
Soil Moisture Control of NO Turnover and N2O Release in Nitrogen-Saturated Subtropical Forest Soils


Abstract
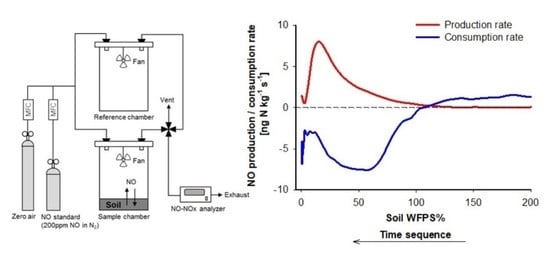
1. Introduction
2. Material and Methods
2.1. Site Characteristics
2.2. Soil Sampling, Pre-Treatment and Experimental Dry-Out
2.3. Soil Chemical and Physical Characteristics
3. Results
3.1. Soil Physical and Chemical Characteristics
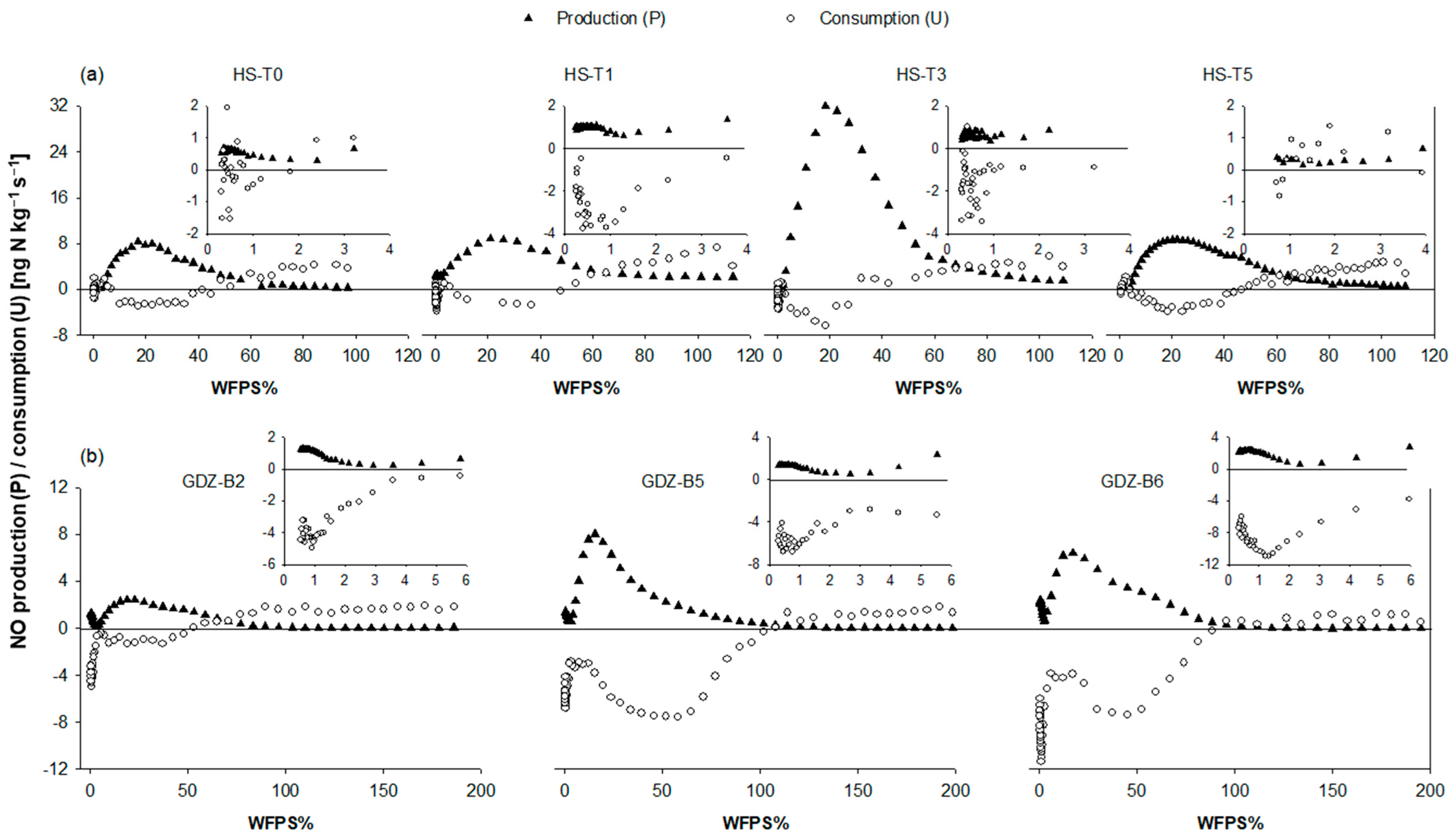
3.2. Dry-Out and Gas Phase Kinetics of NO
3.3. NO Compensation Mixing Ratios
3.4. N2O and CO2 Response to Dry-Out
3.5. Mineral Nitrogen (NH4+ and NO3−) during Dry-Out
3.6. NO/N2O Partitioning
4. Discussion
4.1. Response of NO Production to Dry-Out
4.2. Response of NO Consumption to Dry Out
4.3. N2O Response to Dry-Out
4.4. NO/N2O Partitioning
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crutzen, P.J. Role of NO and NO2 in the Chemistry of the Troposphere and Stratosphere. Annu. Rev. Earth Planet. Sci. 1979, 7, 443–472. [Google Scholar] [CrossRef]
- Larssen, T.; Lydersen, E.; Tang, D.G.; He, Y.; Gao, J.X.; Liu, H.Y.; Duan, L.; Seip, H.M.; Vogt, R.D.; Mulder, J.; et al. Acid rain in China. Environ. Sci. Technol. 2006, 40, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Volk, C.M.; Elkins, J.W.; Fahey, D.W.; Dutton, G.S.; Gilligan, J.M.; Loewenstein, M.; Podolske, J.R.; Chan, K.R.; Gunson, M.R. Evaluation of source gas lifetimes from stratospheric observations. J. Geophys. Res.-Atmos. 1997, 102, 25543–25564. [Google Scholar] [CrossRef]
- Ravishankara, A.R.; Daniel, J.S.; Portmann, R.W. Nitrous Oxide (N2O): The Dominant Ozone-Depleting Substance Emitted in the 21st Century. Science 2009, 326, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Denman, K.L.; Brasseur, G.; Chidthaisong, A.; Ciais, P.; Cox, P.M.; Dickinson, R.E.; Hauglustaine, D.; Heinze, C.; Holland, E.; Jacob, D.; et al. Couplings Between Changes in the Climate System and Biogeochemistry. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Steinkamp, J.; Lawrence, M.G. Improvement and evaluation of simulated global biogenic soil NO emissions in an AC-GCM. Atmos. Chem. Phys. 2011, 11, 6063–6082. [Google Scholar] [CrossRef]
- Butterbach-Bahl, K.; Gasche, R.; Huber, C.; Kreutzer, K.; Papen, H. Impact of N-input by wet deposition on N-trace gas fluxes and CH4-oxidation in spruce forest ecosystems of the temperate zone in Europe. Atmos. Environ. 1998, 32, 559–564. [Google Scholar] [CrossRef]
- Pilegaard, K.; Skiba, U.; Ambus, P.; Beier, C.; Bruggemann, N.; Butterbach-Bahl, K.; Dick, J.; Dorsey, J.; Duyzer, J.; Gallagher, M.; et al. Factors controlling regional differences in forest soil emission of nitrogen oxides (NO and N2O). Biogeosciences 2006, 3, 651–661. [Google Scholar] [CrossRef]
- Li, D.; Wang, X.; Mo, J.; Sheng, G.; Fu, J. Soil nitric oxide emissions from two subtropical humid forests in south China. J. Geophys. Res. 2007, 112, 8680. [Google Scholar] [CrossRef]
