
DELLA-Mediated Gibberellin Acid Participation in the Regulation of Dormancy and Growth of Fraxinus mandshurica Rupr.

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture Conditions
2.1.1. Seed Germination
2.1.2. Culture Methods and Conditions
2.1.3. Transplanting Process
2.2. Extraction of Endogenous Hormones
2.3. Sample Preparation for Transcriptional Profiling
2.3.1. Sample for Dormancy-Related Genes
2.3.2. Sample for FmDELLA Genes for Different Months, Tissues, and Developmental Stages
2.3.3. Effects of Different Exogenous Hormone Signals on FmDELLA Genes
2.3.4. Effects of Exogenous GA on Top Bud Dormancy Release in Transplanted Seedlings
2.4. RNA Extraction and cDNA Library Preparation
- (1)
- A total of 650 μL Tris CTAB and 50 μL β-Mercaptoethanol were added to a 1.5 mL sterile centrifuge tube, and the extraction buffer was preheated at 65 °C (aqueous solution).
- (2)
- The mortar, rod, and spoon were thoroughly cooled with liquid nitrogen three–four times, and 0.1–0.5 g of blades (−40 °C) were put into liquid nitrogen and ground to powder.
- (3)
- The prepared powder was added to the preheated Tris-CTAB extraction buffer, subjected to intense vortex oscillation for 30 s, and a water bath at 65 °C for 15 min.
- (4)
- A total of 700 μL of chloroform was added to a centrifuge tube, which was centrifuged for 10 min at 4 °C.
- (5)
- Extract the supernatant and repeat step 4 twice.
- (6)
- Extract the supernatant and add 560 μL lithium chloride (5 M), 350 μL anhydrous ethanol, and 60 μL sodium acetate and stand on ice for 60–90 min.
- (7)
- Centrifuge (12,000 rmp) at a low temperature (4 °C) for 10 min; extract the supernatant and discard it.
- (8)
- The sediment was rinsed twice with 70% ethanol; the centrifuge tube was opened in the fume hood to air dry the sediment, and 20 μL ddH2O was added to the centrifuge tube.
