
A Bioactive Compounds Profile Present in the Selected Wood Rot

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.2. The Extracts Preparation
2.3. Chemical Analyzes
2.3.1. Determination of the Phenolic Compounds
Alkaline Hydrolysis
Acid Hydrolysis
2.3.2. Determination of the Free Phenolic Acids (FPA) and Antioxidant Activity (ABTS Method)
2.3.3. Determination of the Carotenoids
2.3.4. Determination of the Ergosterol (ERG)
2.4. Statistical Analysis
3. Results
3.1. Concentration of Phenolic Compounds
3.2. Antioxidant Activities of Mushroom Isolate Extracts
3.3. Concentration of Carotenoids
3.4. Concentration of Ergosterol
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wolna-Maruwka, A.; Piotrowska, M.; Niżewski, P. Zmiany liczebności grzybów pleśniowych oraz aktywności proteolitycznej osadu ściekowego kompostowanego w warunkach kontrolowanych. Nauka Przyr. Technol. 2010, 4, 6. [Google Scholar]
- Kulik, T.; Stuper-Szablewska, K.; Bilska, K.; Busko, M.; Ostrowska-Kołodziejczak, A.; Załuski, D.; Perkowski, J. Trans-Cinnamic and Chlorogenic Acids Affect the Secondary Metabolic Profiles and Ergosterol Biosynthesis by Fusarium culmorum and F. graminearum Sensu Stricto. Toxins 2017, 9, 198. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. Medicinal mushroom science: Current perspectives, advances, evidences and challenges. Biomed. J. 2014, 37, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Yong, Y.; Gu, Y.; Zhang, S.; Lu, L. Comparison of antioxidant and antiproliferation activities of polysaccharides from eight species of medicinal mushrooms. Int. J. Med. Mushrooms 2015, 17, 287–295. [Google Scholar] [CrossRef]
- Waktola, G.; Temesgen, T. Application of mushroom as food and medicine. Adv. Biotechnol. Microbiol. 2018, 11, 555817. [Google Scholar]
- Wu, F.; Zhou, L.W.; Vlasák, J.; Dai, Y.C. Global diversity and systematics of Hymenochaetaceae with poroid hymenophore. Fungal Divers. 2022, 113, 1–192. [Google Scholar] [CrossRef]
- Wojewoda, W. Checklist of Polish Larger Basidiomycetes. In Krytyczna Lista Wielkoowocnikowych Grzybów Podstawkowych Polski; Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2003; ISBN 83-89648-09-1. [Google Scholar]
- Zadrazil, F.; Kurtzman, R.H. The Biology of Pleurotus Cultivation in the Tropics in Tropical Mushrooms; Chang, S.T., Quimio, T.H., Eds.; The Chinese Press: Hong Kong, China, 1982; Volume 493, pp. 277–298. [Google Scholar]
- Gerhardt, E. Grzyby: Wielki Ilustrowany Przewodnik: Ponad 1000 Opisanych Gatunków; Łukomski, S., Ed.; Bauer-Weltbild Media: Warsaw, Poland, 2006; ISBN 83-7404-513-2. [Google Scholar]
- Sturion, G.L.; Oetterer, M. Composição química de cogumelos comestíveis (Pleurotus spp.) originados de cultivos em diferentes substratos. Ciência Tecnol. Aliment. 1995, 15, 189–193. [Google Scholar]
- Justo, M.B.; Guzmán, G.A.; Mejía, E.G.; Díaz, C.L.G.; Martínez, G.; Corona, E.B. Composition química de tres cepas mexicanas de setas (Pleurotus ostreatus). Arch. Latinoam. Nutr. 1998, 48, 359–363. [Google Scholar]
- Manzi, P.; Gambelli, L.; Marconi, S.; Vivanti, V.; Pizzoferrato, L. Nutrients in edible mushrooms: An inter-species comparative study. Food Chem. 1999, 65, 477–482. [Google Scholar] [CrossRef]
- Sadler, M. Nutritional properties of edible fungi. Br. Nutr. Found. Nutr. Bull. 2003, 28, 305–308. [Google Scholar] [CrossRef]
- Khan, M.A.; Amin, S.M.R.; Uddin, M.N.; Tania, M.; Alam, N. Comparative study of the nutritional composition of oyster mushrooms cultivated in Bangladesh. Bangladesh J. Mushroom 2008, 2, 9–14. [Google Scholar]