- Huang, Y.; Li, D. Soil nitric oxide emissions from terrestrial ecosystems in China: A synthesis of modeling and measurements. Sci. Rep. 2014, 4, 7406. [Google Scholar] [CrossRef]
- Ke, P.; Kang, R.; Avery, L.K.; Zhang, J.; Yu, Q.; Xie, D.; Duan, L. Temporal variations of soil NO and NO2 fluxes in two typical subtropical forests receiving contrasting rates of N deposition. Environ. Pollut. 2022, 295, 118696. [Google Scholar] [CrossRef]
- Zhu, J.; Mulder, J.; Wu, L.P.; Meng, X.X.; Wang, Y.H.; Dorsch, P. Spatial and temporal variability of N2O emissions in a subtropical forest catchment in China. Biogeosciences 2013, 10, 1309–1321. [Google Scholar] [CrossRef]
- Zhu, J.; Mulder, J.; Solheimslid, S.O.; Dorsch, P. Functional traits of denitrification in a subtropical forest catchment in China with high atmogenic N deposition. Soil Biol. Biochem. 2013, 57, 577–586. [Google Scholar] [CrossRef]
- Zhu, J.; Mulder, J.; Bakken, L.; Dorsch, P. The importance of denitrification for N2O emissions from an N-saturated forest in SW China: Results from in situ N-15 labeling experiments. Biogeochemistry 2013, 116, 103–117. [Google Scholar] [CrossRef]
- Yu, L.F.; Zhu, J.; Mulder, J.; Dorsch, P. Multiyear dual nitrate isotope signatures suggest that N-saturated subtropical forested catchments can act as robust N sinks. Glob. Chang. Biol. 2016, 22, 3662–3674. [Google Scholar] [CrossRef]
- Yu, L.; Mulder, J.; Zhu, J.; Zhang, X.; Wang, Z.; Dorsch, P. Denitrification as a major regional nitrogen sink in subtropical forest catchments: Evidence from multi-site dual nitrate isotopes. Glob. Chang. Biol. 2019, 25, 1765–1778. [Google Scholar] [CrossRef]
- Firestone, M.K.; Davidson, E.A. Microbiological Basis of NO and N2O Production and Consumption in Soil. In Exchange of Trace Gases between Terrestrial Ecosystems and the Atmosphere; Andreae, M.O., Schimel, D.S., Eds.; John Wiley & Sons Ltd.: New York, NY, USA, 1989; Volume 47, pp. 7–21. [Google Scholar]
- Skopp, J.; Jawson, M.D.; Doran, J.W. Steady-State Aerobic Microbial Activity as a Function of Soil-Water Content. Soil Sci. Soc. Am. J. 1990, 54, 1619–1625. [Google Scholar] [CrossRef]
- Oswald, R.; Behrendt, T.; Ermel, M.; Wu, D.; Su, H.; Cheng, Y.; Breuninger, C.; Moravek, A.; Mougin, E.; Delon, C.; et al. HONO Emissions from Soil Bacteria as a Major Source of Atmospheric Reactive Nitrogen. Science 2013, 341, 1233–1235. [Google Scholar] [CrossRef]
- VanCleemput, O.; Samater, A.H. Nitrite in soils: Accumulation and role in the formation of gaseous N compounds. Fertil. Res. 1996, 45, 81–89. [Google Scholar] [CrossRef]
- Wei, J.; Amelung, W.; Lehndorff, E.; Schloter, M.; Vereecken, H.; Brüggemann, N. N2O and NOx emissions by reactions of nitrite with soil organic matter of a Norway spruce forest. Biogeochemistry 2017, 132, 325–342. [Google Scholar] [CrossRef]