2.5. Reverse Transcriptional Quantitative PCR(RT-qPCR)
2.6. Statistical Analysis
3. Results
3.1. Germination Characteristics of the F. mandshurica Embryo
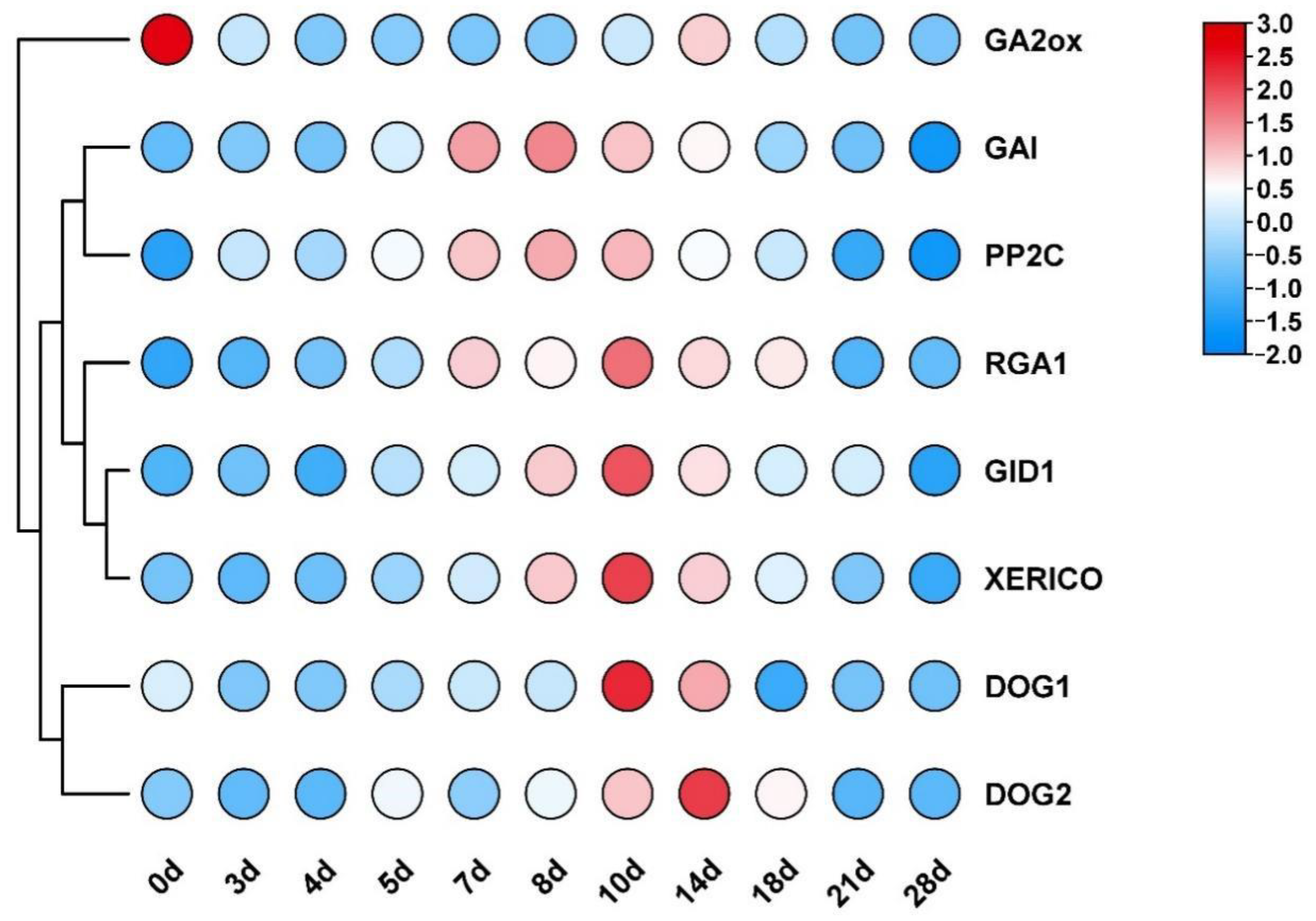
3.2. Variations in the Transcription Levels of Genes Related to GA and ABA Signaling Pathways and DOG Genes during Seed Development
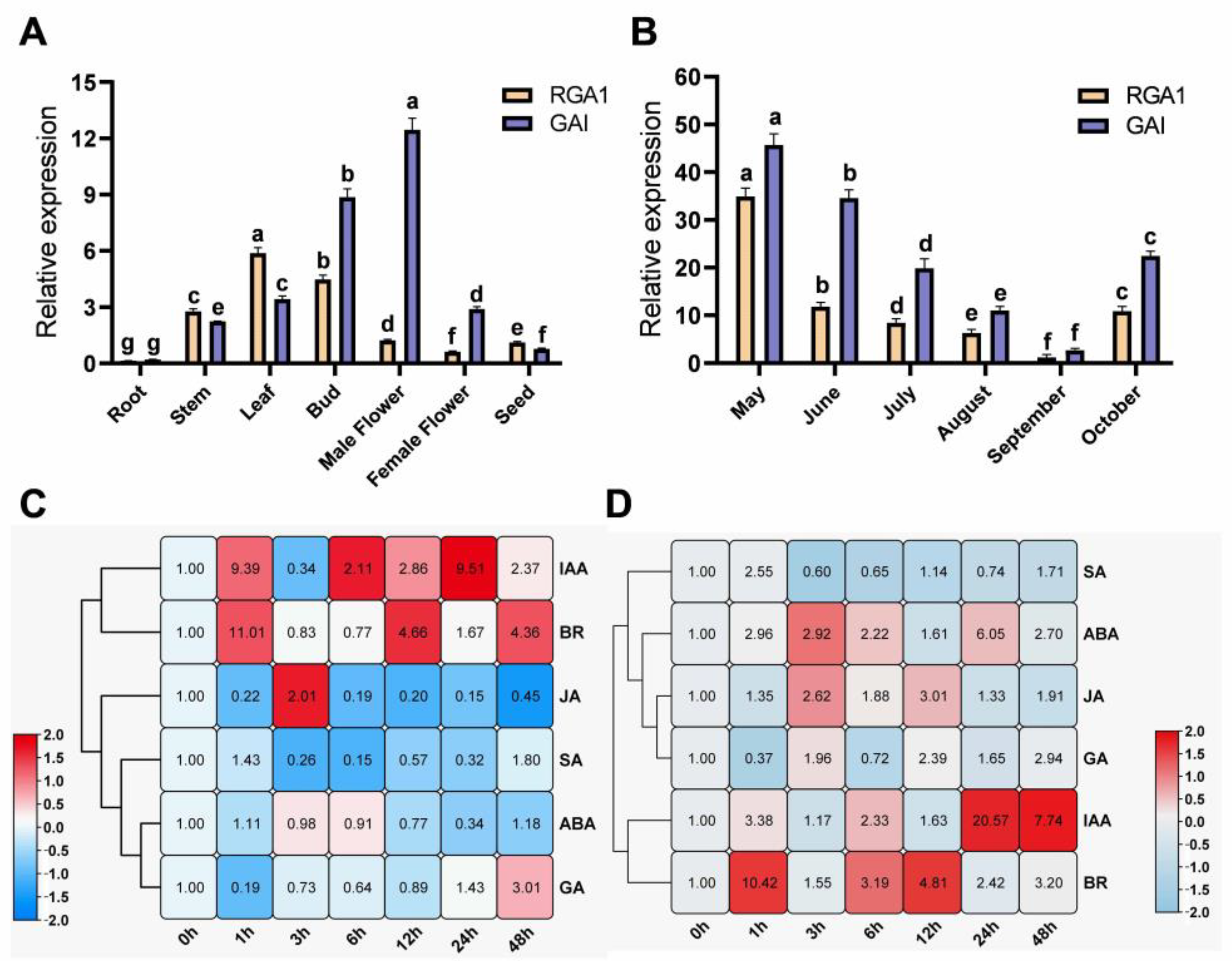
3.3. Spatiotemporal Transcriptional Patterns of FmDELLA Genes
3.4. Effect of Exogenous Hormone Signals on the Transcriptional Levels of FmDELLA Genes
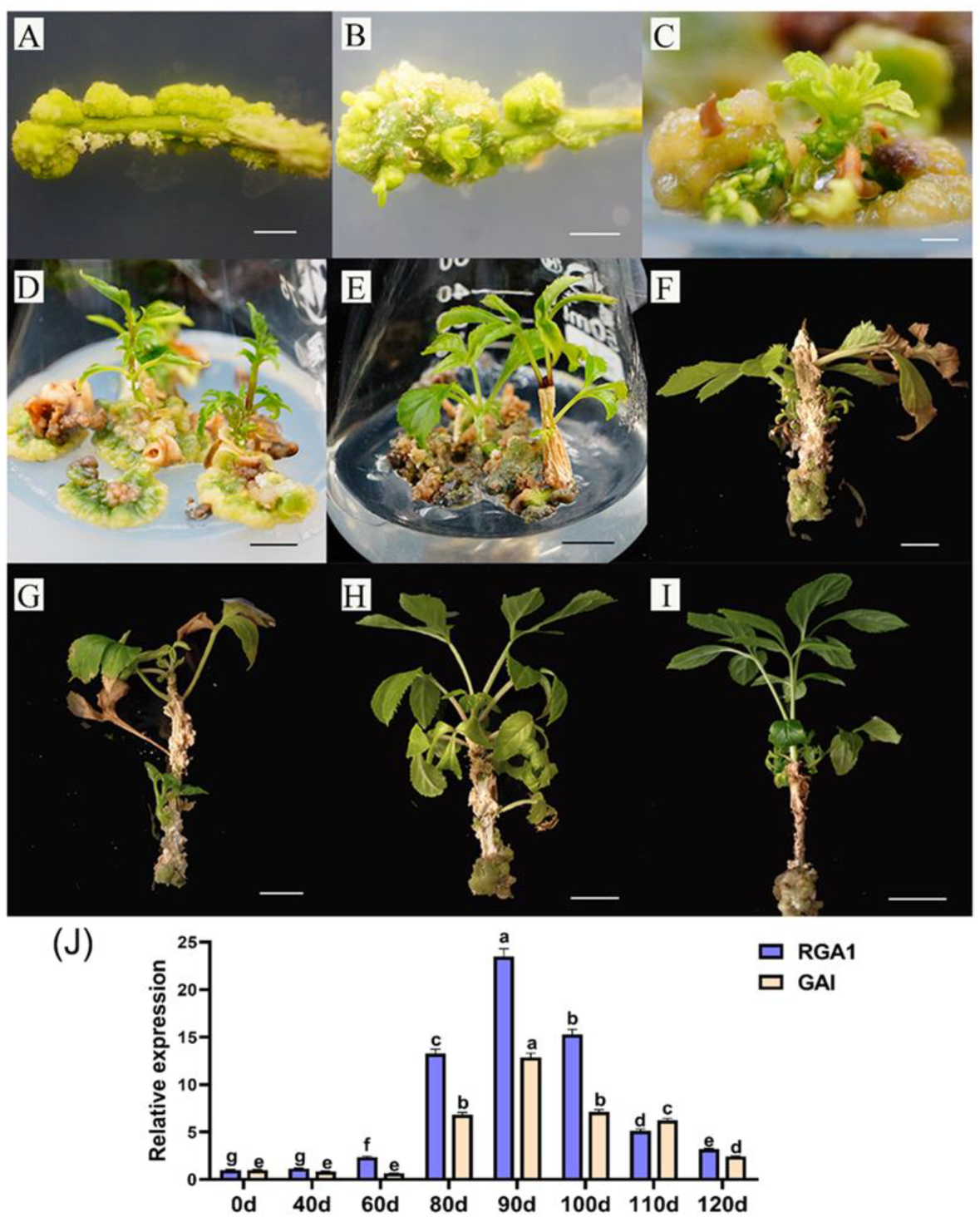
3.5. Variation Analysis of FmDELLA Genes’ Transcriptional Levels at Different Developmental Stages of Tissue Culture Seedlings
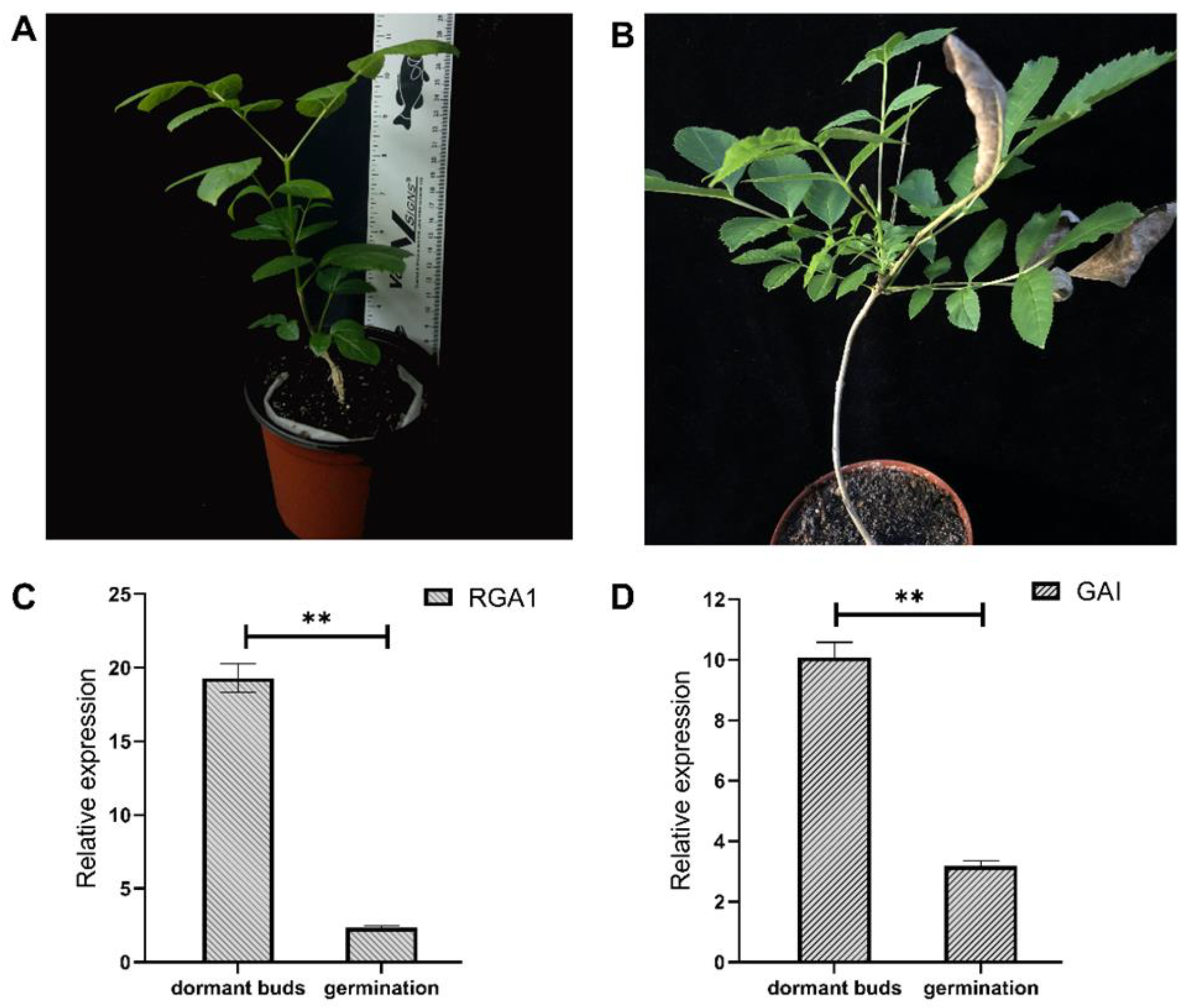
3.6. Variation in the Transcriptional Level of the FmDELLA Gene in the Dormant Top Buds of Transplanted F. mandshurica Seedlings and Top Bud Dormancy Release
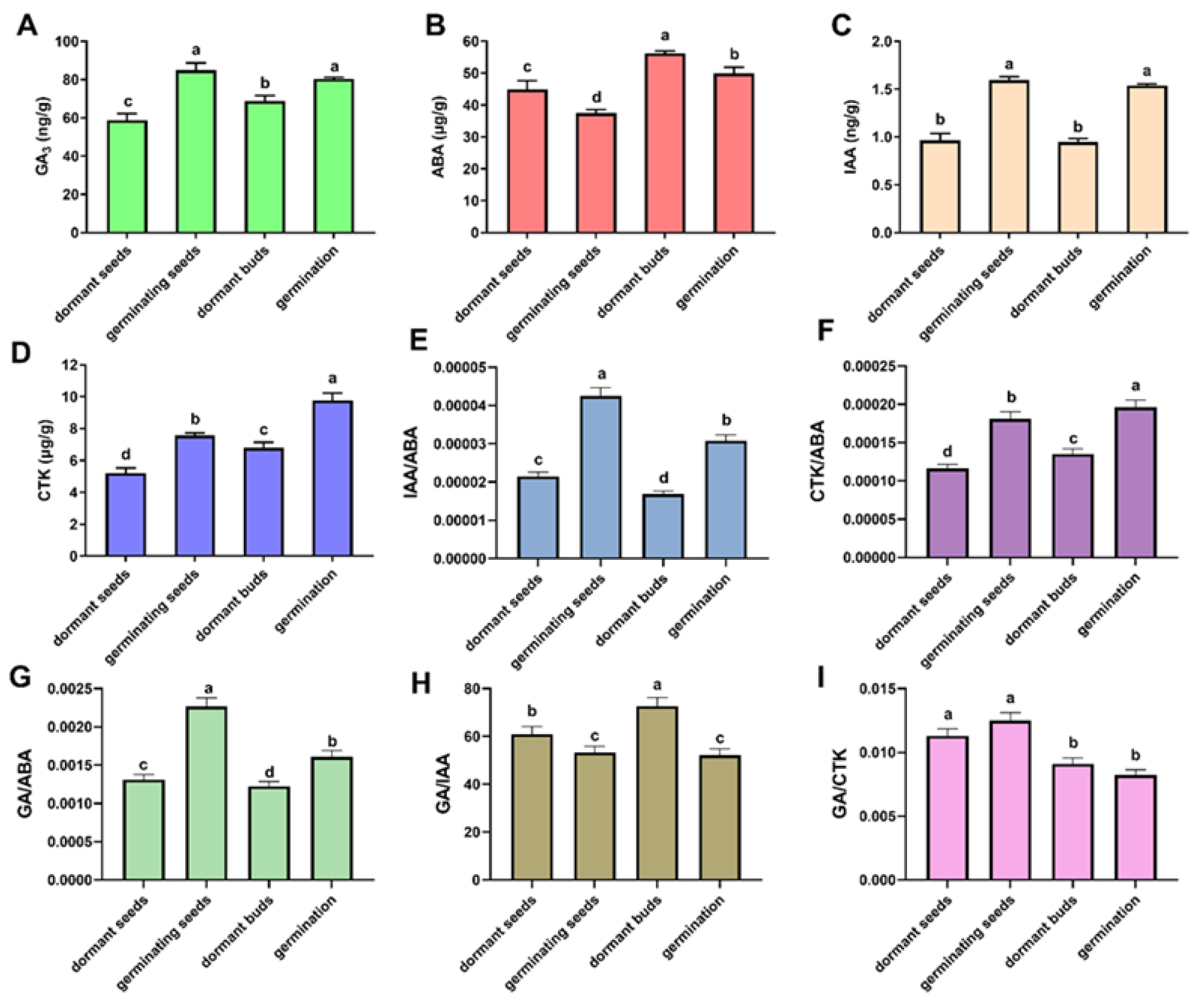
3.7. Relationship between Endogenous Hormones and Dormancy
4. Discussion
4.1. Variations in the Transcriptional Level of Genes Related to GA and ABA Signaling Pathways and DOG Genes during Seed Development
4.2. The Function of FmDELLA in the Passive Dormancy of F. mandshurica
4.3. The Spatiotemporal Transcriptional Expression Patterns of DELLA
4.4. Spraying Gibberellin Can Break the Dormancy of Apical Buds
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bassel, G.W. To Grow or not to Grow? Trends Plant Sci. 2016, 21, 498–505. [Google Scholar] [CrossRef] [PubMed]
- El Yaacoubi, A.; Malagi, G.; Oukabli, A.; Citadin, I.; Hafidi, M.; Bonhomme, M.; Legave, J.M. Differentiated dynamics of bud dormancy and growth in temperate fruit trees relating to bud phenology adaptation, the case of apple and almond trees. Int. J. Biometeorol. 2016, 60, 1695–1710. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, Q.; Wen, B.; Zhang, R.; Jing, X.; Xiao, W.; Chen, X.; Tan, Q.; Li, L. Endodormancy Release Can Be Modulated by the GA(4)-GID1c-DELLA2 Module in Peach Leaf Buds. Front. Plant Sci. 2021, 12, 713514. [Google Scholar] [CrossRef] [PubMed]
