
Machine Learning: Crown Diameter Predictive Modeling for Open-Grown Trees in the Cerrado Biome, Brazil

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
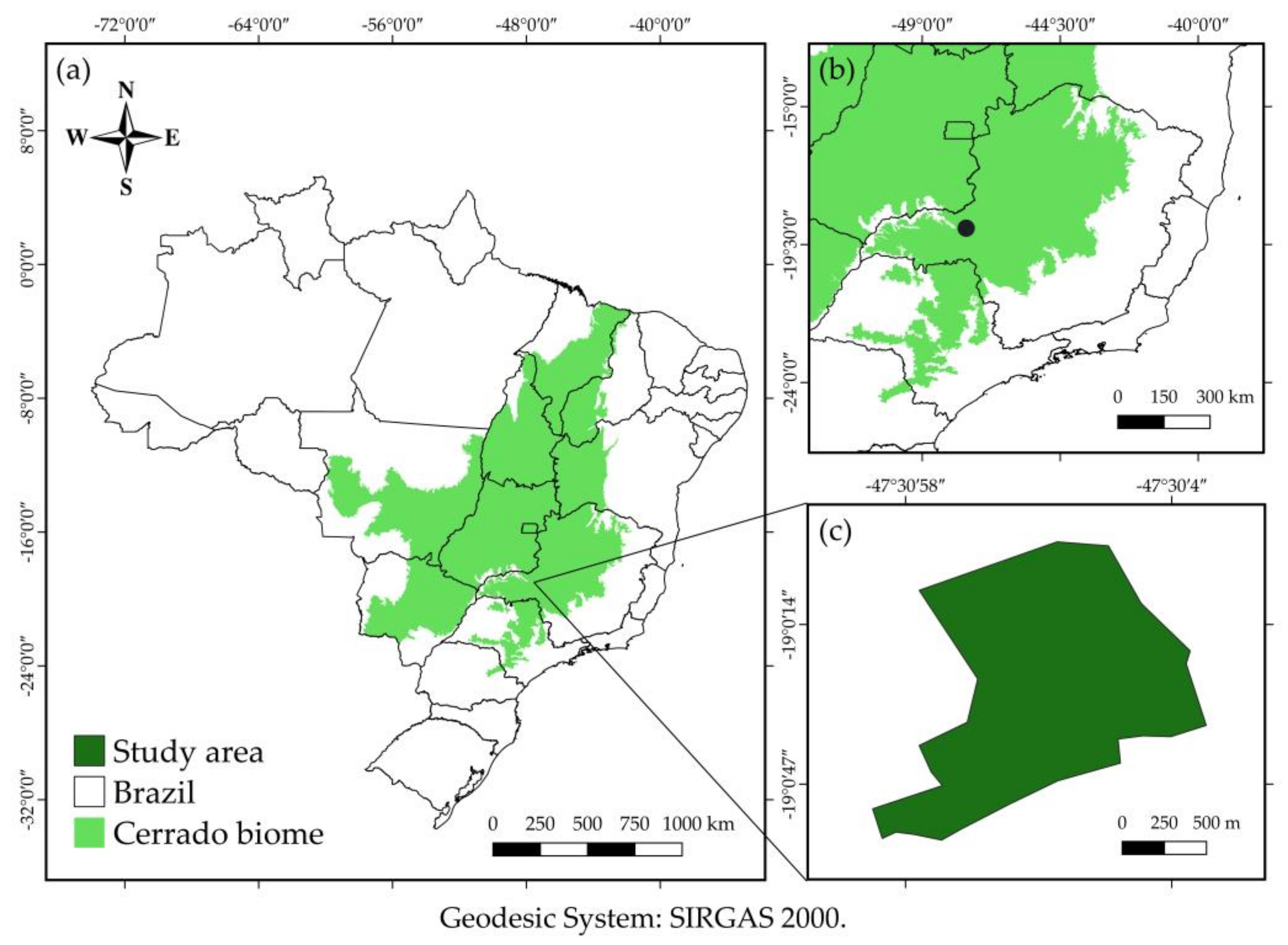
2.1. Study Area Description
2.2. Data Collection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Description |
|---|---|
| Diameter measured 0.3 m (D03) | D03 was measured with a diameter tape (cm), 0.30 m above the soil. |
| Diameter measured 0.7 m (D07) | D07 was measured with a diameter tape (cm), 0.70 m above the soil. |
| Equivalent diameter (Deq) | Deq was measured with a diameter tape (cm) 1.30 m above the soil. In case of forked stems, their equivalent diameter (Deq) was estimated by Equation (1): (1), see, e.g., Figure 2a, in which Deq is their equivalent diameter in cm and Di, their diameter measured 1.3 m above the soil in cm. |
| Crown base height (CBH) | Measured by a Vertex hypsometer, accurate to tenths of meters, from the soil up to the insertion point of the crown (Figure 2b). |
| Height (H) | Measured by a Vertex hypsometer, accurate to tenths of meters (see, e.g., Figure 2b). |
| Crown length (CL) | Calculated by the difference between total and crown base height. |
| Crown diameter (CD) |
Crown radii were measured according to their cardinal positions. Crown diameter was estimated by Equation (2): |
| in which CD refers to crown diameter in m; cr, to crown radii (in which N stands for north; NO, to northeast; E, to east; SO, to southeast; S, to south; SW, to southwest; W, to west; and NO, to northeast) in m. | |
| Crown geometric shapes (CGS) | Tree crown geometric shapes (CGS) were classified into circular (a), elliptic (b), columnar (c), umbelliform (d), and pyramidal I (Figure 3). |
| Stoniness (ST) | The presence of rocky outcrops on the site was classified as (0) no visual outcrop; (1) outcrop lower than 33.3%; (2) outcrop from 33.3% to 66.6%; and (3) outcrop higher than 66.6%. |
| Tree position on the relief (PR) | Classified into (1) lower hillside, (2) average hillside, (3) upper hillside, and (4) plateau. |
| Vitality (VT) | The number of leaves alive and their distribution in the crown were determined visually and via health classes as (1) high vitality, (2) average vitality, and (3) low vitality. |
| Branch arrangement (BA) | Assessed as (1) homogeneously well-spread branches over all four quadrants, (2) distributed into three quadrants, and (3) branches distributed into one or two quadrants. |
2.3. Crown Radius
2.4. Selection of the Most Important Predictor Variables
2.5. Modeling of the Crown Diameter
2.6. Selecting the Best Models
3. Results
3.1. Characteristics of Measured Trees
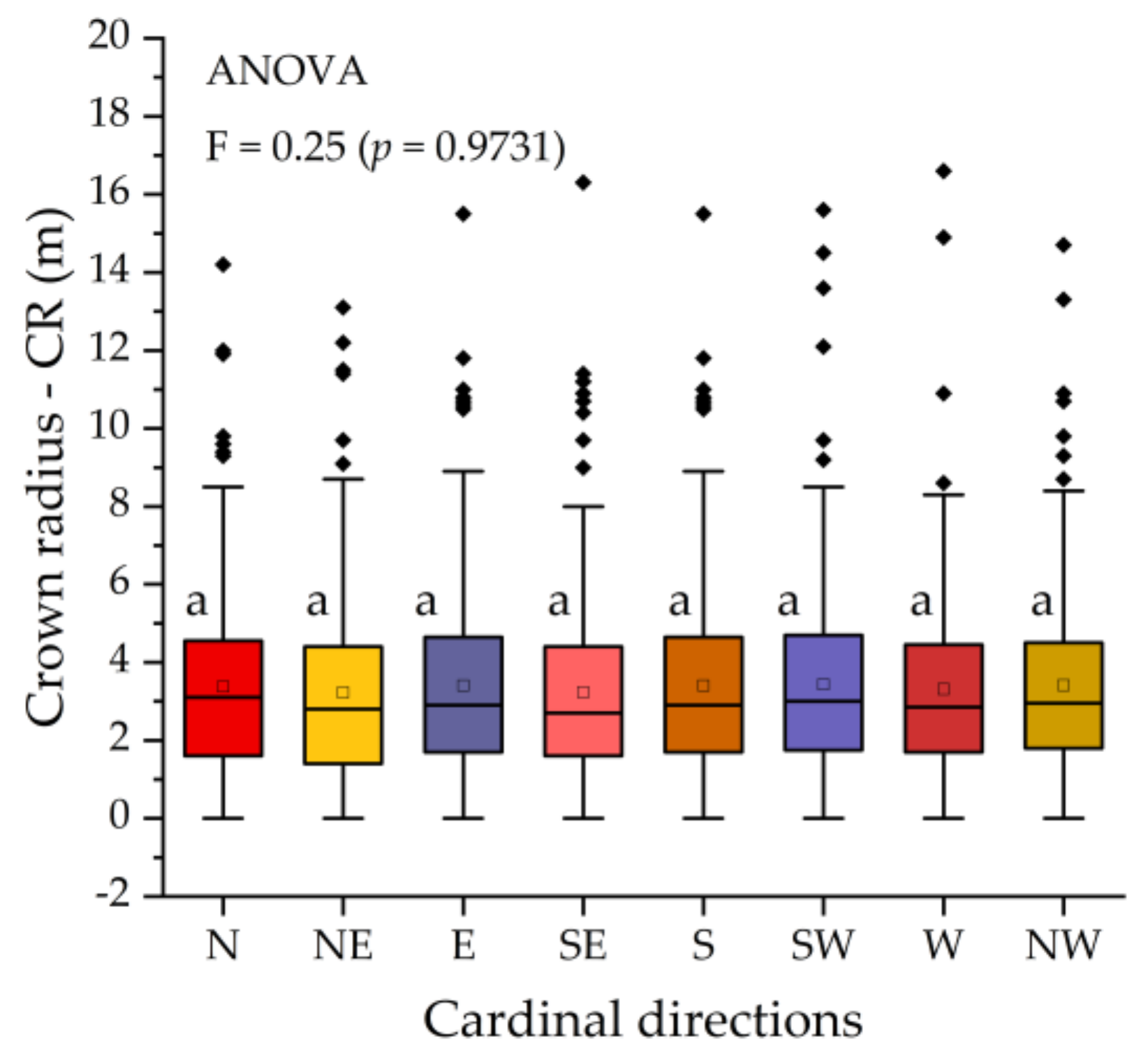
3.2. Crown Radius Variations between Cardinal Directions
3.3. Selection of Important Variables
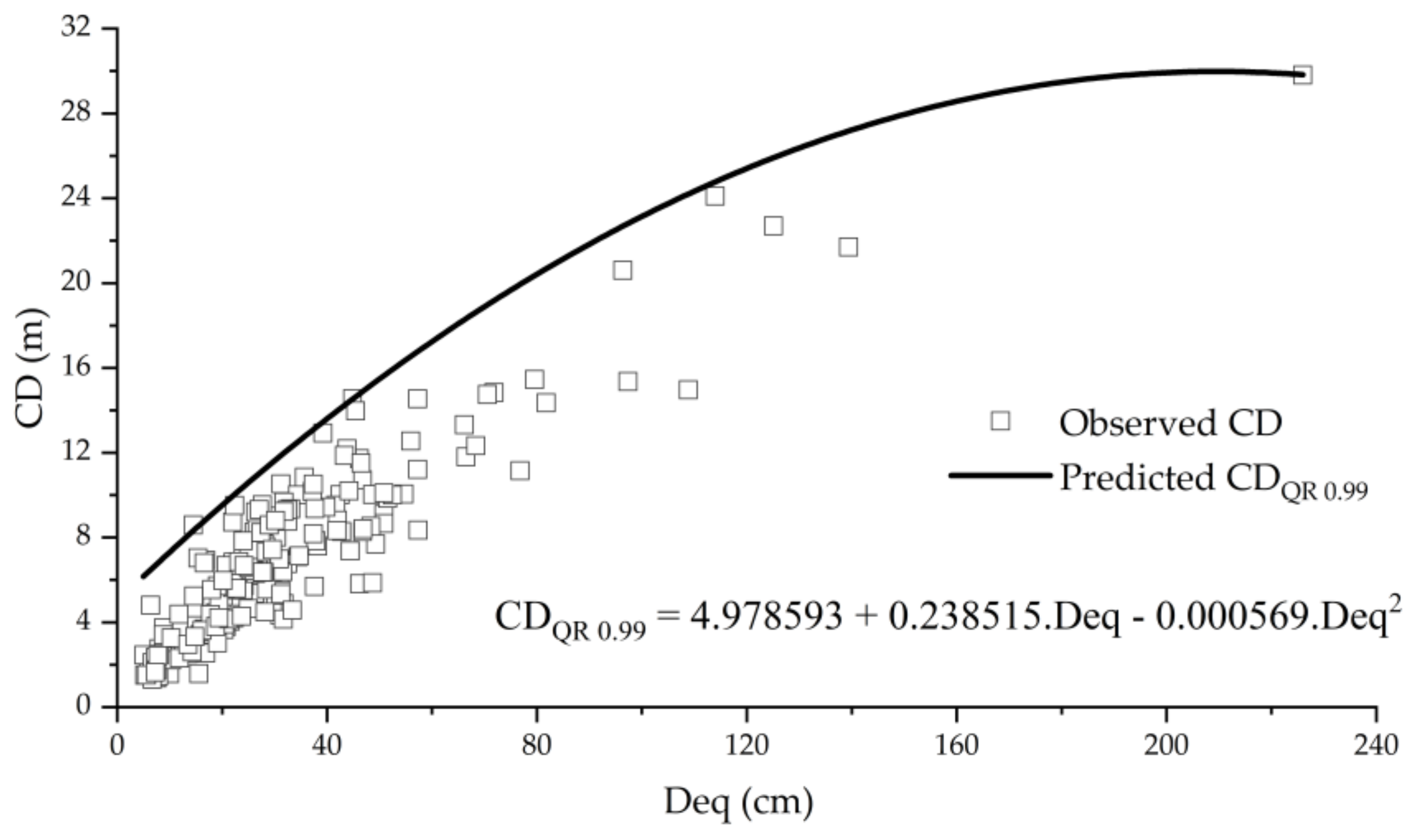
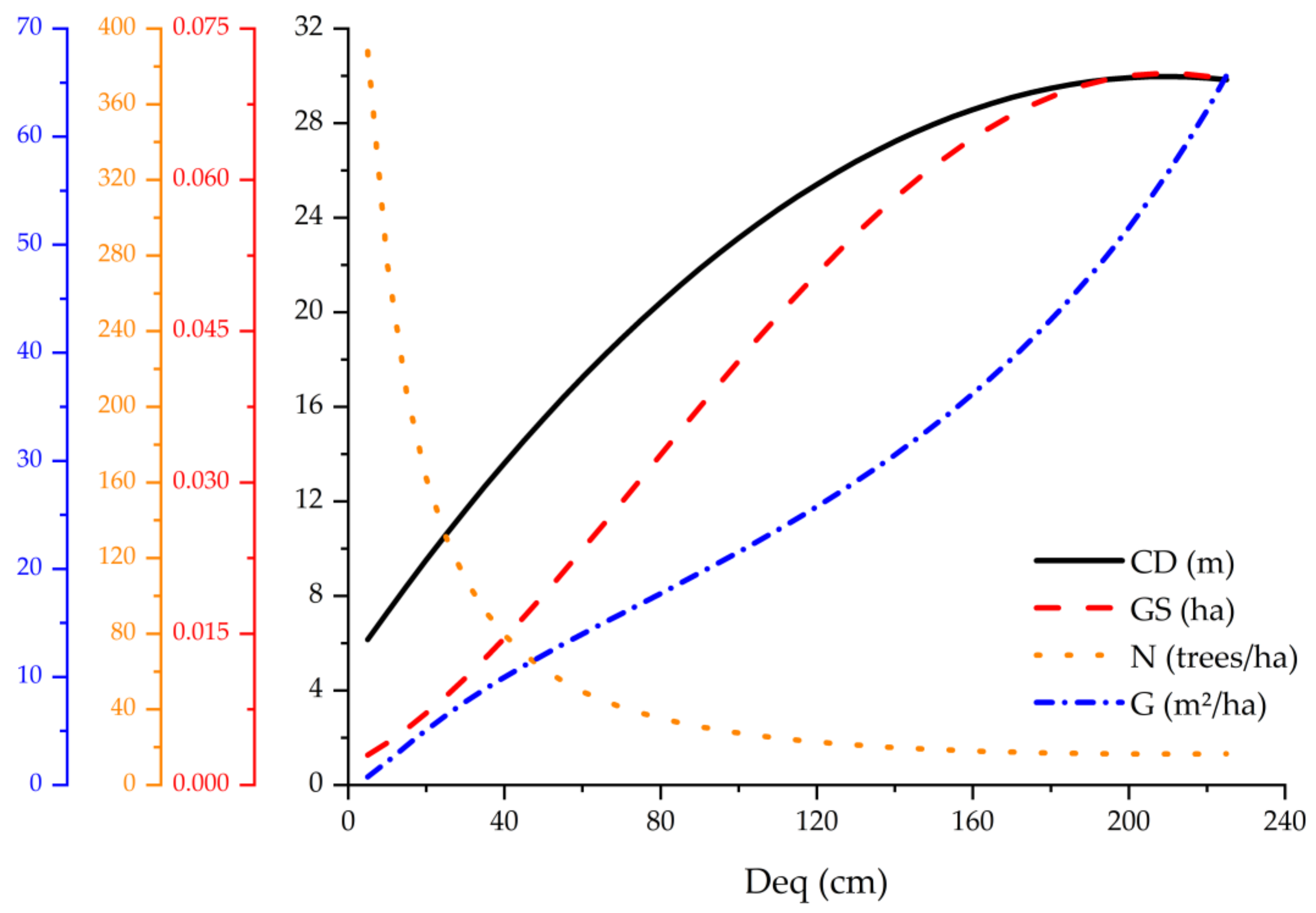
3.4. Crown Diameter Potential
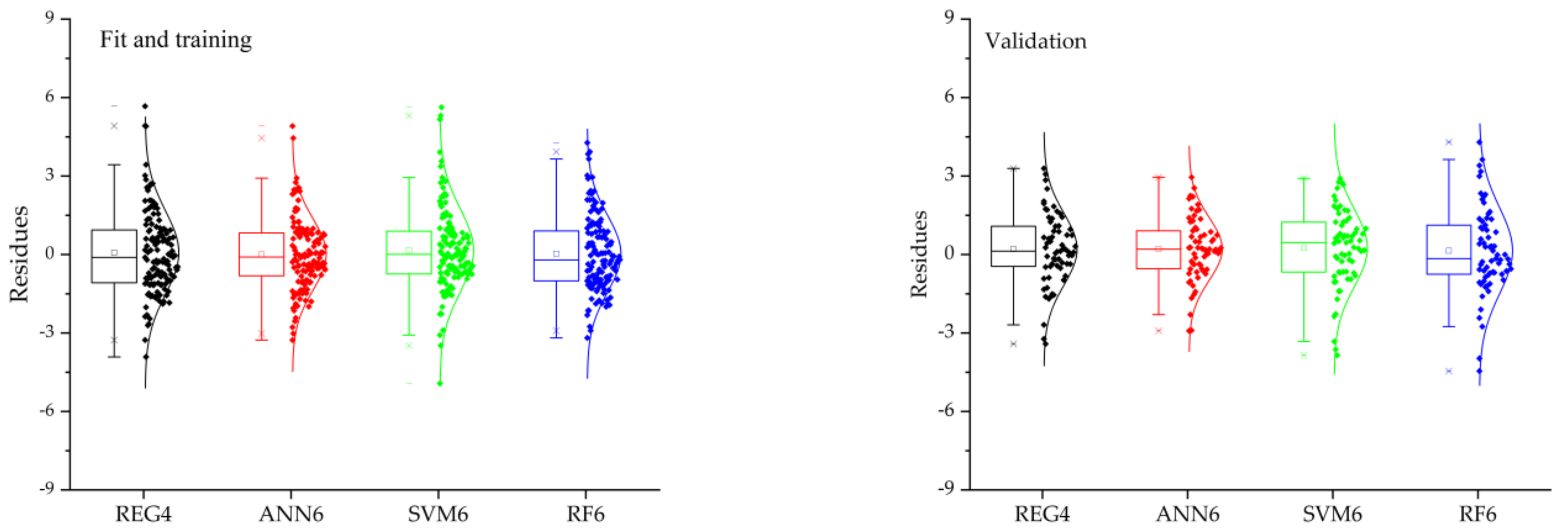
3.5. Evaluation of the Regression Analysis and Machine Learning Techniques to Predict Crown Diameter
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Oliveira, V.A.; de Mello, C.R.; Beskow, S.; Viola, M.R.; Srinivasan, R. Modeling the effects of climate change on hydrology and sediment load in a headwater basin in the Brazilian Cerrado biome. Ecol. Eng. 2019, 133, 20–31. [Google Scholar] [CrossRef]