- Kang, R.H.; Mulder, J.; Duan, L.; Dorsch, P. Spatial and temporal variability of soil nitric oxide emissions in N-saturated subtropical forest. Biogeochemistry 2017, 134, 337–351. [Google Scholar] [CrossRef]
- Chen, X.Y.; Mulder, J. Atmospheric deposition of nitrogen at five subtropical forested sites in South China. Sci. Total Environ. 2007, 378, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.M.; Kang, R.H.; Mulder, J.; Zhang, T.; Duan, L. Nitrogen saturation, soil acidification, and ecological effects in a subtropical pine forest on acid soil in southwest China. J. Geophys. Res. Biogeosci. 2015, 120, 2457–2472. [Google Scholar] [CrossRef]
- Chen, X.Y.; Mulder, J. Indicators for nitrogen status and leaching in subtropical forest ecosystems, South China. Biogeochemistry 2007, 82, 165–180. [Google Scholar] [CrossRef]
- Sorbotten, L.E.; Stolte, J.; Wang, Y.H.; Mulder, J. Hydrological Responses and Flow Pathways in an Acrisol on a Forested Hillslope with a Monsoonal Subtropical Climate. Pedosphere 2017, 27, 1037–1048. [Google Scholar] [CrossRef]
- Bargsten, A.; Falge, E.; Pritsch, K.; Huwe, B.; Meixner, F.X. Laboratory measurements of nitric oxide release from forest soil with a thick organic layer under different understory types. Biogeosciences 2010, 7, 1425–1441. [Google Scholar] [CrossRef]
- Behrendt, T.; Veres, P.R.; Ashuri, F.; Song, G.; Flanz, M.; Mamtimin, B.; Bruse, M.; Williams, J.; Meixner, F.X. Characterisation of NO production and consumption: New insights by an improved laboratory dynamic chamber technique. Biogeosciences 2014, 11, 5463–5492. [Google Scholar] [CrossRef]
- Remde, A.; Slemr, F.; Conrad, R. Microbial-Production and Uptake of Nitric-Oxide in Soil. FEMS Microbiol. Ecol. 1989, 62, 221–230. [Google Scholar] [CrossRef]
- Molstad, L.; Dorsch, P.; Bakken, L.R. Robotized incubation system for monitoring gases (O2, NO, N2O N2) in denitrifying cultures. J. Microbiol. Methods 2007, 71, 202–211. [Google Scholar] [CrossRef]
- Homyak, P.M.; Sickman, J.O. Influence of soil moisture on the seasonality of nitric oxide emissions from chaparral soils, Sierra Nevada, California, USA. J. Arid Environ. 2014, 103, 46–52. [Google Scholar] [CrossRef][Green Version]
- Werner, C.; Reiser, K.; Dannenmann, M.; Hutley, L.B.; Jacobeit, J.; Butterbach-Bahl, K. N2O, NO, N2 and CO2 emissions from tropical savanna and grassland of northern Australia: An incubation experiment with intact soil cores. Biogeosciences 2014, 11, 6047–6065. [Google Scholar] [CrossRef]
- Homyak, P.M.; Blankinship, J.C.; Marchus, K.; Lucero, D.M.; Sickman, J.O.; Schimel, J.P. Aridity and plant uptake interact to make dryland soils hotspots for nitric oxide (NO) emissions. Proc. Natl. Acad. Sci. USA 2016, 113, E2608–E2616. [Google Scholar] [CrossRef] [PubMed]