- Howe, G.T.; Horvath, D.P.; Dharmawardhana, P.; Priest, H.D.; Mockler, T.C.; Strauss, S.H. Extensive Transcriptome Changes During Natural Onset and Release of Vegetative Bud Dormancy in Populus. Front. Plant Sci. 2015, 6, 989. [Google Scholar] [CrossRef] [PubMed]
- Horvath, D. Common mechanisms regulate flowering and dormancy. Plant Sci. 2009, 177, 523–531. [Google Scholar] [CrossRef]
- De Vleesschauwer, D.; Seifi, H.S.; Filipe, O.; Haeck, A.; Huu, S.N.; Demeestere, K.; Hofte, M. The DELLA Protein SLR1 Integrates and Amplifies Salicylic Acid- and Jasmonic Acid-Dependent Innate Immunity in Rice. Plant Physiol. 2016, 170, 1831–1847. [Google Scholar] [CrossRef]
- Gao, L.; Niu, D.; Chi, T.; Yuan, Y.; Liu, C.; Gai, S.; Zhang, Y. PsRGL1 negatively regulates chilling and gibberellin induced dormancy release by PsF box1 mediated targeting for proteolytic degradation in tree peony. Hortic. Res. 2023, 10, uhad044. [Google Scholar] [CrossRef]
- Daviere, J.M.; Wild, M.; Regnault, T.; Baumberger, N.; Eisler, H.; Genschik, P.; Achard, P. Class I TCP-DELLA interactions in inflorescence shoot apex determine plant height. Curr. Biol. 2014, 24, 1923–1928. [Google Scholar] [CrossRef]
- Hauvermale, A.L.; Ariizumi, T.; Steber, C.M. Gibberellin signaling: A theme and variations on DELLA repression. Plant Physiol. 2012, 160, 83–92. [Google Scholar] [CrossRef]
- Leone, M.; Keller, M.M.; Cerrudo, I.; Ballare, C.L. To grow or defend? Low red: Far-red ratios reduce jasmonate sensitivity in Arabidopsis seedlings by promoting DELLA degradation and increasing JAZ10 stability. N. Phytol. 2014, 204, 355–367. [Google Scholar] [CrossRef]