- Chihara, G. Immunopharmacology of lentinan, a polysaccharide isolated from Lentinus edodes: Its application as a host defense potentiator. Int. J. Orient. Med. 1992, 17, 55–77. [Google Scholar]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers. 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Cohen, R.; Persky, L.; Hadar, Y. Biotechnological applications and potential of wood-degrading mushrooms of the genus Pleurotus. Appl. Microbiol. Biotechnol. 2002, 58, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Bobek, P.; Galbavy, S. Effect of pleuran (beta-glucan from Pleurotus ostreatus) on the antioxidant status of the organism and on dimethylhydrazine-induced precancerous lesions in rat colon. Br. J. Biomed. Sci. 2001, 58, 164–168. [Google Scholar]
- Gumińska, B.; Wojewoda, W. Grzyby I Ich Oznaczanie; PWRiL: Warszawa, Poland, 1985; ISBN 83-09-00714-0. [Google Scholar]
- McQuilken, M.P.; Gemmell, J.; Whipps, J.M. Some Nutritional Factors Affecting Production of Biomass and Antifungal Metabolites of Coniothyrium minitans. Biocontrol Sci. Technol. 2010, 12, 443–454. [Google Scholar] [CrossRef]
- Sabat, J.; Gupta, N. Nutritional factors affecting the antifungal activity of Penicillium steckii of mangrove origin. Afr. J. Microbiol. Res. 2010, 4, 126–135. [Google Scholar]
- Manjunathan, J.; Kaviyarasan, V. Optimization of mycelia growth and antimicrobial activity of new edible mushroom, Lentinus tuberregium (Fr.). Tamil Nadu, India. Int. J. PharmTech Res. 2011, 3, 497–504. [Google Scholar]
- Nicolotti, G.; Varese, G. Screening of antagonistic fungi against air-borne infection by Heterobasidion annosum on Norway spruce. For. Ecol. Manag. 1996, 88, 249–257. [Google Scholar] [CrossRef]
- Łakomy, P.; Świdkiewicz, T.; Zieniewicz, J. Different influence of Hypholoma fasciculare (Huds: Fr.) Kummer on the growth of Armillaria ostoyae (Romagnesi) Herink in vitro. Phytopathol. Pol. 1999, 17, 91–98. [Google Scholar]
- Nicolotti, G.; Gonthier, P.; Varese, G.C. Effectiveness of some biocontrol and chemical treatments against Heterobasidion annosum on Norway spruce stumps. For. Pathol. 1999, 29, 339–346. [Google Scholar] [CrossRef]
- Woods, C.M.; Woodward, S.; Redfern, D.B. In vitro interactions in artificial and wood-based media between fungi colonizing stumps of Sitka spruce. For. Pathol. 2005, 35, 213–229. [Google Scholar] [CrossRef]
- de Boer, W.; Folman, L.B.; Gunnewiek, P.J.K.; Svensson, T.; Bastviken, D.; Öberg, G.; del Rio, J.C.; Boddy, L. Mechanism of antibacterial activity of the white-rot fungus Hypholoma fasciculare colonizing wood. Can. J. Microbiol. 2010, 56, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.C. Polypore diversity in China with an annotated checklist of Chinese polypores. Mycoscience 2012, 53, 49–80. [Google Scholar] [CrossRef]
- Łakomy, P.; Kwaśna, H. Atlas Hub; Multico: Warszawa, Poland, 2008; pp. 142–143. [Google Scholar]
- Kamperidou, V. The Biological Durability of Thermally-and Chemically-Modified Black Pine and Poplar Wood Against Basidiomycetes and Mold Action. Forests 2019, 10, 1111. [Google Scholar] [CrossRef]
- Staszczak, M.; Zdunek, E.; Leonowicz, A. Studies on the role of proteases in the white-rot fungus Trametes versicolor: Effect of PMSF and chloroquine on ligninolytic enzymes activity. J. Basic Microbiol. 2000, 40, 1. [Google Scholar] [CrossRef]
- Binion, D.E.; Stephenson, S.L.; Roody, W.C.; Burdsall, H.H.; Vasilyeva, L.N.; Miller, O.K. Macrofungi Associated with Oaks of Eastern North America; West Virginia University Press: Morgantown, WV, USA, 2008. [Google Scholar]
- Journal of Laws, No. of 2014, Item 1408. Available online: https://pbsociety.org.pl/journals/index.php/am/article/view/am.1094 (accessed on 2 July 2022).