- Homyak, P.M.; Kamiyama, M.; Sickman, J.O.; Schimel, J.P. Acidity and organic matter promote abiotic nitric oxide production in drying soils. Glob. Chang. Biol. 2017, 23, 1735–1747. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, T.; Braker, G.; Song, G.; Pommerenke, B.; Dörsch, P. Nitric oxide emission response to soil moisture is linked to transcriptional activity of functional microbial groups. Soil Biol. Biochem. 2017, 115, 337–345. [Google Scholar] [CrossRef]
- Song, L.; Drewer, J.; Zhu, B.; Zhou, M.; Cowan, N.; Levy, P.; Skiba, U. The impact of atmospheric N deposition and N fertilizer type on soil nitric oxide and nitrous oxide fluxes from agricultural and forest Eutric Regosols. Biol. Fertil. Soils 2020, 56, 1077–1090. [Google Scholar] [CrossRef]
- Krichels, A.H.; Homyak, P.M.; Aronson, E.L.; Sickman, J.O.; Botthoff, J.; Shulman, H.; Piper, S.; Andrews, H.M.; Jenerette, G.D. Rapid nitrate reduction produces pulsed NO and N2O emissions following wetting of dryland soils. Biogeochemistry 2022, 158, 233–250. [Google Scholar] [CrossRef]
- van Dijk, S.M.; Meixner, F.X. Production and consumption of NO in forest and pasture soils from the Amazon basin. Water Air Soil Pollut. Focus 2001, 1, 119–130. [Google Scholar] [CrossRef]
- Yu, J.B.; Meixner, F.X.; Sun, W.D.; Mamtimin, B.; Xia, C.H.; Xie, W.J. Biogenic Nitric Oxide Emission of Mountain Soils Sampled from Different Vertical Landscape Zones in the Changbai Mountains, Northeastern China. Environ. Sci. Technol. 2010, 44, 4122–4128. [Google Scholar] [CrossRef]
- Pilegaard, K.; Hummelshoj, P.; Jensen, N.O. Nitric oxide emission from a Norway spruce forest floor. J. Geophys. Res. Atmos. 1999, 104, 3433–3445. [Google Scholar] [CrossRef]
- Gharahi Ghehi, N.; Werner, C.; Cizungu Ntaboba, L.; Mbonigaba Muhinda, J.J.; Van Ranst, E.; Butterbach-Bahl, K.; Kiese, R.; Boeckx, P. Spatial variations of nitrogen trace gas emissions from tropical mountain forests in Nyungwe, Rwanda. Biogeosciences 2012, 9, 1451–1463. [Google Scholar] [CrossRef]
- Luo, G.J.; Bruggemann, N.; Wolf, B.; Gasche, R.; Grote, R.; Butterbach-Bahl, K. Decadal variability of soil CO2, NO, N2O, and CH4 fluxes at the Hoglwald Forest, Germany. Biogeosciences 2012, 9, 1741–1763. [Google Scholar] [CrossRef]
- Vourlitis, G.L.; DeFotis, C.; Kristan, W. Effects of soil water content, temperature and experimental nitrogen deposition on nitric oxide (NO) efflux from semiarid shrubland soil. J. Arid Environ. 2015, 117, 67–74. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F. The Temperature-Dependence of Soil Organic-Matter Decomposition, and the Effect of Global Warming on Soil Organic-C Storage. Soil Biol. Biochem. 1995, 27, 753–760. [Google Scholar] [CrossRef]
- Parker, S.S.; Schimel, J.P. Soil nitrogen availability and transformations differ between the summer and the growing season in a California grassland. Appl. Soil Ecol. 2011, 48, 185–192. [Google Scholar] [CrossRef]
