
Identification of CpbZIP11 in Cyclocarya paliurus Involved in Environmental Stress Responses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Stress Treatment
2.2. Differential Expression Analysis
2.3. Classification and Sequence Analysis of bZIP Members
2.4. CpbZIP11 mRNA Secondary Structure Prediction
2.5. mRNA Splicing Assay and RT–PCR
2.6. Subcellular Localization Analysis
2.7. Transcriptional Activation Activity Assay in Yeast
3. Results
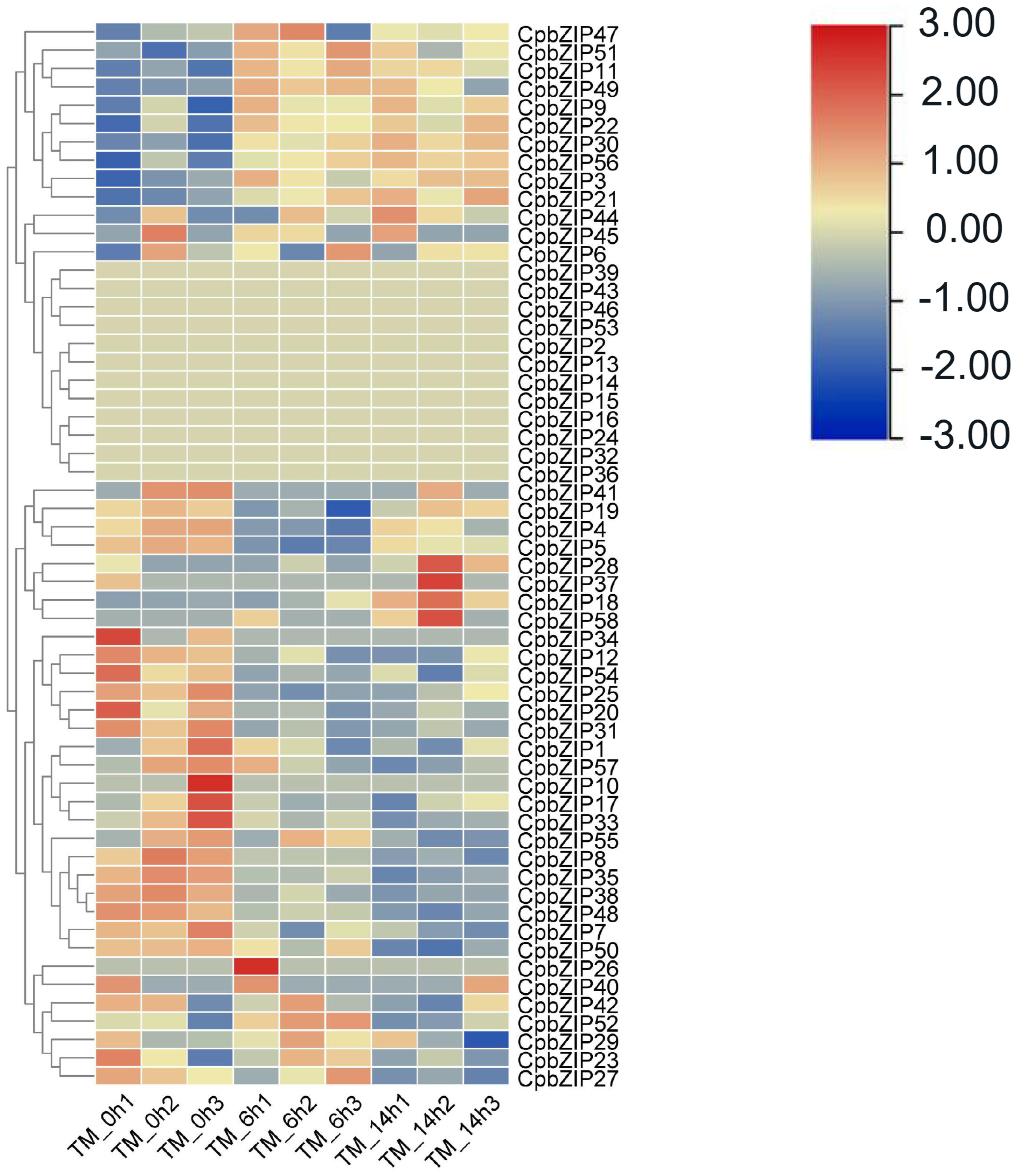
3.1. The Expression Profile of CpbZIP Genes under ER Stress
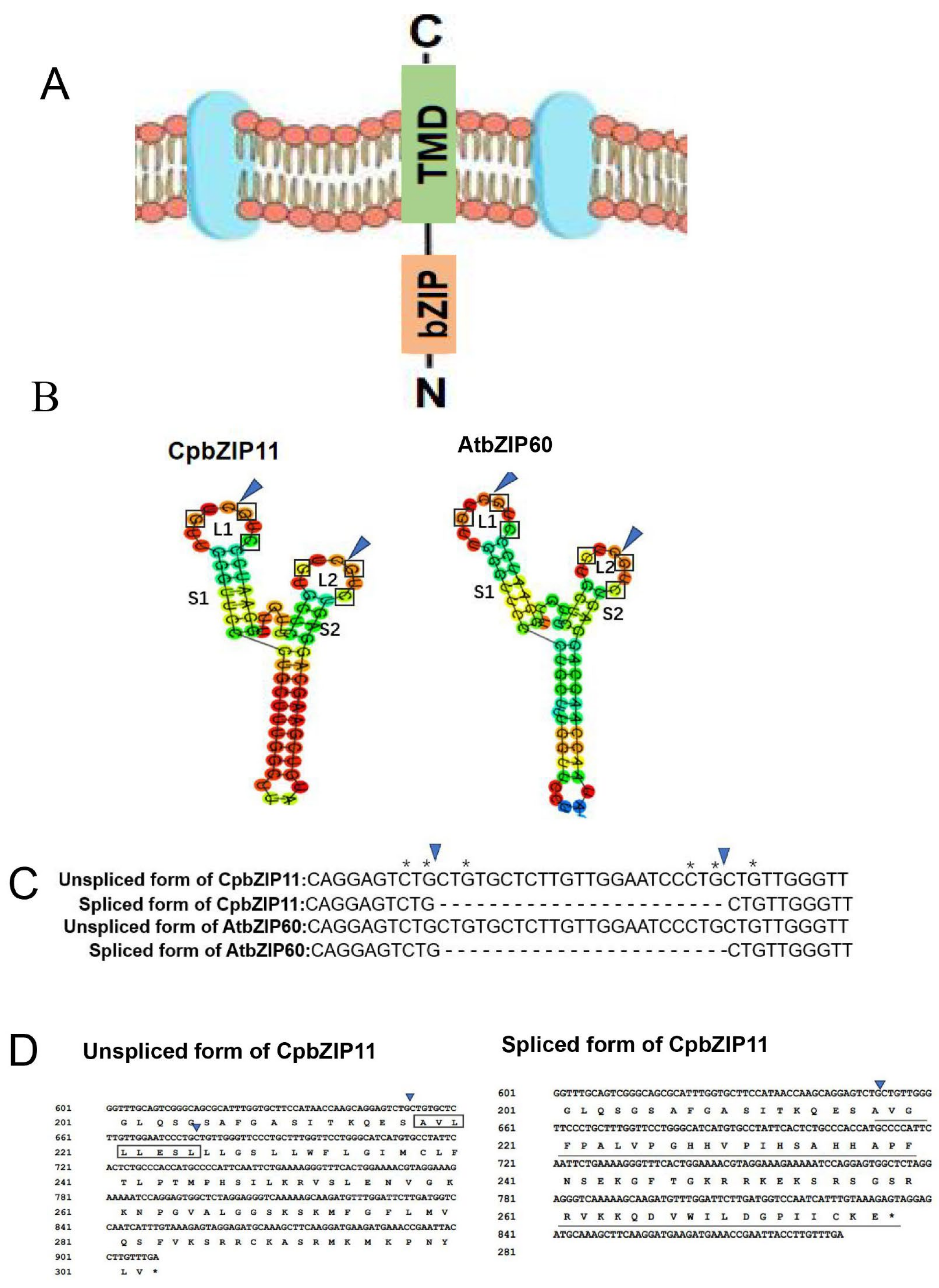
3.2. CpbZIP11 Sequence Homology with AtbZIP60 and Splicing Site Prediction
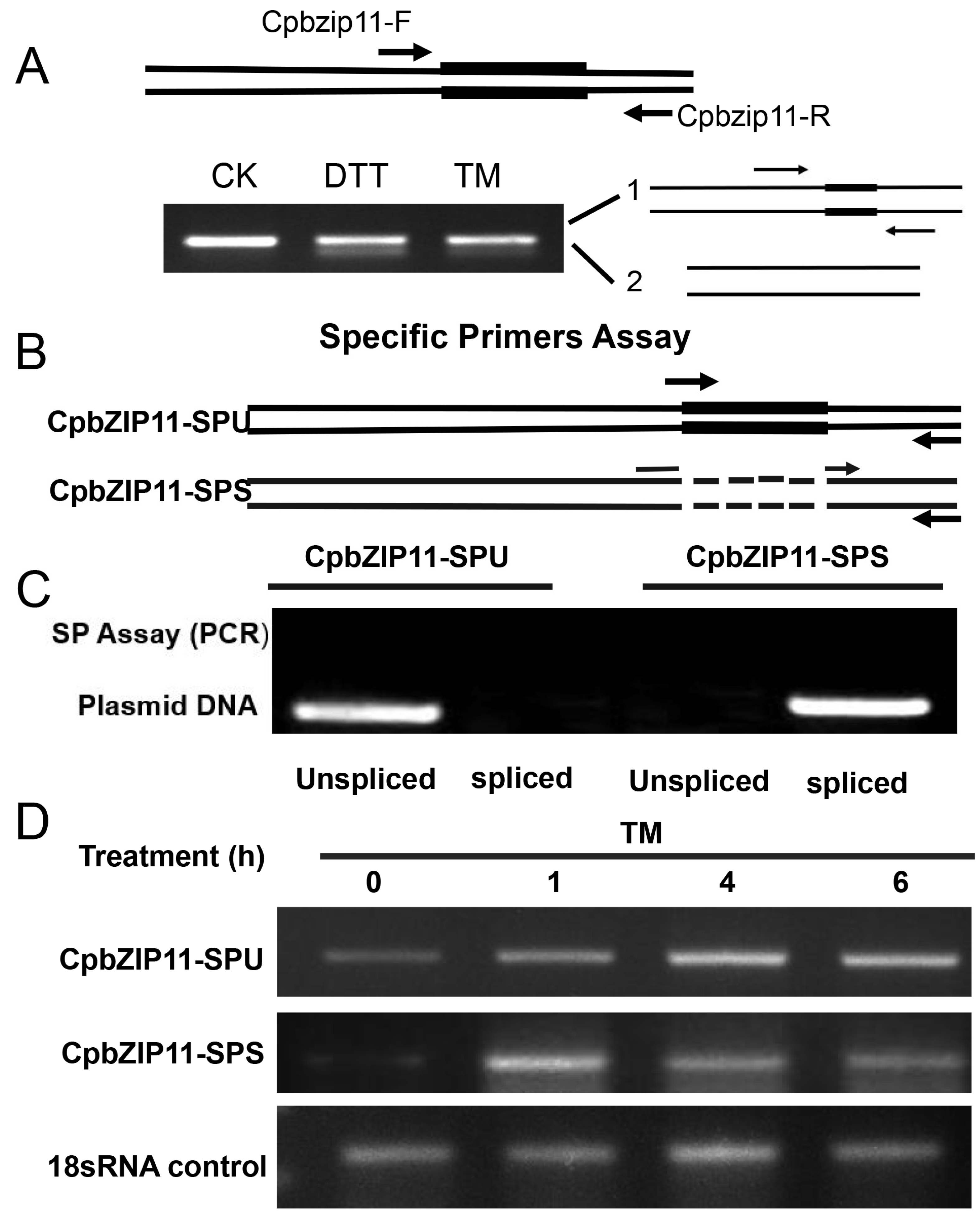
3.3. CpbZIP11 mRNA Splicing Produces the Active Form of Transcription Factor bZIP11
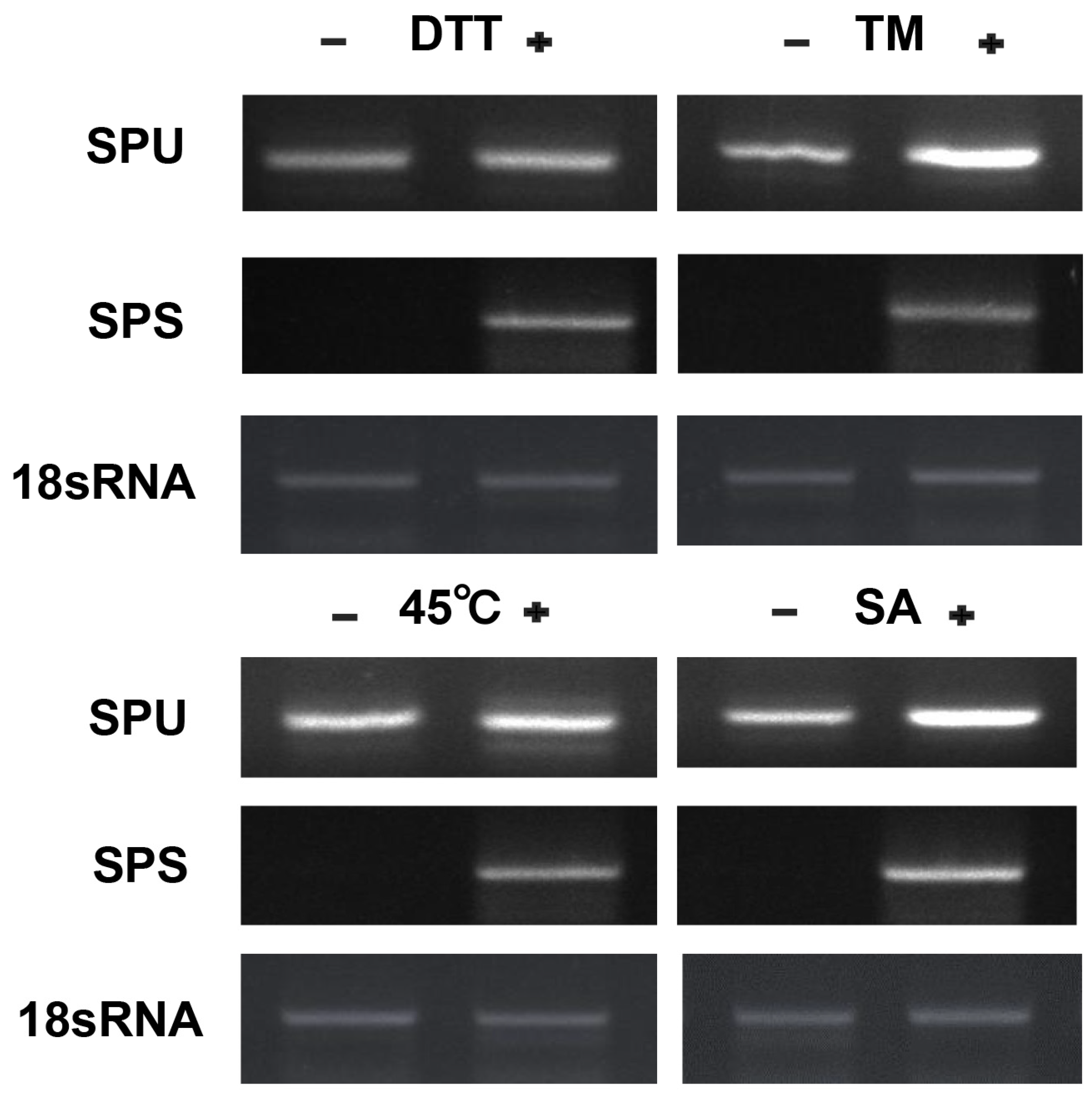
3.4. CpbZIP11 Is Induced by Environmental Stresses
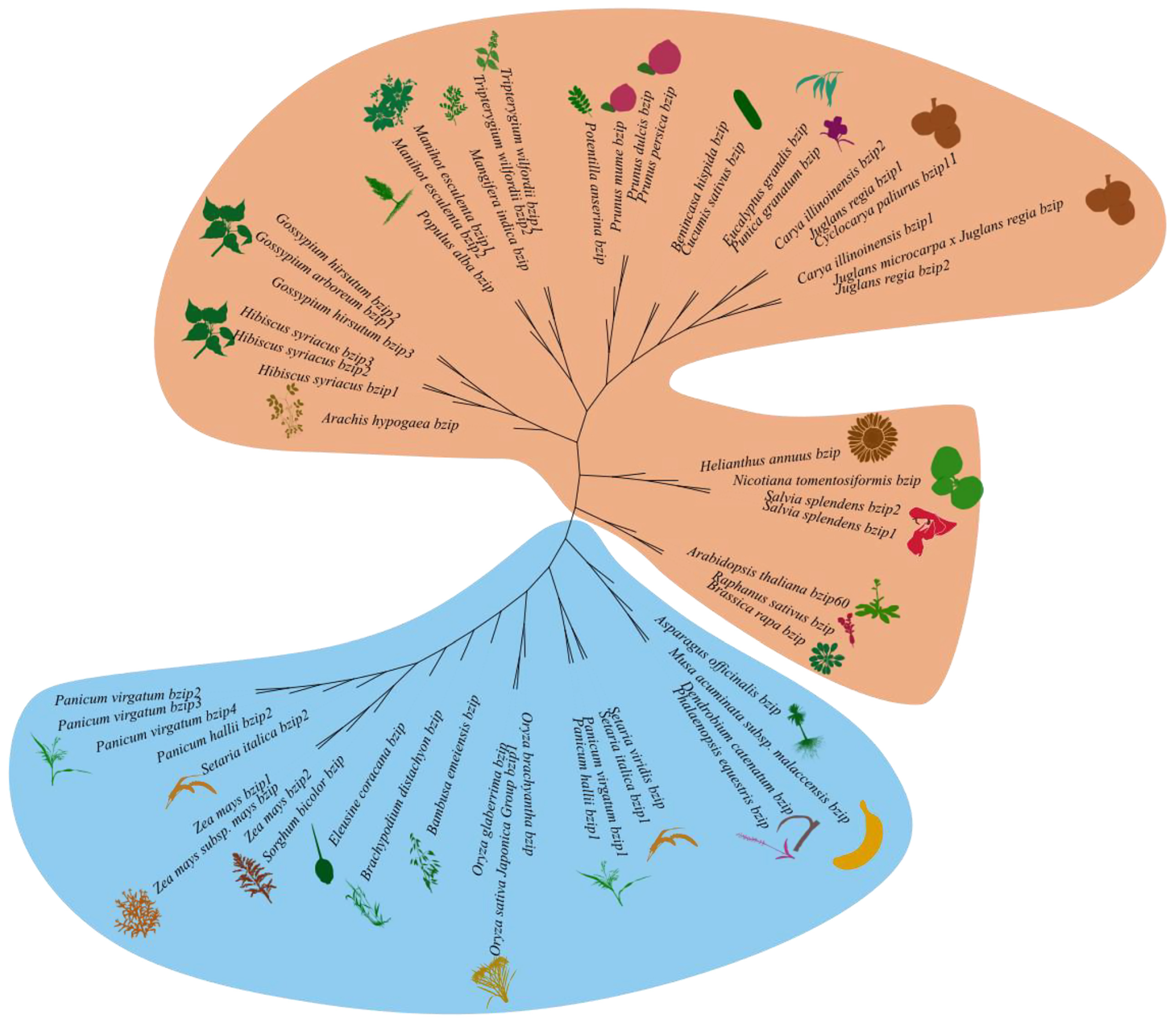
3.5. Evolutionary Relationship and Motif Analysis of CpbZIP11 Homologous Genes across Monocots and Dicots
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Xie, J.; Wang, W.; Dong, C.; Huang, L.; Wang, H.; Li, C.; Nie, S.; Xie, M. Protective effect of flavonoids from Cyclocarya paliurus leaves against carbon tetrachloride-induced acute liver injury in mice. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 119, 392–399. [Google Scholar] [CrossRef]
- Yang, Z.; Xiong, M.; Yan Jian, S.F.; Luo, S.; An, Y.; Li, K.; Yi, Y. Transcriptome Analysis of ER Stress—Related Genes and Validation of Reference Genes in Gene Expression RT-qPCR for Cyclocarya paliurus (Batal.) Iljinskaja. Int. J. Agric. Biol. 2019, 22, 1588–1598. [Google Scholar]