- Barbieri, A.R.; Szabó, M.P.J.; Costa, F.B.; Martins, T.F.; Soares, H.S.; Pascoli, G.; Torga, K.; Saraiva, D.G.; Ramos, V.N.; Osava, C.; et al. Species richness and seasonal dynamics of ticks with notes on rickettsial infection in a Natural Park of the Cerrado biome in Brazil. Ticks Tick-Borne Dis. 2019, 10, 442–453. [Google Scholar] [CrossRef] [PubMed]
- De Miranda, S.D.C.; Bustamante, M.; Palace, M.; Hagen, S.; Keller, M.; Ferreira, L.G. Regional Variations in Biomass Distribution in Brazilian Savanna Woodland. Biotropica 2014, 46, 125–138. [Google Scholar] [CrossRef]
- Morandi, P.S.; Marimon, B.S.; Marimon-Junior, B.H.; Ratter, J.A.; Feldpausch, T.R.; Colli, G.R.; Munhoz, C.B.R.; Júnior, M.C.D.S.; Lima, E.D.S.; Haidar, R.F.; et al. Tree diversity and above-ground biomass in the South America Cerrado biome and their conservation implications. Biodivers. Conserv. 2018, 29, 1519–1536. [Google Scholar] [CrossRef]
- Loram-Lourenço, L.; Farnese, F.D.S.; de Sousa, L.F.; Alves, R.D.F.B.; De Andrade, M.C.P.; Almeida, S.E.D.S.; Moura, L.M.D.F.; Costa, A.C.; Silva, F.G.; Galmés, J.; et al. A Structure Shaped by Fire, but Also Water: Ecological Consequences of the Variability in Bark Properties Across 31 Species from the Brazilian Cerrado. Front. Plant Sci. 2020, 10, 1718. [Google Scholar] [CrossRef]
- Roque, M.P.B.; Neto, J.A.F.; de Faria, A.L.L.; Ferreira, F.M.; Teixeira, T.H.; Coelho, L.L. Effectiveness of Arguments Used in the Creation of Protected Areas of Sustainable Use in Brazil: A Case Study from the Atlantic Forest and Cerrado. Sustainability 2019, 11, 1700. [Google Scholar] [CrossRef]
- Nunes, A.R.; Rodrigues, A.L.M.; de Queiróz, D.B.; Vieira, I.G.P.; Neto, J.F.C.; Junior, J.T.C.; Tintino, S.R.; de Morais, S.M.; Coutinho, H.D.M. Photoprotective potential of medicinal plants from Cerrado biome (Brazil) in relation to phenolic content and antioxidant activity. J. Photochem. Photobiol. B Biol. 2018, 189, 119–123. [Google Scholar] [CrossRef]
- Bechtold, W.A. Crown-Diameter Prediction Models for 87 Species of Stand-Grown Trees in the Eastern United States. South. J. Appl. For. 2003, 27, 269–278. [Google Scholar] [CrossRef]
- Hasenauer, H. Dimensional relationships of open-grown trees in Austria. For. Ecol. Manag. 1997, 96, 197–206. [Google Scholar] [CrossRef]
- Foli, E.; Alder, D.; Miller, H.; Swaine, M. Modelling growing space requirements for some tropical forest tree species. For. Ecol. Manag. 2003, 173, 79–88. [Google Scholar] [CrossRef]
- Raptis, D.; Kazana, V.; Kazaklis, A.; Stamatiou, C. A Crown Width-Diameter Model for Natural Even-Aged Black Pine Forest Management. Forests 2018, 9, 610. [Google Scholar] [CrossRef]
- Márquez-Linares, M.A.; Alvarez-Zagoya, R. Construcción de una guía de densidad para Pinus cooperi var. ornelasi con base en la clase de copa en Durango, México. Madera Y Bosques 1995, 1, 23–36. [Google Scholar] [CrossRef]
- Pretzsch, H. Trees grow modulated by the ecological memory of their past growth. Consequences for monitoring, modelling, and silvicultural treatment. For. Ecol. Manag. 2021, 487, 118982. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, K.; Schumacher, J.; Gougeon, F.A. Scale-dependent competition at the stand level assessed from crown areas. For. Ecol. Manag. 2008, 255, 2478–2485. [Google Scholar] [CrossRef]