- Xu, F.; Li, T.; Xu, P.B.; Li, L.; Du, S.S.; Lian, H.L.; Yang, H.Q. DELLA proteins physically interact with CONSTANS to regulate flowering under long days in Arabidopsis. FEBS Lett. 2016, 590, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; An, F.; Li, W.; Ma, M.; Feng, Y.; Zhang, X.; Guo, H. DELLA proteins interact with FLC to repress flowering transition. J. Integr. Plant Biol. 2016, 58, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Hirano, K.; Sato, T.; Mitsuda, N.; Nomoto, M.; Maeo, K.; Koketsu, E.; Mitani, R.; Kawamura, M.; Ishiguro, S.; et al. DELLA protein functions as a transcriptional activator through the DNA binding of the indeterminate domain family proteins. Proc. Natl. Acad. Sci. USA 2014, 111, 7861–7866. [Google Scholar] [CrossRef] [PubMed]
- Fonouni-Farde, C.; Tan, S.; Baudin, M.; Brault, M.; Wen, J.; Mysore, K.S.; Niebel, A.; Frugier, F.; Diet, A. DELLA-mediated gibberellin signalling regulates Nod factor signalling and rhizobial infection. Nat. Commun. 2016, 7, 12636. [Google Scholar] [CrossRef] [PubMed]
- Bian, T.T.; Ma, Y.; Guo, J.; Wu, Y.; Shi, D.M.; Guo, X.F. Herbaceous peony (Paeonia lactiflora Pall.) PlDELLA gene negatively regulates dormancy release and plant growth. Plant Sci. 2020, 297, 110539. [Google Scholar] [CrossRef] [PubMed]
- Veerabagu, M.; van der Schoot, C.; Tureckova, V.; Tarkowska, D.; Strnad, M.; Rinne, P.L.H. Light on perenniality: Para-dormancy is based on ABA-GA antagonism and endo-dormancy on the shutdown of GA biosynthesis. Plant Cell Environ. 2023, 46, 1785–1804. [Google Scholar] [CrossRef]
- Sun, T.P. Gibberellin-GID1-DELLA: A pivotal regulatory module for plant growth and development. Plant Physiol. 2010, 154, 567–570. [Google Scholar] [CrossRef]
- Zheng, Y.; Gao, Z.; Zhu, Z. DELLA-PIF Modules: Old Dogs Learn New Tricks. Trends Plant Sci. 2016, 21, 813–815. [Google Scholar] [CrossRef]