- Sun, T.; Ablaev, A.G.; Wang, Y.; Li, C. Cyclocarya paliurus (Batal.) Iljinskaja (Juglandaceae) from the Hunchun Formation (Eocene), Jilin Province, China. J. Integr. Plant Biol. 2005, 47, 1281–1287. [Google Scholar] [CrossRef]
- Zheng, X.; Xiao, H.; Chen, J.; Zhu, J.; Fu, Y.; Ouyang, S.; Chen, Y.; Chen, D.; Su, J.; Xue, T. Metabolome and Whole-Transcriptome Analyses Reveal the Molecular Mechanisms Underlying Hypoglycemic Nutrient Metabolites Biosynthesis in Cyclocarya paliurus Leaves During Different Harvest Stages. Front. Nutr. 2022, 9, 851569. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Peng, Y.; Zhu, X.; Li, H.; Zhang, L.; Kong, F.; Wang, J.; Yu, D. The phytochemicals and health benefits of Cyclocarya paliurus (Batalin) Iljinskaja. Front. Nutr. 2023, 10, 1158158. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tang, C.; Gao, Z.; Huang, Y.; Zhang, B.; Wei, J.; Zhao, L.; Tong, X. Potential Role of Natural Plant Medicine Cyclocarya paliurus in the Treatment of Type 2 Diabetes Mellitus. J. Diabetes Res. 2021, 2021, 1655336. [Google Scholar] [CrossRef]
- Chen, Z.; Jian, Y.; Wu, Q.; Wu, J.; Sheng, W.; Jiang, S.; Shehla, N.; Aman, S.; Wang, W. Cyclocarya paliurus (Batalin) Iljinskaja: Botany, Ethnopharmacology, phytochemistry and pharmacology. J. Ethnopharmacol. 2022, 285, 114912. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fu, X.; Shang, X.; Yang, W.; Fang, S. Natural population structure and genetic differentiation for heterodicogamous plant: Cyclocarya paliurus (Batal.) Iljinskaja (Juglandaceae). Tree Genet. Genomes 2017, 13, 80. [Google Scholar] [CrossRef]