- Zheng, W.; Zhang, M.; Zhao, Y.; Wang, Y.; Miao, K.; Wei, Z. Accumulation of antioxidant phenolic constituents in submerged cultures of Inonotus obliquus. Bioresour. Technol. 2009, 100, 1327–1335. [Google Scholar] [CrossRef]
- Mizuno, T.; Zhuang, C.; Abe, K.; Okamoto, H.; Kiho, T.; Ukai, S.; Meijer, L. Antitumor and hypoglycemic activities of polysaccharides from the sclerotia and mycelia of Inonotus obliquus (Pers.: Fr.) Pil. (Aphyllophoromycetideae). Int. J. Med. Mushrooms 1999, 1, 301–316. [Google Scholar] [CrossRef]
- Kahlos, K.; Lesnau, A.; Lange, W.; Lindequist, U. Preliminary tests of antiviral activity of two Inonotus obliquus strains. Fitoterapia 1996, 67, 344–347. [Google Scholar]
- Ichimura, T.; Watanabe, O.; Maruyama, S. Inhibition of HIV-1 Protease by Water-Soluble Lignin-Like Substance from an Edible Mushroom, Fuscoporia obliqua. Biosci. Biotechnol. Biochem. 1998, 62, 575–577. [Google Scholar] [CrossRef]
- Ham, S.-S.; Kim, S.-H.; Moon, S.-Y.; Chung, M.J.; Cui, C.-B.; Han, E.-K.; Chung, C.-K.; Choe, M.S.; Kim, S.-H.; Moon, S.-Y.; et al. Antimutagenic effects of subfractions of Chaga mushroom (Inonotus obliquus) extract. Mutat. Res. Toxicol. Environ. Mutagen. 2009, 672, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Kahlos, K.; Kangas, L.; Hiltunen, R. Antitumor activity of some compounds and fractions from an n-hexane extract of Inonotus obiquus. Acta Pharm. Fenn. 1987, 96, 33–40. [Google Scholar]
- Jarosz, A.; Skórska, M.; Rzymowska, J.; KochmańSka-Rdest, J.; Malarczyk, E. Effect of the extracts from fungus Inonotus obliquus on catalase level in HeLa and nocardia cells. Acta Biochim. Pol. 1990, 37, 149–151. [Google Scholar] [PubMed]
- Kahlos, K. Inonotus obliquus (Chaga Fungus): In vitro culture and the production of inotodiol, sterols, and other secondary metabolites. In Medicinal and Aromatic Plants VI; Springer: Berlin/Heidelberg, Germany, 1994; pp. 179–198. [Google Scholar]
- Burczyk, J.; Gawron, A.; Slotwinska, M.; Smietana, B.; Terminska, K. Antimitotic activity of aqueous extracts of Inonotus obliquus. Boll. Chim. Farm. 1996, 135, 306–309. [Google Scholar]
- Rzymowska, J. The effect of aqueous extracts from Inonotus obliquus on the mitotic index and enzyme activities. Boll. Chim. Farm. 1998, 137, 13–15. [Google Scholar]
- Jiang, O.P.; Don, Y.; Feng, Y.J.; Bondartseva, M.A.; Gao, T.H.; Chen, F.M. The antitumor activity and M.D.R. reversal properties of constituents from Inoxotus obliquus. Микoлoгия Фитoпатoлoгия 2007, 41, 455–460. [Google Scholar]