- Sharma, R.P.; Bílek, L.; Vacek, Z.; Vacek, S. Modelling crown width–diameter relationship for Scots pine in the central Europe. Trees 2017, 31, 1875–1889. [Google Scholar] [CrossRef]
- Fu, L.; Sharma, R.P.; Hao, K.; Tang, S. A generalized interregional nonlinear mixed-effects crown width model for Prince Rupprecht larch in northern China. For. Ecol. Manag. 2017, 389, 364–373. [Google Scholar] [CrossRef]
- Goodman, R.C.; Phillips, O.; Baker, T.R. The importance of crown dimensions to improve tropical tree biomass estimates. Ecol. Appl. 2014, 24, 680–698. [Google Scholar] [CrossRef]
- Lau, A.; Calders, K.; Bartholomeus, H.; Martius, C.; Raumonen, P.; Herold, M.; Vicari, M.; Sukhdeo, H.; Singh, J.; Goodman, R.C. Tree Biomass Equations from Terrestrial LiDAR: A Case Study in Guyana. Forests 2019, 10, 527. [Google Scholar] [CrossRef]
- Özçelik, R.; Diamantopoulou, M.; Brooks, J. The use of tree crown variables in over-bark diameter and volume prediction models. Iforest Biogeosciences For. 2014, 7, 132–139. [Google Scholar] [CrossRef]
- Gill, S.J.; Biging, G.S.; Murphy, E.C. Modeling conifer tree crown radius and estimating canopy cover. For. Ecol. Manag. 2000, 126, 405–416. [Google Scholar] [CrossRef]
- Pretzsch, H.; Biber, P.; Uhl, E.; Dahlhausen, J.; Rötzer, T.; Caldentey, J.; Koike, T.; van Con, T.; Chavanne, A.; Seifert, T.; et al. Crown size and growing space requirement of common tree species in urban centres, parks, and forests. Urban For. Urban Green. 2015, 14, 466–479. [Google Scholar] [CrossRef]
- Costa, E.A.; Finger, C.A.G.; Fleig, F.D. Modelagem do espaço de crescimento para araucária. Ciência Florest. 2018, 28, 725–734. [Google Scholar] [CrossRef]
- Pretzsch, H.; Ahmed, S.; Jacobs, M.; Schmied, G.; Hilmers, T. Linking crown structure with tree ring pattern: Methodological considerations and proof of concept. Trees 2022, 36, 1349–1367. [Google Scholar] [CrossRef]
- Srinivasan, S.; Popescu, S.; Eriksson, M.; Sheridan, R.; Ku, N.-W.J.R.S. Terrestrial Laser Scanning as an Effective Tool to Retrieve Tree Level Height, Crown Width, and Stem Diameter. Remote Sens. 2015, 7, 1877–1896. [Google Scholar] [CrossRef]
- Zhen, Z.; Quackenbush, L.J.; Zhang, L. Impact of Tree-Oriented Growth Order in Marker-Controlled Region Growing for Individual Tree Crown Delineation Using Airborne Laser Scanner (ALS) Data. Remote Sens. 2013, 6, 555–579. [Google Scholar] [CrossRef]
- Lim, Y.S.; La, P.H.; Park, J.S.; Lee, M.H.; Pyeon, M.W.; Kim, J.-I. Calculation of Tree Height and Canopy Crown from Drone Images Using Segmentation. Korean J. Geomat. 2015, 33, 605–614. [Google Scholar] [CrossRef]
- Lockhart, B.R.; Weih, R.C., Jr.; Smith, K. Crown Radius and Diameter at Breast Height Relationships for Six Bottomland Hardwood Species. J. Ark. Acad. Sci. 2005, 59, 110–115. [Google Scholar]
- Quadri, A. Development and Evaluation of Linear and Non-Linear Models for Diameter at Breast Height and Crown Diameter of Triplochiton scleroxylon (K. Schum) Plantations in Oyo State, Nigeria. J. Agric. Vet. Sci. 2019, 12, 47–52. [Google Scholar] [CrossRef]
- Goodfellow, I.; Bengio, Y.; Courville, A. Deep Learning; MIT Press: Cambridge, MA, USA, 2016; Volume 1, pp. 1–800. ISBN 978-0-262-03561-3. [Google Scholar]
