | 71.70 | 3.15 | 46.41 |
| LC | 2.02 | 20.58 | 2.02 | 20.26 | 2.61 | 56.28 | 2.42 | 44.75 |
| SSD a | 2.44 | 19.94 | 2.60 | 14.41 | 2.18 | 28.34 | 2.30 | 24.32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, M.; Zhou, L.; Yu, N.; Li, R.; Wu, S.; Yang, J.; Su, J. Genetic Parameters and Selection Responses for Important Breeding Traits in Liquidambar formosana Based on a Provenance–Family Trial. Forests 2023, 14, 2293. https://doi.org/10.3390/f14122293
Dong M, Zhou L, Yu N, Li R, Wu S, Yang J, Su J. Genetic Parameters and Selection Responses for Important Breeding Traits in Liquidambar formosana Based on a Provenance–Family Trial. Forests. 2023; 14(12):2293. https://doi.org/10.3390/f14122293
Chicago/Turabian StyleDong, Mingliang, Li Zhou, Niu Yu, Rongsheng Li, Shijun Wu, Jinchang Yang, and Jianyu Su. 2023. "Genetic Parameters and Selection Responses for Important Breeding Traits in Liquidambar formosana Based on a Provenance–Family Trial" Forests 14, no. 12: 2293. https://doi.org/10.3390/f14122293
APA StyleDong, M., Zhou, L., Yu, N., Li, R., Wu, S., Yang, J., & Su, J. (2023). Genetic Parameters and Selection Responses for Important Breeding Traits in Liquidambar formosana Based on a Provenance–Family Trial. Forests, 14(12), 2293. https://doi.org/10.3390/f14122293