- Fang, S.; Wang, J.; Wei, Z.; Zhu, Z. Methods to break seed dormancy in Cyclocarya paliurus (Batal.) Iljinskaja. Sci. Hortic-Amst. 2006, 110, 305–309. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, M.; Qiao, Y.; Li, R.; Alkan, N.; Chen, J.; Chen, F. Cyclocarya paliurus Reprograms the Flavonoid Biosynthesis Pathway against Colletotrichum fructicola. Front. Plant Sci. 2022, 13, 933484. [Google Scholar] [CrossRef]
- Weidemüller, P.; Kholmatov, M.; Petsalaki, E.; Zaugg, J.B. Transcription factors: Bridge between cell signaling and gene regulation. Proteomics 2021, 21, e2000034. [Google Scholar] [CrossRef]
- Hobert, O. Gene regulation by transcription factors and microRNAs. Science 2008, 319, 1785–1786. [Google Scholar] [CrossRef] [PubMed]
- Renkawitz, R. Transcription Factors and Regulation of Gene Expression; Springer: Berlin/Heidelberg, Germany, 2006; pp. 1886–1890. [Google Scholar]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; He, F.; An, Y.; Zhang, N.; Fan, S.; Tang, M.; Li, K. Genome-Wide Identification and Expression Analysis of Salt Tolerance-Associated WRKY Family Genes in Cyclocarya paliurus. Forests 2023, 14, 1771. [Google Scholar] [CrossRef]
- Lemmer, I.L.; Willemsen, N.; Hilal, N.; Bartelt, A. A guide to understanding endoplasmic reticulum stress in metabolic disorders. Mol. Metab. 2021, 47, 101169. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Tang, J.; Srivastava, R.; Bassham, D.C.; Howell, S.H. The Transcription Factor bZIP60 Links the Unfolded Protein Response to the Heat Stress Response in Maize. Plant Cell 2020, 32, 3559–3575. [Google Scholar] [CrossRef]