- Costa, E.A.; Liesenberg, V.; Finger, C.A.G.; Hess, A.F.; Schons, C.T. Understanding bark thickness variations for Araucaria angustifolia in southern Brazil. J. For. Res. 2020, 32, 1077–1087. [Google Scholar] [CrossRef]
- Zhang, X.-D. Machine Learning. In A Matrix Algebra Approach to Artificial Intelligence; Springer: Singapore, 2020; pp. 223–440. ISBN 02-62-01243-X. [Google Scholar]
- El Naqa, I.; Murphy, M.J. What is machine learning? In Machine Learning in Radiation Oncology; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Silva, A.K.V.; Vinicius, M.; Borges, V.; Batista, T.S.; Antonio, C.; Elis, D.; Furuya, G.; Osco, L.P.; Pereira, L.; Teodoro, R.; et al. Predicting Eucalyptus Diameter at Breast Height and Total Height with UAV-Based Spectral Indices and Machine Learning. Forests 2021, 12, 582. [Google Scholar] [CrossRef]
- Dos Reis, A.A.; Carvalho, M.C.; de Mello, J.M.; Gomide, L.R.; Filho, A.C.F.; Junior, F.W.A. Spatial prediction of basal area and volume in Eucalyptus stands using Landsat TM data: An assessment of prediction methods. N. Z. J. For. Sci. 2018, 48, 1. [Google Scholar] [CrossRef]
- Bayat, M.; Bettinger, P.; Heidari, S.; Khalyani, A.H.; Jourgholami, M.; Hamidi, S.K. Estimation of Tree Heights in an Uneven-Aged, Mixed Forest in Northern Iran Using Artificial Intelligence and Empirical Models. Forests 2020, 11, 324. [Google Scholar] [CrossRef]
- Júnior, I.D.S.T.; Torres, C.M.M.E.; Leite, H.G.; de Castro, N.L.M.; Soares, C.P.B.; Castro, R.V.O.; Farias, A.A. Machine learning: Modeling increment in diameter of individual trees on Atlantic Forest fragments. Ecol. Indic. 2020, 117, 106685. [Google Scholar] [CrossRef]
- Zhang, Z. Artificial Neural Network. In Multivariate Time Series Analysis in Climate and Environmental Research; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; pp. 1–35. ISBN 9783319673400. [Google Scholar]
- Castro, R.V.O.; Soares, C.P.B.; Leite, H.G.; De Souza, A.L.; Nogueira, G.S.; Martins, F.B. Individual Growth Model for Eucalyptus Stands in Brazil Using Artificial Neural Network. ISRN For. 2013, 2013, 196832. [Google Scholar] [CrossRef]
- Vieira, G.C.; de Mendonça, A.R.; da Silva, G.F.; Zanetti, S.S.; da Silva, M.M.; dos Santos, A.R. Prognoses of diameter and height of trees of eucalyptus using artificial intelligence. Sci. Total Environ. 2018, 619–620, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, S.J.S.S.; Torres, C.M.M.E.; Jacovine, L.A.G.; Leite, H.G.; Gelcer, E.M.; Neves, K.M.; Schettini, B.L.S.; Villanova, P.H.; da Silva, L.F.; Reis, L.P.; et al. Artificial neural networks: Modeling tree survival and mortality in the Atlantic Forest biome in Brazil. Sci. Total Environ. 2018, 645, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Sanquetta, C.R.; Piva, L.R.; Wojciechowski, J.; Corte, A.P.; Schikowski, A.B. Volume estimation of Cryptomeria japonica logs in southern Brazil using artificial intelligence models. South. For. A J. For. Sci. 2017, 80, 29–36. [Google Scholar] [CrossRef]
- Sakici, O.E.; Ozdemir, G. Stem taper estimations with artificial neural networks for mixed oriental beech and kazdaği fir stands in Karabük Region, Turkey. Cerne 2018, 24, 439–451. [Google Scholar] [CrossRef]
- Karatzoglou, A.; Meyer, D.; Hornik, K. Support Vector Machines in R. J. Stat. Softw. 2006, 15, 1–28. [Google Scholar] [CrossRef]