- Zentella, R.; Zhang, Z.L.; Park, M.; Thomas, S.G.; Endo, A.; Murase, K.; Fleet, C.M.; Jikumaru, Y.; Nambara, E.; Kamiya, Y.; et al. Global analysis of della direct targets in early gibberellin signaling in Arabidopsis. Plant Cell 2007, 19, 3037–3057. [Google Scholar] [CrossRef]
- Yu, L.; Li, X.; Tian, H.; Liu, H.; Xiao, Y.; Liang, N.; Zhao, X.; Zhan, Y. Effects of Hormones and Epigenetic Regulation on the Callus and Adventitious Bud Induction of Fraxinus mandshurica Rupr. Forests 2020, 11, 590. [Google Scholar] [CrossRef]
- Graeber, K.; Linkies, A.; Steinbrecher, T.; Mummenhoff, K.; Tarkowská, D.; Turečková, V.; Ignatz, M.; Sperber, K.; Voegele, A.; de Jong, H.; et al. DELAY OF GERMINATION 1 mediates a conserved coat-dormancy mechanism for the temperature- and gibberellin-dependent control of seed germination. Proc. Natl. Acad. Sci. USA 2014, 111, E3571–E3580. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, J.; Palusinska, M.; Wroblewska-Swiniarska, A.; Pietras, Z.; Szewc, L.; Dolata, J.; Jarmolowski, A.; Swiezewski, S. Alternative Polyadenylation of the Sense Transcript Controls Antisense Transcription of DELAY OF GERMINATION 1 in Arabidopsis. Mol. Plant 2017, 10, 1349–1352. [Google Scholar] [CrossRef] [PubMed]
- Obroucheva, N.V. Distinct regulatory patterns of seed dormancy release and germination commencement. Seed Sci. Technol. 2010, 38, 265–279. [Google Scholar] [CrossRef]
- Obroucheva, N.V. Transition from hormonal to nonhormonal regulation as exemplified by seed dormancy release and germination triggering. Russ. J. Plant Physiol. 2012, 59, 546–555. [Google Scholar] [CrossRef]
- Nishimura, N.; Tsuchiya, W.; Moresco, J.J.; Hayashi, Y.; Satoh, K.; Kaiwa, N.; Irisa, T.; Kinoshita, T.; Schroeder, J.I.; Yates, J.R., 3rd; et al. Control of seed dormancy and germination by DOG1-AHG1 PP2C phosphatase complex via binding to heme. Nat. Commun. 2018, 9, 2132. [Google Scholar] [CrossRef]