- Song, Y.; Hui, J.; Kou, W.; Xin, R.; Jia, F.; Wang, N.; Hu, F.; Zhang, H.; Liu, H. Identification of Inonotus obliquus and Analysis of Antioxidation and Antitumor Activities of Polysaccharides. Curr. Microbiol. 2008, 57, 454–462. [Google Scholar] [CrossRef]
- Chung, M.J.; Chung, C.-K.; Jeong, Y.; Ham, S.-S. Anticancer activity of subfractions containing pure compounds of Chaga mushroom (Inonotus obliquus) extract in human cancer cells and in Balbc/c mice bearing Sarcoma-180 cells. Nutr. Res. Pr. 2010, 4, 177–182. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Yu, C.J.; Zhou, L.W. Species diversity and utilization of medicinal mushrooms and fungi in China. Int J. Med. Mushrooms 2009, 11, 287–302. [Google Scholar] [CrossRef]
- Cui, Y.; Kim, D.-S.; Park, K.-C. Antioxidant effect of Inonotus obliquus. J. Ethnopharmacol. 2005, 96, 79–85. [Google Scholar] [CrossRef]
- Nakajima, Y.; Nishida, H.; Matsugo, S.; Konishi, T. Cancer Cell Cytotoxicity of Extracts and Small Phenolic Compounds from Chaga [Inonotus obliquus (persoon) Pilat]. J. Med. Food 2009, 12, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Sato, Y.; Konishi, T. Antioxidant Small Phenolic Ingredients in Inonotus obliquus (persoon) Pilat (Chaga). Chem. Pharm. Bull. 2007, 55, 1222–1226. [Google Scholar] [CrossRef] [PubMed]
- Kawashte, G.; Sonawane, H.; Mittal, S.; Borde, M. Medicinal Fungi: A Natural Source of Pharmacologically Important Metabolites. In Recent Developments in Microbial Technologies; Springer: Singapore, 2021; pp. 379–394. ISBN 978-981-15-4439-2. [Google Scholar]
- Stuper-Szablewska, K.; Kurasiak-Popowska, D.; Nawracała, J.; Perkowski, J. Response of non enzymatic antioxidative mechanisms to stress caused by infection with Fusarium fungi and chemical protection in different wheat genotypes. Chem. Ecol. 2017, 33, 949–962. [Google Scholar] [CrossRef]
- Przybylska-Balcerek, A.; Frankowski, J.; Stuper-Szablewska, K. The influence of weather conditions on bioactive compound content in sorghum grain. Eur. Food Res. Technol. 2020, 246, 13–22. [Google Scholar] [CrossRef]
- Kurasiak-Popowska, D.; Ryńska, B.; Stuper-Szablewska, K. Analysis of Distribution of Selected Bioactive Compounds in Camelina sativa from Seeds to Pomace and Oil. Agronomy 2019, 9, 168. [Google Scholar] [CrossRef]
- Perkowski, J.; Buśko, M.; Stuper-Szablewska, K.; Kostecki, M.; Matysiak, A.; Szwajkowska-Michałek, L. Concentration of ergosterol in small-grained naturally contaminated and inoculated cereals. Biologia 2008, 63, 542–547. [Google Scholar] [CrossRef]
- Bravo, L. Polyphenols: Chemistry, dietary, sources, metabolism and nutrional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef]