- Lafetá, B.O.; Trindade, R.N.R.; Aguiar, V.F.; Penido, T.M.A.; Vieira, D.D.S. Crown morphometric indexes of eucalypt estimated by logistic regression and support vector machines. Floresta 2020, 50, 1633–1642. [Google Scholar] [CrossRef]
- Cordeiro, M.A.; Pereira, N.N.D.J.; Binoti, D.H.B.; Binoti, M.L.M.D.S.; Leite, H.G. Estimativa do volume de Acacia mangium utilizando técnicas de redes neurais artificiais e máquinas vetor de suporte. Pesqui. Florest. Bras. 2015, 35, 255–261. [Google Scholar] [CrossRef][Green Version]
- Binoti, D.H.B.; Binoti, M.L.M.D.S.; Leite, H.G.; Andrade, A.V.; Nogueira, G.S.; Romarco, M.L.; Pitangui, C.G. Support vector machine to estimate volume of eucalypt trees. Rev. Árvore 2016, 40, 689–693. [Google Scholar] [CrossRef][Green Version]
- Montano, R.A.N.R.; Sanquetta, C.R.; Wojciechowski, J.; Mattar, E.; Corte, A.P.D.; Todt, E. Artificial Intelligence Models to Estimate Biomass of Tropical Forest Trees. Polibits 2017, 56, 29–37. [Google Scholar]
- Ali, J.; Khan, R.; Ahmad, N.; Maqsood, I. Random Forests and Decision Trees. Int. J. Comput. Sci. Issues 2012, 9, 272–278. [Google Scholar]
- Fayad, I.; Baghdadi, N.; Bailly, J.-S.; Barbier, N.; Gond, V.; El Hajj, M.; Fabre, F.; Bourgine, B. Canopy Height Estimation in French Guiana with LiDAR ICESat/GLAS Data Using Principal Component Analysis and Random Forest Regressions. Remote Sens. 2014, 6, 11883–11914. [Google Scholar] [CrossRef]
- Srinet, R.; Nandy, S.; Patel, N. Estimating leaf area index and light extinction coefficient using Random Forest regression algorithm in a tropical moist deciduous forest, India. Ecol. Inform. 2019, 52, 94–102. [Google Scholar] [CrossRef]
- Silva, C.A.; Klauberg, C.; Hudak, A.T.; Vierling, L.A.; Jaafar, W.S.W.M.; Mohan, M.; Garcia, M.; Ferraz, A.; Cardil, A.; Saatchi, S. Predicting Stem Total and Assortment Volumes in an Industrial Pinus taeda L. Forest Plantation Using Airborne Laser Scanning Data and Random Forest. Forests 2017, 8, 254. [Google Scholar] [CrossRef]
- Jahani, A.; Saffariha, M. Tree failure prediction model (TFPM): Machine learning techniques comparison in failure hazard assessment of Platanus orientalis in urban forestry. Nat. Hazards 2021, 110, 881–898. [Google Scholar] [CrossRef]
- Jahani, A.; Saffariha, M. Modeling of trees failure under windstorm in harvested Hyrcanian forests using machine learning techniques. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Moraes, G.J.L.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Strobl, C.; Boulesteix, A.-L.; Kneib, T.; Augustin, T.; Zeileis, A. Conditional variable importance for random forests. BMC Bioinform. 2008, 9, 307. [Google Scholar] [CrossRef] [PubMed]
- Koenker, R.; Chernozhukov, V.; He, X.; Peng, L. Handbook of Quantile Regression; Koenker, R., Chernozhukov, V., He, X., Peng, L., Eds.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2017; ISBN 9781315120256. [Google Scholar]
- Draper, N.R.; Smith, H. Applied Regression Analysis, 3rd ed.; Wiley Series in Probability and Statistics; Wiley: New York, NY, USA, 1998; ISBN 978-0-471-17082-2. [Google Scholar]
- Fritsch, S.; Guenther, F.; Wright, M.N. R Package ‘Neuralnet’. Available online: https://cran.r-project.org/web/packages/neuralnet/neuralnet.pdf (accessed on 1 October 2021).