- Hou, X.; Lee, L.Y.; Xia, K.; Yan, Y.; Yu, H. DELLAs modulate jasmonate signaling via competitive binding to JAZs. Dev. Cell 2010, 19, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Josse, E.M.; Gan, Y.B.; Bou-Torrent, J.; Stewart, K.L.; Gilday, A.D.; Jeffree, C.E.; Vaistij, F.E.; Martinez-Garcia, J.F.; Nagy, F.; Graham, I.A.; et al. A DELLA in Disguise: SPATULA Restrains the Growth of the Developing Arabidopsis Seedling. Plant Cell 2011, 23, 1337–1351. [Google Scholar] [CrossRef] [PubMed]
- Ariizumi, T.; Hauvermale, A.L.; Nelson, S.K.; Hanada, A.; Yamaguchi, S.; Steber, C.M. Lifting della repression of Arabidopsis seed germination by nonproteolytic gibberellin signaling. Plant Physiol. 2013, 162, 2125–2139. [Google Scholar] [CrossRef]
- Penfield, S.; Gilday, A.D.; Halliday, K.J.; Graham, I.A. DELLA-mediated cotyledon expansion breaks coat-imposed seed dormancy. Curr. Biol. 2006, 16, 2366–2370. [Google Scholar] [CrossRef]
- Yu, H.; Ito, T.; Zhao, Y.; Peng, J.; Kumar, P.; Meyerowitz, E.M. Floral homeotic genes are targets of gibberellin signaling in flower development. Pans 2004, 101, 7827–7832. [Google Scholar] [CrossRef]
- Cheng, H.; Qin, L.; Lee, S.; Fu, X.; Richards, D.E.; Cao, D.; Luo, D.; Harberd, N.P.; Peng, J. Gibberellin regulates Arabidopsis floral development via suppression of DELLA protein function. Development 2004, 131, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Plackett, A.R.G.; Ferguson, A.C.; Powers, S.J.; Wanchoo-Kohli, A.; Phillips, A.L.; Wilson, Z.A.; Hedden, P.; Thomas, S.G. DELLA activity is required for successful pollen development in the Columbia ecotype of Arabidopsis. N. Phytol. 2014, 201, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, R.; Zhu, K.M.; Jiang, T.; Ding, P.; Gao, Y.; Tan, X.L. DELLAs directed gibberellins responses orchestrate crop development: A brief review. Crop Sci. 2022, 63, 1–28. [Google Scholar] [CrossRef]