- Hwang, J.; Qi, L. Quality Control in the Endoplasmic Reticulum: Crosstalk between ERAD and UPR pathways. Trends Biochem. Sci. 2018, 43, 593–605. [Google Scholar] [CrossRef]
- Rellmann, Y.; Eidhof, E.; Dreier, R. Review: ER stress-induced cell death in osteoarthritic cartilage. Cell Signal 2021, 78, 109880. [Google Scholar] [CrossRef]
- Read, A.; Schröder, M. The Unfolded Protein Response: An Overview. Biology 2021, 10, 384. [Google Scholar] [CrossRef]
- Wiseman, R.L.; Mesgarzadeh, J.S.; Hendershot, L.M. Reshaping endoplasmic reticulum quality control through the unfolded protein response. Mol. Cell 2022, 82, 1477–1491. [Google Scholar] [CrossRef]
- Wan, S.; Jiang, L. Endoplasmic reticulum (ER) stress and the unfolded protein response (UPR) in plants. Protoplasma 2016, 253, 753–764. [Google Scholar] [CrossRef]
- Srivastava, R.; Deng, Y.; Howell, S.H. Stress sensing in plants by an ER stress sensor/transducer, bZIP28. Front. Plant Sci. 2014, 5, 59. [Google Scholar] [CrossRef] [PubMed]
- Iwata, Y.; Ashida, M.; Hasegawa, C.; Tabara, K.; Mishiba, K.; Koizumi, N. Activation of the Arabidopsis membrane-bound transcription factor bZIP28 is mediated by site-2 protease, but not site-1 protease. Plant J. Cell Mol. Biol. 2017, 91, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Moreno, A.A.; Mukhtar, M.S.; Blanco, F.; Boatwright, J.L.; Moreno, I.; Jordan, M.R.; Chen, Y.; Brandizzi, F.; Dong, X.; Orellana, A.; et al. IRE1/bZIP60-mediated unfolded protein response plays distinct roles in plant immunity and abiotic stress responses. PLoS ONE 2012, 7, e31944. [Google Scholar] [CrossRef] [PubMed]