- Sullivan, B.W.; Selmants, P.C.; Hart, S.C. New evidence that high potential nitrification rates occur in soils during dry seasons: Are microbial communities metabolically active during dry seasons? Soil Biol. Biochem. 2012, 53, 28–31. [Google Scholar] [CrossRef]
- Venterea, R.T.; Rolston, D.E.; Cardon, Z.G. Effects of soil moisture, physical, and chemical characteristics on abiotic nitric oxide production. Nutr. Cycl. Agroecosyst. 2005, 72, 27–40. [Google Scholar] [CrossRef]
- Gardner, P.R.; Gardner, A.M.; Martin, L.A.; Salzman, A.L. Nitric oxide dioxygenase: An enzymic function for flavohemoglobin. Proc. Natl. Acad. Sci. USA 1998, 95, 10378–10383. [Google Scholar] [CrossRef] [PubMed]
- Gusarov, I.; Gautier, L.; Smolentseva, O.; Shamovsky, I.; Eremina, S.; Mironov, A.; Nudler, E. Bacterial Nitric Oxide Extends the Lifespan of C. elegans. Cell 2013, 152, 818–830. [Google Scholar] [CrossRef] [PubMed]
- Gardner, A.M.; Helmick, R.A.; Gardner, P.R. Flavorubredoxin, an inducible catalyst for nitric oxide reduction and detoxification Escherichia coli. J. Biol. Chem. 2002, 277, 8172–8177. [Google Scholar] [CrossRef]
- Van Spanning, R.J.M.; Richardson, D.; Ferguson, S. Introduction to the biochemistry and molecular biology of denitrification. In Biology of the Nitrogen Cycle; Bothe, H., Ferguson, S., Newton, W.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Nadeem, S.; Dorsch, P.; Bakken, L.R. Autoxidation and acetylene-accelerated oxidation of NO in a 2-phase system: Implications for the expression of denitrification in ex situ experiments. Soil Biol. Biochem. 2013, 57, 606–614. [Google Scholar] [CrossRef]
- Koschorreck, M.; Conrad, R. Kinetics of nitric oxide consumption in tropical soils under oxic and anoxic conditions. Biol. Fertil. Soils 1997, 25, 82–88. [Google Scholar] [CrossRef]
- Remde, A.; Ludwig, J.; Meixner, F.X.; Conrad, R. A Study to Explain the Emission of Nitric-Oxide from a Marsh Soil. J. Atmos. Chem. 1993, 17, 249–275. [Google Scholar] [CrossRef]
- Otter, L.B.; Yang, W.X.; Scholes, M.C.; Meixner, F.X. Nitric oxide emissions from a southern African savanna. J. Geophys. Res. Atmos. 1999, 104, 18471–18485. [Google Scholar] [CrossRef]
- Feig, G.T.; Mamtimin, B.; Meixner, F.X. Soil biogenic emissions of nitric oxide from a semi-arid savanna in South Africa. Biogeosciences 2008, 5, 1723–1738. [Google Scholar] [CrossRef]
- Johansson, C. Field-Measurements of Emission of Nitric-Oxide from Fertilized and Unfertilized Forest Soils in Sweden. J. Atmos. Chem. 1984, 1, 429–442. [Google Scholar] [CrossRef]
- Wolf, I.; Russow, R. Different pathways of formation of N2O, N2 and NO in black earth soil. Soil Biol. Biochem. 2000, 32, 229–239. [Google Scholar] [CrossRef]