- Mansouri, A.; Makris, D.P.; Kefalas, P. Determination of hydrogen peroxide scavenging activity of cinnamic and benzoic acids employing a highly sensitive peroxyoxalate chemiluminescence-based assay: Structure–activity relationships. J. Pharm. Biomed. Anal. 2005, 39, 22–26. [Google Scholar] [CrossRef]
- Karaman, M.; Jovin, E.; Malbaša, R.; Matavuly, M.; Popović, M. Medicinal and edible lignicolous fungi as natural sources of antioxidative and antibacterial agents. Phytotherapy Res. 2010, 24, 1473–1481. [Google Scholar] [CrossRef]
- Szymański, M.; Smolibowska, J.; Szymański, A. An investigation into the relationships between antioxidant activity and chemical elements as well as polyphenolics in fungal fruiting bodies growing on Betula L. J. Elem. 2019, 24, 1. [Google Scholar] [CrossRef]
- Glamočlija, J.; Ćirić, A.; Nikolić, M.; Fernandes, A.; Barros, L.; Calhelha, R.C.; Ferreira, I.C.F.R.; Soković, M.; van Griensven, L.J.L.D. Chemical characterization and biological activity of Chaga (Inonotus obliquus), a medicinal “mushroom”. J. Ethnopharmacol. 2015, 162, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Stojkovic, D.; Smiljkovic, M.; Ciric, A.; Glamoclija, J.; van Griensven, L.; Ferreira, I.C.F.R.; Sokovic, M. An insight into antidiabetic properties of six medicinal and edible mushrooms: Inhibition of α-amylase and α-glucosidase linked to type-2 diabetes. S. Afri. J. Bot. 2019, 120, 100–103. [Google Scholar] [CrossRef]
- Ju, H.K.; Chung, H.W.; Hong, S.S.; Park, J.H.; Lee, J.; Kwon, S.W. Effect of steam treatment on soluble phenolic content and antioxidant activity of the Chaga mushroom (Inonotus obliquus). Food Chem. 2010, 119, 619–625. [Google Scholar] [CrossRef]
- Janjušević, L.; Karaman, M.; Šibul, F.; Tommonaro, G.; Iodice, C.; Jakovljević, D.; Pejin, B. The lignicolous fungus Trametes versicolor (L.) Lloyd (1920): A promising natural source of antiradical and AChE inhibitory agents. J. Enzym. Inhib. Med. Chem. 2017, 32, 355–362, PMCID:PMC6010034. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, B.; Sułkowska-Ziaja, K.; Ekiert, H. Phenolic acids in selected edible Basidiomycotaspecies: Armillaria mellea, Boletus badius, Boletus edulis, Cantharellus cibarius, Lactarius deliciosus, and Pleurotus ostreatus. Acta Sci. Pol. Hortorum Cultus. 2013, 12, 12. [Google Scholar]
- Gąsecka, M.; Mleczek, M.; Siwulski, M.; Niedzielski, P. Phenolic composition and antioxidant properties of Pleurotus ostreatus and Pleurotus eryngii enriched with selenium and zinc. Eur. Food Res. Technol. 2015, 226, 737–743. [Google Scholar] [CrossRef]