- Howell, S.H. Evolution of the unfolded protein response in plants. Plant Cell Environ. 2021, 44, 2625–2635. [Google Scholar] [CrossRef]
- Nakamura, M.; Nozaki, M.; Iwata, Y.; Koizumi, N.; Sato, Y. THESEUS1 is involved in tunicamycin-induced root growth inhibition, ectopic lignin deposition, and cell wall damage-induced unfolded protein response. Plant Biotechnol. 2022, 39, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Messias Sandes, J.; Nascimento Moura, D.M.; Divina da Silva Santiago, M.; Barbosa de Lima, G.; Cabral Filho, P.E.; da Cunha Gonçalves de Albuquerque, S.; de Paiva Cavalcanti, M.; Fontes, A.; Bressan Queiroz Figueiredo, R.C. The effects of endoplasmic reticulum stressors, tunicamycin and dithiothreitol on Trypanosoma cruzi. Exp. Cell Res. 2019, 383, 111560. [Google Scholar] [CrossRef] [PubMed]
- Back, S.H.; Schröder, M.; Lee, K.; Zhang, K.; Kaufman, R.J. ER stress signaling by regulated splicing: IRE1/HAC1/XBP1. Methods 2005, 35, 395–416. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, S.; Lu, S.; Liu, J. Site-1 protease cleavage site is important for the ER stress-induced activation of membrane-associated transcription factor bZIP28 in Arabidopsis. Sci. China Life Sci. 2015, 58, 270–275. [Google Scholar] [CrossRef]
- Paridon, A.; Fox, A.; Alvero, A.B. Detection of Unfolded Protein Response by Polymerase Chain Reaction. Methods Mol. Biol. 2021, 2255, 13–20. [Google Scholar] [CrossRef]