- Betlach, M.R.; Tiedje, J.M. Kinetic Explanation for Accumulation of Nitrite, Nitric-Oxide, and Nitrous-Oxide during Bacterial Denitrification. Appl. Environ. Microb. 1981, 42, 1074–1084. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wang, J.; Wang, S.Q.; Cai, Z.C.; Wang, L. Effects of temperature change and tree species composition on N2O and NO emissions in acidic forest soils of subtropical China. J. Environ. Sci. China 2014, 26, 617–625. [Google Scholar] [CrossRef]
- Davidson, E.A.; Nepstad, D.C.; Ishida, F.Y.; Brando, P.M. Effects of an experimental drought and recovery on soil emissions of carbon dioxide, methane, nitrous oxide, and nitric oxide in a moist tropical forest. Glob. Chang. Biol. 2008, 14, 2582–2590. [Google Scholar] [CrossRef]
- van Lent, J.; Hergoualc’h, K.; Verchot, L.V. Reviews and syntheses: Soil N2O and NO emissions from land use and land-use change in the tropics and subtropics: A meta-analysis. Biogeosciences 2015, 12, 7299–7313. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | Horizon | Depth (cm) | NH4+-N (mg kg−1 dw) | NO3−-N (mg kg−1 dw) | pHH2O | TOC (g kg−1) | TN (g kg−1) | C/N | Bulk Density (kg m−3) † | Particle Density (kg m−3) † | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| HS | T0 T1 T3 T5 | O (F and H layers) | ~0–1 | 82.1 (0.4) 63.7 (3.6) 66.8 (1.7) 90.6 (2.8) | 17.9 (0.9) 31.6 (1.9) 26.7 (1.0) 14.5 (0.7) | 4.4 (0.0) 5.1 (0.0) 4.2 (0.0) 4.3 (0.0) | 230 285 250 225 | 11.6 12.8 12.3 10.0 | 19.8 22.3 20.3 22.5 | 300 300 300 370 | 1600 |
| HS-Mean (SD) | 75.8 (11.7) | 22.7 (7.1) | 4.5 (0.4) | 248 (27) | 11.7 (1.2) | 21.2 (1.4) | 318 (35) | ||||
| GDZ | B2 B5 B6 | A | ~0–2 | 22.3 (0.9) 10.2 (0.8) 8.3 (0.6) | <0.01 mg N L−1 ‡ 1.2 (0.0) 0.8 (0.0) | 5.3 (0.0) 4.6 (0.0) 5.2 (0.0) | 29.4 62.5 42.7 | 2.0 4.7 3.7 | 14.7 13.3 11.5 | 1330 920 1000 | 2600 |
| GDZ-Mean (SD) | 13.6 (6.6) | 1.0 (0.2) | 5.0 (0.3) | 44.9 (16.7) | 3.5 (1.4) | 13.2 (1.6) | 1083 (217) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, R.; Behrendt, T.; Mulder, J.; Dörsch, P. Soil Moisture Control of NO Turnover and N2O Release in Nitrogen-Saturated Subtropical Forest Soils. Forests 2022, 13, 1291. https://doi.org/10.3390/f13081291
Kang R, Behrendt T, Mulder J, Dörsch P. Soil Moisture Control of NO Turnover and N2O Release in Nitrogen-Saturated Subtropical Forest Soils. Forests. 2022; 13(8):1291. https://doi.org/10.3390/f13081291
Chicago/Turabian StyleKang, Ronghua, Thomas Behrendt, Jan Mulder, and Peter Dörsch. 2022. "Soil Moisture Control of NO Turnover and N2O Release in Nitrogen-Saturated Subtropical Forest Soils" Forests 13, no. 8: 1291. https://doi.org/10.3390/f13081291
APA StyleKang, R., Behrendt, T., Mulder, J., & Dörsch, P. (2022). Soil Moisture Control of NO Turnover and N2O Release in Nitrogen-Saturated Subtropical Forest Soils. Forests, 13(8), 1291. https://doi.org/10.3390/f13081291