- Awad, M.; Khanna, R. Efficient Learning Machines: Theories, Concepts, and Applications for Engineers and System Designers; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Meyer, D.; Dimitriadou, E.H.K.; Weingessel, A.; Leisch, F.; Chang, C.-C.; Lin, C.-C. R Package ‘E1071’. Available online: https://cran.r-project.org/web/packages/e1071/index.html (accessed on 1 October 2021).
- Cutler, A.; Cutler, D.R.; Stevens, J.R. Ensemble Machine Learning. Ensemble Machine Learning; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. R Package “RandomForest”. Available online: https://cran.r-project.org/web/packages/randomForest/index.html (accessed on 1 October 2021).
- Hiruma-Lima, C.; Santos, L.; Kushima, H.; Pellizzon, C.; Silveira, G.; Vasconcelos, P.; Vilegas, W.; Brito, A.S. Qualea grandiflora, a Brazilian “Cerrado” medicinal plant presents an important antiulcer activity. J. Ethnopharmacol. 2006, 104, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Traesel, G.K.; de Araújo, F.H.S.; Castro, L.H.A.; de Lima, F.F.; Menegati, S.E.L.T.; Justi, P.N.; Kassuya, C.A.L.; Cardoso, C.A.L.; Argandoña, E.J.S.; Oesterreich, S.A. Safety Assessment of Oil from Pequi (Caryocar Brasiliense Camb.): Evaluation of the Potential Genotoxic and Clastogenic Effects. J. Med. Food 2017, 20, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Zardo, R.N.; Henriques, R.P.B. Growth and fruit production of the tree Caryocar brasiliense in the Cerrado of central Brazil. Agrofor. Syst. 2011, 82, 15–23. [Google Scholar] [CrossRef]
- Siqueira, E.M.D.A.; Rosa, F.R.; Fustinoni, A.M.; De Sant’Ana, L.P.; Arruda, S.F. Brazilian Savanna Fruits Contain Higher Bioactive Compounds Content and Higher Antioxidant Activity Relative to the Conventional Red Delicious Apple. PLoS ONE 2013, 8, e72826. [Google Scholar] [CrossRef] [PubMed]
- Junior, I.E.S.; Filho, V.C.; Zacchino, S.A.; Lima, J.C.D.S.; Martins, D.T.D.O. Antimicrobial screening of some medicinal plants from Mato Grosso Cerrado. Rev. Bras. Farm. 2009, 19, 242–248. [Google Scholar] [CrossRef]
- Colli, G.R.; Vieira, C.R.; Dianese, J.C. Biodiversity and conservation of the Cerrado: Recent advances and old challenges. Biodivers. Conserv. 2020, 29, 1465–1475. [Google Scholar] [CrossRef]
- Dos Santos, G.L.; Pereira, M.G.; Delgado, R.C.; Magistrali, I.C.; da Silva, C.G.; de Oliveira, C.M.M.; Larangeira, J.P.B.; da Silva, T.P. Degradation of the Brazilian Cerrado: Interactions with human disturbance and environmental variables. For. Ecol. Manag. 2020, 482, 118875. [Google Scholar] [CrossRef]
- Silva, J.P.M.; da Silva, M.L.M.; de Mendonça, A.R.; da Silva, G.F.; Junior, A.A.d.B.; da Silva, E.F.; Aguiar, M.O.; Santos, J.S.; Rodrigues, N.M.M. Prognosis of forest production using machine learning techniques. Inf. Process. Agric. 2021. [Google Scholar] [CrossRef]
- Lopes, I.L.; Araújo, L.A.; Miranda, E.N.; Bastos, T.A.; Gomide, L.R.; Castro, G.P. A comparative approach of methods to estimate machine productivity in wood cutting. Int. J. For. Eng. 2021, 33, 43–55. [Google Scholar] [CrossRef]
- Davies, O.; Pommerening, A. The contribution of structural indices to the modelling of Sitka spruce (Picea sitchensis) and birch (Betula spp.) crowns. For. Ecol. Manag. 2008, 256, 68–77. [Google Scholar] [CrossRef]
- Korpela, I.; Dahlin, B.; Schäfer, H.; Bruun, E.; Haapaniemi, F.; Honkasalo, J.; Ilvesniemi, S.; Kuutti, V.; Linkosalmi, M.; Mustonen, J.; et al. Single-Tree Forest Inventory Using Lidar and Aerial Images for 3D Treetop Positioning, Species Recognition, Height and Crown Width Estimation. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2007, 36, 227–233. [Google Scholar]
- Gspaltl, M.; Sterba, H.; O’Hara, K.L. The relationship between available area efficiency and area exploitation index in an even-aged coast redwood (Sequoia sempervirens) stand. For. Int. J. For. Res. 2012, 85, 567–577. [Google Scholar] [CrossRef]
- Ramírez-Iglesias, E.; Hernández-Hernandez, R.M.; Iglesias, J.R. Nutrient dynamics of corn fed to livestock in conservation-based agroecosystems in neotropical savannas. Rev. De Cienc. Agrovet. 2021, 20, 086–092. [Google Scholar] [CrossRef]
- Ansolin, R.D.; Santos, K.S.M.; Fernandes, A.P.D.; Schinato, F. Environmental Valuation of Permanent Preservation Areas of the Passaúna River Watershed, Paraná State. Rev. De Cienc. Agrovet. 2018, 17, 118–127. [Google Scholar] [CrossRef]
- Yang, K.; Tu, J.; Chen, T. Homoscedasticity: An overlooked critical assumption for linear regression. Gen. Psychiatry 2019, 32, e100148. [Google Scholar] [CrossRef] [PubMed]
- Graham, M.H. Confronting multicollinearity in ecological multiple regression. Ecology 2003, 84, 2809–2815. [Google Scholar] [CrossRef]
- Coombes, A.; Martin, J.; Slater, D. Defining the allometry of stem and crown diameter of urban trees. Urban For. Urban Green. 2019, 44, 126421. [Google Scholar] [CrossRef]
- Panagiotidis, D.; Abdollahnejad, A.; Surový, P.; Chiteculo, V. Determining tree height and crown diameter from high-resolution UAV imagery. Int. J. Remote Sens. 2017, 38, 2392–2410. [Google Scholar] [CrossRef]
- De Freitas, E.C.S.; de Paiva, H.N.; Neves, J.C.L.; Marcatti, G.E.; Leite, H.G. Modeling of eucalyptus productivity with artificial neural networks. Ind. Crop. Prod. 2020, 146, 112149. [Google Scholar] [CrossRef]