- Nagashima, Y.; Iwata, Y.; Ashida, M.; Mishiba, K.; Koizumi, N. Exogenous salicylic acid activates two signaling arms of the unfolded protein response in Arabidopsis. Plant Cell Physiol. 2014, 55, 1772–1778. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Kaitheri Kandoth, P. Tomato bZIP60 mRNA undergoes splicing in endoplasmic reticulum stress and in response to environmental stresses. Plant Physiol. Bioch 2021, 160, 397–403. [Google Scholar] [CrossRef]
- Lu, S.; Yang, Z.; Sun, L.; Sun, L.; Song, Z.; Liu, J. Conservation of IRE1-Regulated bZIP74 mRNA Unconventional Splicing in Rice (Oryza sativa L.) Involved in ER Stress Responses. Mol. Plant 2012, 5, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Pan, X.; Yu, Y.; Wang, W.; Zhang, F.; Ge, Y.; Shen, X.; Shen, F.; Liu, X. De novo characterization of the Anthurium transcriptome and analysis of its digital gene expression under cold stress. Bmc Genom. 2013, 14, 827. [Google Scholar] [CrossRef]
- Davidson, N.M.; Oshlack, A. Corset: Enabling differential gene expression analysis for de novo assembled transcriptomes. Genome Biol. 2014, 15, 410. [Google Scholar] [CrossRef]
- Huang, Q.; Sun, M.; Yan, P. Pathway and Network Analysis of Differentially Expressed Genes in Transcriptomes. Methods Mol. Biol. 2018, 1751, 35–55. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Yang, Z.T.; Fan, S.X.; Li, R.; Huang, T.M.; An, Y.; Guo, Z.Q.; Li, F.; Yi, Y.; Li, K. The optimal reference gene validation in Cyclocarya paliurus (Batal.) Iljinskaja under environmental stresses. Agron. J. 2022, 114, 2044–2055. [Google Scholar] [CrossRef]