- Fonouni-Farde, C.; Diet, A.; Frugier, F. Root Development and Endosymbioses: DELLAs Lead the Orchestra. Trends Plant Sci. 2016, 21, 898–900. [Google Scholar] [CrossRef] [PubMed]
- Ubeda-Tomas, S.; Swarup, R.; Coates, J.; Swarup, K.; Laplaze, L.; Beemster, G.T.; Hedden, P.; Bhalerao, R.; Bennett, M.J. Root growth in Arabidopsis requires gibberellin/DELLA signalling in the endodermis. Nat. Cell Biol. 2008, 10, 625–628. [Google Scholar] [CrossRef]
- Daviere, J.M.; Achard, P. A Pivotal Role of DELLAs in Regulating Multiple Hormone Signals. Mol. Plant 2016, 9, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, R.R.; Lynch, T.J. Overexpression of ABI5 Binding Proteins Suppresses Inhibition of Germination Due to Overaccumulation of DELLA Proteins. Int. J. Mol. Sci. 2022, 23, 5537. [Google Scholar] [CrossRef]
- He, L.; Zhang, J.; Guo, D.; Tian, H.; Cao, Y.; Ji, X.; Zhan, Y. Establishment of the technology of cambial meristematic cells (CMCs) culture from shoots and high expression of FmPHV (PHAVOLUTA) functions in identification and differentiation of CMCs and promoting the shoot regeneration by hypocotyl in Fraxinus mandshurica. Plant Physiol. Biochem. 2021, 160, 352–364. [Google Scholar] [CrossRef]
- Cui, J.W.; Zhao, J.G.; Zhao, J.Y.; Xu, H.M.; Wang, L.; Jin, B. Cytological and miRNA expression changes during the vascular cambial transition from the dormant stage to the active stage in Ginkgo biloba L. Trees-Struct. Funct. 2016, 30, 2177–2188. [Google Scholar] [CrossRef]
- Gao, J.; Ni, X.P.; Li, H.T.; Hayat, F.; Shi, T.; Gao, Z.H. miR169 and PmRGL2 synergistically regulate the NF-Y complex to activate dormancy release in Japanese apricot (Prunus mume Sieb. et Zucc.). Plant Mol. Biol. 2021, 105, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Artlip, T.; McDermaid, A.; Ma, Q.; Wisniewski, M. Differential gene expression in non-transgenic and transgenic “M.26” apple overexpressing a peach CBF gene during the transition from eco-dormancy to bud break. Hortic. Res. 2019, 6, 86. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Chu, C. Gibberellin Metabolism and Signaling: Targets for Improving Agronomic Performance of Crops. Plant Cell Physiol. 2020, 61, 1902–1911. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Explant Source | Stem | Adventitious Bud | Secondary Proliferation |
|---|---|---|---|
| Basic Medium Type | MSB5 | MSB5 | WPM |
| Sucrose (g/L) | 30 | 30 | 20 |
| Agar (g/L) | 7 | 7 | 7 |
| 6-BA (mg/L) | 5 | 5 | 8 |
| TDZ (mg/L) | 8 | 8 | |
| glycine (mg/L) | 2 | 2 | |
| IBA (mg/L) | 0.1 | 0.1 | |
| coconut water | 5% | 5% | |
| TSA (mg/L) | 15 | 15 |
| Concentration (mg/L) | Germination Rate of New Buds (%) |
|---|---|
| 0 | 11.67 e |
| 30 | 35.00 d |
| 50 | 55.00 c |
| 100 | 90.00 a |
| 300 | 76.67 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Yu, L.; Li, C.; Liang, N.; Zeng, F.; Zhan, Y. DELLA-Mediated Gibberellin Acid Participation in the Regulation of Dormancy and Growth of Fraxinus mandshurica Rupr. Forests 2023, 14, 2128. https://doi.org/10.3390/f14112128
Zhao X, Yu L, Li C, Liang N, Zeng F, Zhan Y. DELLA-Mediated Gibberellin Acid Participation in the Regulation of Dormancy and Growth of Fraxinus mandshurica Rupr. Forests. 2023; 14(11):2128. https://doi.org/10.3390/f14112128
Chicago/Turabian StyleZhao, Xingtang, Lei Yu, Chuanzhou Li, Nansong Liang, Fansuo Zeng, and Yaguang Zhan. 2023. "DELLA-Mediated Gibberellin Acid Participation in the Regulation of Dormancy and Growth of Fraxinus mandshurica Rupr." Forests 14, no. 11: 2128. https://doi.org/10.3390/f14112128
APA StyleZhao, X., Yu, L., Li, C., Liang, N., Zeng, F., & Zhan, Y. (2023). DELLA-Mediated Gibberellin Acid Participation in the Regulation of Dormancy and Growth of Fraxinus mandshurica Rupr. Forests, 14(11), 2128. https://doi.org/10.3390/f14112128