- Cosenza, D.N.; Soares, A.; De Alcântara, A.E.M.; Da Silva, A.A.L.; Rode, R.; Soares, V.P.; Leite, H.G. Site classification for eucalypt stands using artificial neural network based on environmental and management features. Cerne 2017, 23, 310–320. [Google Scholar] [CrossRef]
- Nwankpa, C.; Ijomah, W.; Gachagan, A.; Marshall, S. Activation Functions: Comparison of Trends in Practice and Research for Deep Learning. arXiv 2018, arXiv:1811.03378. [Google Scholar]
- Sharma, S.; Sharma, S.; Anidhya, A. Activation Function in Neural Networks. Int. J. Eng. Appl. Sci. Technol. 2020, 4, 310–316. [Google Scholar] [CrossRef]
- Mercioni, M.A.; Holban, S. The most used activation functions: Classic versus current. In Proceedings of the 15th International Conference on Development and Application Systems, DAS, Suceava, Romania, 21–21 May 2020; pp. 141–145. [Google Scholar] [CrossRef]
- Reis, P.C.M.; de Souza, A.L.; Reis, L.P.; Carvalho, A.M.M.L.; Mazzei, L.; Rêgo, L.J.S.; Leite, H.G. Artificial Neural Networks to Estimate the Physical-Mechanical Properties of Amazon Second Cutting Cycle Wood. Maderas Cienc. Y Tecnol. 2018, 20, 343–352. [Google Scholar] [CrossRef]
- Sheela, K.G.; Deepa, S.N. Selection of Number of Hidden Neurons in Neural Networks in Renewable Energy Systems. J. Sci. Ind. Res. 2013, 73, 686–688. [Google Scholar]
- Dantas, D.; Pinto, L.O.R.; Terra, M.D.C.N.S.; Calegario, N.; De Oliveira, M.L.R. Reduction of sampling intensity in forest inventories to estimate the total height of eucalyptus trees. Bosque 2020, 41, 353–364. [Google Scholar] [CrossRef]
- Júnior, I.D.S.T.; da Rocha, J.E.C.; Ebling, A.A.; Chaves, A.D.S.; Zanuncio, J.C.; Farias, A.A.; Leite, H.G. Artificial Neural Networks and Linear Regression Reduce Sample Intensity to Predict the Commercial Volume of Eucalyptus Clones. Forests 2019, 10, 268. [Google Scholar] [CrossRef]
- Monnet, J.-M.; Chanussot, J.; Berger, F. Support Vector Regression for the Estimation of Forest Stand Parameters Using Airborne Laser Scanning. IEEE Geosci. Remote Sens. Lett. 2011, 8, 580–584. [Google Scholar] [CrossRef]
- Feizizadeh, B.; Roodposhti, M.S.; Blaschke, T.; Aryal, J. Comparing GIS-based support vector machine kernel functions for landslide susceptibility mapping. Arab. J. Geosci. 2017, 10, 122. [Google Scholar] [CrossRef]
- Nanda, M.A.; Seminar, K.B.; Nandika, D.; Maddu, A. A Comparison Study of Kernel Functions in the Support Vector Machine and Its Application for Termite Detection. Information 2018, 9, 5. [Google Scholar] [CrossRef]
- Hong, H.; Pradhan, B.; Bui, D.T.; Xu, C.; Youssef, A.M.; Chen, W. Comparison of four kernel functions used in support vector machines for landslide susceptibility mapping: A case study at Suichuan area (China). Geomat. Nat. Hazards Risk 2017, 8, 544–569. [Google Scholar] [CrossRef]
- Dantas, D.; Calegario, N.; Acerbi, F.W.; Carvalho, S.D.P.C.; Júnior, F.W.A.; Melo, E.D.A. Multilevel nonlinear mixed-effects model and machine learning for predicting the volume of Eucalyptus spp. trees. CERNE 2020, 26, 48–57. [Google Scholar] [CrossRef]
- De Souza, G.S.A.; Soares, V.P.; Leite, H.G.; Gleriani, J.M.; Amaral, C.H.D.; Ferraz, A.S.; Silveira, M.V.D.F.; dos Santos, J.F.C.; Velloso, S.G.S.; Domingues, G.F.; et al. Multi-sensor prediction of Eucalyptus stand volume: A support vector approach. ISPRS J. Photogramm. Remote Sens. 2019, 156, 135–146. [Google Scholar] [CrossRef]
- Ahmed, O.S.; Franklin, S.E.; Wulder, M.A.; White, J.C. Characterizing stand-level forest canopy cover and height using Landsat time series, samples of airborne LiDAR, and the Random Forest algorithm. ISPRS J. Photogramm. Remote Sens. 2015, 101, 89–101. [Google Scholar] [CrossRef]
- Muhairwe, C.K. Taper equations for Eucalyptus pilularis and Eucalyptus grandis for the north coast in New South Wales, Australia. For. Ecol. Manag. 1999, 113, 251–269. [Google Scholar] [CrossRef]
- Oshiro, T.M.; Perez, P.S.; Baranauskas, J.A. How many trees in a random forest? In Lecture Notes in Computer Science; Including Subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics; Springer: Berlin/Heidelberg, Germany, 2012; pp. 154–168. [Google Scholar] [CrossRef]
- Probst, P.; Boulesteix, A.L. To Tune or Not to Tune the Number of Trees in Random Forest. J. Mach. Learn. Res. 2018, 18, 1–8. [Google Scholar]
- Liu, K.; Shen, X.; Cao, L.; Wang, G.; Cao, F. Estimating forest structural attributes using UAV-LiDAR data in Ginkgo plantations. ISPRS J. Photogramm. Remote Sens. 2018, 146, 465–482. [Google Scholar] [CrossRef]
- Osco, L.P.; Ramos, A.P.M.; Pereira, D.R.; Moriya, A.S.; Imai, N.N.; Matsubara, E.T.; Estrabis, N.; de Souza, M.; Junior, J.M.; Gonçalves, W.N.; et al. Predicting Canopy Nitrogen Content in Citrus-Trees Using Random Forest Algorithm Associated to Spectral Vegetation Indices from UAV-Imagery. Remote Sens. 2019, 11, 2925. [Google Scholar] [CrossRef]
- Lee, J.; Im, J.; Kim, K.-M.; Quackenbush, L.J. Machine Learning Approaches for Estimating Forest Stand Height Using Plot-Based Observations and Airborne LiDAR Data. Forests 2018, 9, 268. [Google Scholar] [CrossRef]
- Wu, C.; Shen, H.; Shen, A.; Deng, J.; Gan, M.; Zhu, J.; Xu, H.; Wang, K. Comparison of machine-learning methods for above-ground biomass estimation based on Landsat imagery. J. Appl. Remote Sens. 2016, 10, 35010. [Google Scholar] [CrossRef]
- Sanquetta, C.R.; Corte, A.P.D.; Jacon, A.D. Crown Area and Trunk Diameter Relationship for Tree Species at a Mixed-Araucaria Natural Forest in the Mid-Southern Parana State, Brazil. Floresta 2011, 41, 63–72. [Google Scholar] [CrossRef]