- Tao, Y.; Chen, L.; Jin, J.; Du, Z.; Li, J. Genome-wide identification and analysis of bZIP gene family reveal their roles during development and drought stress in Wheel Wingnut (Cyclocarya paliurus). BMC Genom. 2022, 23, 743. [Google Scholar] [CrossRef]
- Deng, Y.; Humbert, S.; Liu, J.; Srivastava, R.; Rothstein, S.J.; Howell, S.H. Heat induces the splicing by IRE1 of a mRNA encoding a transcription factor involved in the unfolded protein response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 7247–7252. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Fan, S.; Wang, J.; An, Y.; Guo, Z.; Li, K.; Liu, J. The plasma membrane-associated transcription factor NAC091 regulates unfolded protein response in Arabidopsis thaliana. Plant Sci. 2023, 334, 111777. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Shen, M.; Liu, S.; Yu, Q.; Chen, Y.; Xie, J. Ameliorative effect of Cyclocarya paliurus polysaccharides against carbon tetrachloride induced oxidative stress in liver and kidney of mice. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2020, 135, 111014. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Shang, X.; Ding, H.; Cao, Y.; Fang, S. Natural variations in flavonoids and triterpenoids of Cyclocarya paliurus leaves. J. For. Res. 2021, 32, 805–814. [Google Scholar] [CrossRef]
- Genome-Wide Identification and Expression Analysis of bZIP Gene Family in Carthamus tinctorius L.—Scientific Reports. Available online: https://www.nature.com/articles/s41598-020-72390-z (accessed on 14 September 2023).