- Kim, M.-Y.; Seguin, P.; Ahn, J.-K.; Kim, J.-J.; Chun, S.-C.; Kim, E.-H.; Seo, S.-H.; Kang, E.-Y.; Kim, S.-L.; Park, Y.-J.; et al. Phenolic Compound Concentration and Antioxidant Activities of Edible and Medicinal Mushrooms from Korea. J. Agric. Food Chem. 2008, 56, 7265–7270. [Google Scholar] [CrossRef]
- Badalyan, S.M. Edible and medicinal higher basidiomycete mushrooms as a source of natural antioxidants. Int. J. Med. Mushrooms. 2003, 5, 153–162. [Google Scholar] [CrossRef]
- Woldegiorgis, A.Z.; Abate, D.; Haki, G.D.; Ziegler, G.R. Antioxidant property of edible mushrooms collected from Ethiopia. Food Chem. 2014, 157, 30–36. [Google Scholar] [CrossRef]
- Palacios, I.; Lozano, M.; Moro, C.; D’Arrigo, M.; Rostagno, M.A.; Martínez, J.A.; García-Lafuente, A.; Guillamón, E.; Villares, A. Antioxidant properties of phenolic compounds occurring in edible mushrooms. Food Chem. 2011, 128, 674–678. [Google Scholar] [CrossRef]
- Barros, L.; Dueñas, M.; Ferreira, I.C.; Baptista, P.; Santos-Buelga, C. Phenolic acids determination by HPLC–D.A.D.–ESI/MS in sixteen different Portuguese wild mushrooms species. Food Chem. Toxicol. 2009, 47, 1076–1079. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.C.; Barros, L.; Abreu, R.M. Antioxidants in wild mushrooms. Curr. Med. Chem. 2009, 16, 1543–1560. [Google Scholar] [CrossRef] [PubMed]
- Sandmann, G.; Misawa, N. Fungal Carotenoids. In Industrial Applications. The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Osiewacz, H.D., Ed.; Springer: Berlin/Heidelberg, Germany, 2002; Volume 10. [Google Scholar] [CrossRef]
- Morris, S.A.C.; Subden, R.E. Effects of ultraviolet radiation on carotenoid containing and albino strains of Neurospora crassa. Mutat. Res. Mol. Mech. Mutagen. 1974, 22, 105–109. [Google Scholar] [CrossRef]
- Avalos, J.; Limón, M.C. Biological roles of fungal carotenoids. Curr. Genet. 2014, 61, 309–324, EPUB 2014 Paź 5. [Google Scholar] [CrossRef] [PubMed]
- Skibsted, L.H. Carotenoids in Antioxidant Networks. Colorants or Radical Scavengers. J. Agric. Food Chem. 2012, 60, 2409–2417. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.D.; Jaeager, C.; Ruck, C.; Schmidt, M.; Walsh, R.; Paust, J.J. Astaxanthin uptake in domestic dogs and cats. ESPEN 1999, 52, 1–8. [Google Scholar]
- Mishra, K.K.; Pal, R.S.; Arunkumar, R. Antioxidant activities and bioactive compound determination from caps and stipes of specialty medicinal mushrooms Calocybe indica and Pleurotus sajor-caju (higher Basidiomycetes) from India. Int. J. Med. Mushrooms 2014, 16, 555–567. [Google Scholar] [CrossRef]
- Jayakumar, T.; Thomas, P.A.; Geraldine, P. In vitro antioxidant activities of an ethanolic extract of the oyster mushroom, Pleurotus ostreatus. Innov. Food Scien. Emerg. Technol. 2009, 10, 228–234. [Google Scholar] [CrossRef]