- Pereira, P.R.M.; Bolfe, E.L.; Rodrigues, T.C.S.; Sano, E.E. Dynamics of agricultural expansion in areas of the Brazilian Savanna between 2000 and 2019. Int. Arch. Photogramm. Remote Sens. Spatial Inf. Sci. 2020, XLIII-B3, 1607–1614. [Google Scholar] [CrossRef]
- Haykin, S. Redes Neurais–Princípios e Prática, 2nd ed.; Engel, P.M., Ed.; Bookman Editora: Porto Alegre, Brazil, 2001; Volume 7659, ISBN 85-7307-718-2. [Google Scholar]
- Heaton, J. Introduction to the Math of Neural Networks, 1st ed.; WordsRU.com, Ed.; Heaton Research, Inc.: Chesterfield, MO, USA, 2012; ISBN 978-1475190878. [Google Scholar]









| Variables | Units | Minimum | Mean | Maximum | Standard Deviation |
|---|---|---|---|---|---|
| D03 | cm | 5.9 | 37.4 | 226.0 | 29.7 |
| D07 | cm | 5.6 | 33.8 | 226.0 | 26.8 |
| Deq | cm | 5.1 | 32.1 | 226.0 | 25.3 |
| CBH | m | 1.0 | 3.3 | 10.4 | 1.5 |
| H | m | 2.3 | 9.5 | 27.5 | 5.0 |
| CL | m | 0.8 | 6.2 | 23.8 | 4.0 |
| CD | m | 1.3 | 7.1 | 29.8 | 4.2 |
| CGS | - | 1.0 | 2.6 | 5.0 | 0.9 |
| ST | - | 2.0 | 2.9 | 3.0 | 0.3 |
| PR | - | 1.0 | 1.2 | 3.0 | 0.4 |
| VT | - | 1.0 | 1.3 | 3.0 | 0.5 |
| BA | - | 1.0 | 1.5 | 5.0 | 0.6 |
| Spearman’s Correlation | Random Forest | |||||
|---|---|---|---|---|---|---|
| CD | Units | ρ | p-Value | Rank | Importance | Rank |
| D03 | cm | 0.860 | <0.0001 | 2 | 0.860 | 2 |
| D07 | cm | 0.854 | <0.0001 | 3 | 0.707 | 4 |
| Deq | cm | 0.867 | <0.0001 | 1 | 1.000 | 1 |
| CBH | m | 0.477 | <0.0001 | 6 | 0.415 | 5 |
| H | m | 0.787 | <0.0001 | 5 | 0.407 | 6 |
| CL | m | 0.792 | <0.0001 | 4 | 0.768 | 3 |
| CGS | - | −0.344 | <0.0001 | 7 | 0.219 | 7 |
| ST | - | −0.017 | 0.8482 | 10 | 0.038 | 11 |
| PR | - | 0.002 | 0.9835 | 11 | 0.056 | 10 |
| VT | - | −0.179 | 0.0379 | 8 | 0.095 | 9 |
| BA | - | −0.153 | 0.0760 | 9 | 0.123 | 8 |
| Id. | Input | β0 | β1 | β2 | β3 | β4 | Fit | Validation | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R2 | RMSE | MAE | MAPE | R2 | RMSE | MAE | MAPE | |||||||
| 1 | Deq | 2.3151 | 0.1463 | 0.81 | 1.89 | 1.48 | 28.22 | 0.81 | 1.71 | 1.36 | 26.07 | |||
| 2 | Deq; CL | 1.6476 | 0.1053 | 0.3254 | 0.84 | 1.74 | 1.40 | 25.60 | 0.83 | 1.59 | 1.23 | 23.30 | ||
| 3 | Deq; CL; CGS | 3.6930 | 0.0923 | 0.3816 | −0.7655 | 0.87 | 1.60 | 1.26 | 22.26 | 0.87 | 1.40 | 1.09 | 19.54 | |
| 4 | Deq; Deq2; CL; CGS | 3.0437 | 0.1271 | −0.0002 | 0.3174 | −0.6895 | 0.87 | 1.59 | 1.24 | 20.98 | 0.88 | 1.38 | 1.05 | 18.09 |
| Technique | Id. | Input | MLP | Activation Function | Trees | Training | Validation | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hidden | Output | R2 | RMSE | MAE | MAPE | R2 | RMSE | MAE | MAPE | |||||
| ANN | 1 | Deq | 1-3-1 | Logistic | Linear | 0.85 | 1.73 | 1.34 | 23.18 | 0.84 | 1.56 | 1.21 | 20.27 | |
| SVM | 1 | 0.80 | 1.95 | 1.45 | 23.76 | 0.81 | 1.72 | 1.33 | 22.11 | |||||
| RF | 1 | 90 | 0.86 | 1.62 | 1.26 | 21.59 | 0.84 | 1.56 | 1.21 | 20.17 | ||||
| ANN | 2 | Deq; D03 | 2-6-1 | Logistic | Linear | 0.85 | 1.70 | 1.34 | 23.42 | 0.83 | 1.58 | 1.25 | 21.24 | |
| SVM | 2 | 0.81 | 1.89 | 1.45 | 24.15 | 0.78 | 1.82 | 1.44 | 24.37 | |||||
| RF | 2 | 75 | 0.88 | 1.54 | 1.23 | 20.96 | 0.82 | 1.64 | 1.30 | 22.14 | ||||
| ANN | 3 | Deq; D03; CL | 3-9-1 | Logistic | Linear | 0.87 | 1.57 | 1.22 | 20.94 | 0.85 | 1.49 | 1.09 | 17.67 | |
| SVM | 3 | 0.84 | 1.76 | 1.33 | 22.13 | 0.82 | 1.63 | 1.23 | 21.15 | |||||
| RF | 3 | 105 | 0.88 | 1.50 | 1.18 | 20.18 | 0.82 | 1.65 | 1.27 | 21.58 | ||||
| ANN | 4 | Deq; D03; CL; D07 | 4-12-1 | Logistic | Linear | 0.87 | 1.59 | 1.21 | 20.32 | 0.86 | 1.48 | 1.07 | 17.58 | |
| SVM | 4 | 0.84 | 1.76 | 1.33 | 21.98 | 0.82 | 1.64 | 1.23 | 21.07 | |||||
| RF | 4 | 30 | 0.89 | 1.47 | 1.16 | 20.23 | 0.81 | 1.69 | 1.32 | 22.07 | ||||
| ANN | 5 | Deq; D03; CL; D07; CBH | 5-15-1 | Logistic | Linear | 0.88 | 1.51 | 1.19 | 19.97 | 0.87 | 1.41 | 0.96 | 16.57 | |
| SVM | 5 | 0.84 | 1.76 | 1.31 | 21.74 | 0.81 | 1.68 | 1.25 | 21.10 | |||||
| RF | 5 | 105 | 0.89 | 1.45 | 1.15 | 19.63 | 0.82 | 1.67 | 1.26 | 21.25 | ||||
| ANN | 6 | Deq; D03; CL; D07; CBH; CGS | 6-18-1 | Logistic | Linear | 0.90 | 1.37 | 1.04 | 18.78 | 0.90 | 1.21 | 0.93 | 16.25 | |
| SVM | 6 | 0.87 | 1.60 | 1.15 | 19.24 | 0.85 | 1.50 | 1.18 | 19.84 | |||||
| RF | 6 | 465 | 0.89 | 1.46 | 1.16 | 20.02 | 0.82 | 1.65 | 1.25 | 21.27 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bueno, G.F.; Costa, E.A.; Finger, C.A.G.; Liesenberg, V.; Bispo, P.d.C. Machine Learning: Crown Diameter Predictive Modeling for Open-Grown Trees in the Cerrado Biome, Brazil. Forests 2022, 13, 1295. https://doi.org/10.3390/f13081295
Bueno GF, Costa EA, Finger CAG, Liesenberg V, Bispo PdC. Machine Learning: Crown Diameter Predictive Modeling for Open-Grown Trees in the Cerrado Biome, Brazil. Forests. 2022; 13(8):1295. https://doi.org/10.3390/f13081295
Chicago/Turabian StyleBueno, Gabriel Fernandes, Emanuel Arnoni Costa, César Augusto Guimarães Finger, Veraldo Liesenberg, and Polyanna da Conceição Bispo. 2022. "Machine Learning: Crown Diameter Predictive Modeling for Open-Grown Trees in the Cerrado Biome, Brazil" Forests 13, no. 8: 1295. https://doi.org/10.3390/f13081295
APA StyleBueno, G. F., Costa, E. A., Finger, C. A. G., Liesenberg, V., & Bispo, P. d. C. (2022). Machine Learning: Crown Diameter Predictive Modeling for Open-Grown Trees in the Cerrado Biome, Brazil. Forests, 13(8), 1295. https://doi.org/10.3390/f13081295