- Yang, Z.; Sun, J.; Chen, Y.; Zhu, P.; Zhang, L.; Wu, S.; Ma, D.; Cao, Q.; Li, Z.; Xu, T. Genome-wide identification, structural and gene expression analysis of the bZIP transcription factor family in sweet potato wild relative Ipomoea trifida. BMC Genet. 2019, 20, 41. [Google Scholar] [CrossRef] [PubMed]
- LIU, J.; SRIVASTAVA, R.; HOWELL, S.H. Stress-induced expression of an activated form of AtbZIP17 provides protection from salt stress in Arabidopsis. Plant Cell Environ. 2008, 31, 1735–1743. [Google Scholar] [CrossRef]
- Yang, O.; Popova, O.V.; Süthoff, U.; Lüking, I.; Dietz, K.; Golldack, D. The Arabidopsis basic leucine zipper transcription factor AtbZIP24 regulates complex transcriptional networks involved in abiotic stress resistance. Gene 2009, 436, 45–55. [Google Scholar] [CrossRef]
- Baoxiang, W.; Yan, L.; Yifeng, W.; Jingfang, L.; Zhiguang, S.; Ming, C.; Yungao, X.; Bo, X.; Bo, Y.; Jian, L.; et al. OsbZIP72 Is Involved in Transcriptional Gene-Regulation Pathway of Abscisic Acid Signal Transduction by Activating Rice High-Affinity Potassium Transporter OsHKT1;1. Rice Sci. 2021, 28, 257–267. [Google Scholar] [CrossRef]
- Liu, C.; Wu, Y.; Wang, X. bZIP transcription factor OsbZIP52/RISBZ5: A potential negative regulator of cold and drought stress response in rice. Planta 2012, 235, 1157–1169. [Google Scholar] [CrossRef]
- Chakraborty, R.; Baek, J.H.; Bae, E.Y.; Kim, W.; Lee, S.Y.; Kim, M.G. Comparison and contrast of plant, yeast, and mammalian ER stress and UPR. Appl. Biol. Chem. 2016, 59, 337–347. [Google Scholar] [CrossRef]
- Iwata, Y.; Koizumi, N. An Arabidopsis transcription factor, AtbZIP60, regulates the endoplasmic reticulum stress response in a manner unique to plants. Proc. Natl. Acad. Sci. USA 2005, 102, 5280–5285. [Google Scholar] [CrossRef]
- Nagashima, Y.; Mishiba, K.; Suzuki, E.; Shimada, Y.; Iwata, Y.; Koizumi, N. Arabidopsis IRE1 catalyses unconventional splicing of bZIP60 mRNA to produce the active transcription factor. Sci. Rep. 2011, 1, 29. [Google Scholar] [CrossRef] [PubMed]
- Rubio, C.; Pincus, D.; Korennykh, A.; Schuck, S.; El-Samad, H.; Walter, P. Homeostatic adaptation to endoplasmic reticulum stress depends on Ire1 kinase activity. J. Cell Biol. 2011, 193, 171–184. [Google Scholar] [CrossRef]
- Siwecka, N.; Rozpędek-Kamińska, W.; Wawrzynkiewicz, A.; Pytel, D.; Diehl, J.A.; Majsterek, I. The Structure, Activation and Signaling of IRE1 and Its Role in Determining Cell Fate. Biomedicines 2021, 9, 156. [Google Scholar] [CrossRef] [PubMed]
- Almanza, A.; Mnich, K.; Blomme, A.; Robinson, C.M.; Rodriguez-Blanco, G.; Kierszniowska, S.; McGrath, E.P.; Le Gallo, M.; Pilalis, E.; Swinnen, J.V.; et al. Regulated IRE1α-dependent decay (RIDD)-mediated reprograming of lipid metabolism in cancer. Nat. Commun. 2022, 13, 2493. [Google Scholar] [CrossRef]
- Park, S.; Kang, T.; So, J. Roles of XBP1s in Transcriptional Regulation of Target Genes. Biomedicines 2021, 9, 791. [Google Scholar] [CrossRef]
- Maurel, M.; Chevet, E.; Tavernier, J.; Gerlo, S. Getting RIDD of RNA: IRE1 in cell fate regulation. Trends Biochem. Sci. 2014, 39, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Bukau, B.; Weissman, J.; Horwich, A. Molecular Chaperones and Protein Quality Control. Cell 2006, 125, 443–451. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, Y.; He, F.; Ye, Q.; Fan, S.; Zeng, Y.; Tang, M.; Yang, Z.; Li, K. Identification of CpbZIP11 in Cyclocarya paliurus Involved in Environmental Stress Responses. Forests 2023, 14, 2104. https://doi.org/10.3390/f14102104
An Y, He F, Ye Q, Fan S, Zeng Y, Tang M, Yang Z, Li K. Identification of CpbZIP11 in Cyclocarya paliurus Involved in Environmental Stress Responses. Forests. 2023; 14(10):2104. https://doi.org/10.3390/f14102104
Chicago/Turabian StyleAn, Yin, Fayin He, Qian Ye, Sixian Fan, Yue Zeng, Ming Tang, Zhengting Yang, and Kun Li. 2023. "Identification of CpbZIP11 in Cyclocarya paliurus Involved in Environmental Stress Responses" Forests 14, no. 10: 2104. https://doi.org/10.3390/f14102104
APA StyleAn, Y., He, F., Ye, Q., Fan, S., Zeng, Y., Tang, M., Yang, Z., & Li, K. (2023). Identification of CpbZIP11 in Cyclocarya paliurus Involved in Environmental Stress Responses. Forests, 14(10), 2104. https://doi.org/10.3390/f14102104