- Robaszkiewicz, A.; Bartosz, G.; Lawrynowicz, M.; Soszynski, M. The role of polyphenols, β-carotene, and lycopene in the antioxidative action of the extract of dried edible mushrooms. J. Nutr. Metabol. 2010, 11, 173–274. [Google Scholar] [CrossRef]
- Jaworska, G.; Pogoń, K.; Bernaś, E.; Duda-Chodak, A. Nutraceuticals and Antioxidant Activity of Prepared for Consumption Commercial Mushrooms Agaricus bisporus and Pleurotus ostreatus. J. Food Qual. 2015, 38, 111–122. [Google Scholar] [CrossRef]
- Turfan, N.; Ayan, S.; Pekşen, A.; Akin, S. Antioxidant Enzyme Activities of Some Wild and Cultivated Edible Mushrooms in Turkey. Uluslararası Tarım Yaban Hayatı Bilimleri Derg. 2020, 6, 202–206. [Google Scholar] [CrossRef]
- Barros, L.; Cruz, T.; Baptista, P.; Estevinho, L.M.; Ferreira, I.C. Wild and commercial mushrooms as source of nutrients and nutraceuticals. Food Chem. Toxicol. 2008, 46, 2742–2747. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Venturini, B.A.; Baptista, P.; Estevinho, L.M.; Ferreira, I.C. Chemical composition and biological properties of Portuguese wild mushrooms: A comprehensive study. J. Agric. Food Chem. 2008, 56, 3856–3862. [Google Scholar] [CrossRef] [PubMed]
- Havsteen, B. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar]
- Pietta, P.-G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic compounds in plants and agri-industrial by-products: Antioxidant activity, occurrence, and potential uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Mattila, P. Sterol and vitamin D2 contents in some wild and cultivated mushrooms. Food Chem. 2002, 76, 293–298. [Google Scholar] [CrossRef]
- Shin, Y.; Tamai, Y.; Terazawa, M. Chemical Constituents of Inonotus obliquus: Triterpene and Steroids from Cultured Mycelia. Eurasian J. For. Res. 2001, 2, 27–30. [Google Scholar]
- Akanmu, D.; Cecchini, R.; Aruoma, O.I.; Halliwell, B. The antioxidant action of ergothioneine. Arch. Biochem. Biophys. 1991, 288, 10–16. [Google Scholar] [CrossRef]
- Matijašević, D.; Pantić, M.; Raskovic, B.; Pavlović, V.; Duvnjak, D.; Sknepnek, A.; Nikšić, M. The Antibacterial Activity of Coriolus versicolor Methanol Extract and Its Effect on Ultrastructural Changes of Staphylococcus aureus and Salmonella Enteritidis. Front. Microbiol. 2016, 7, 1226. [Google Scholar] [CrossRef]
- Vamanu, E.; Voica, A. Total phenolic analysis, antimicrobial and antioxidant activity of some mushroom tinctures from medicinal and edible species, by in vitro and in vivo tests. Sci. Bulletin. Ser. F. Biotechnol. 2017, 21, 318–324. [Google Scholar]
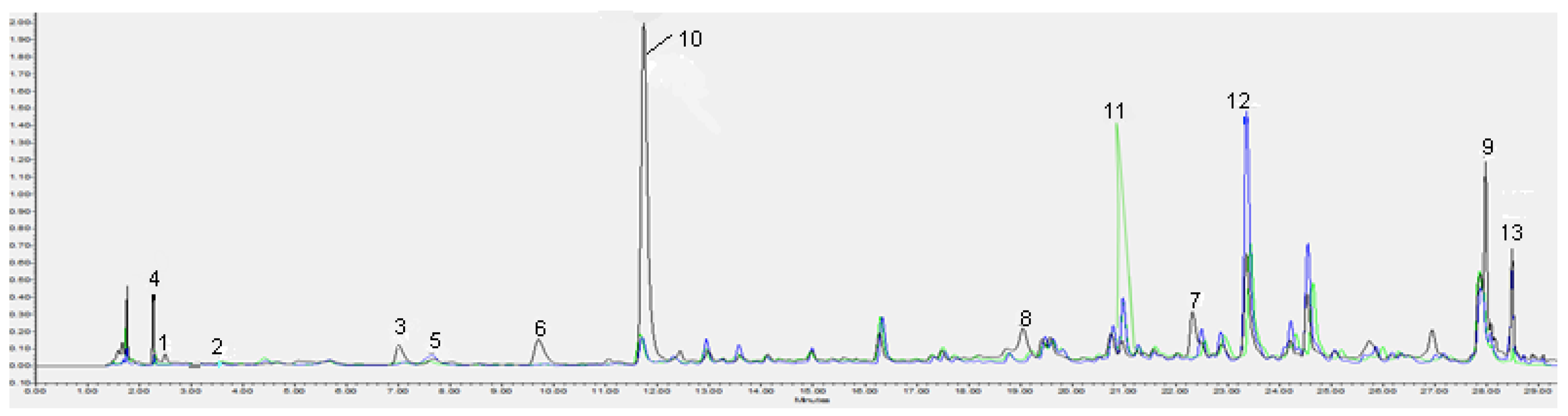

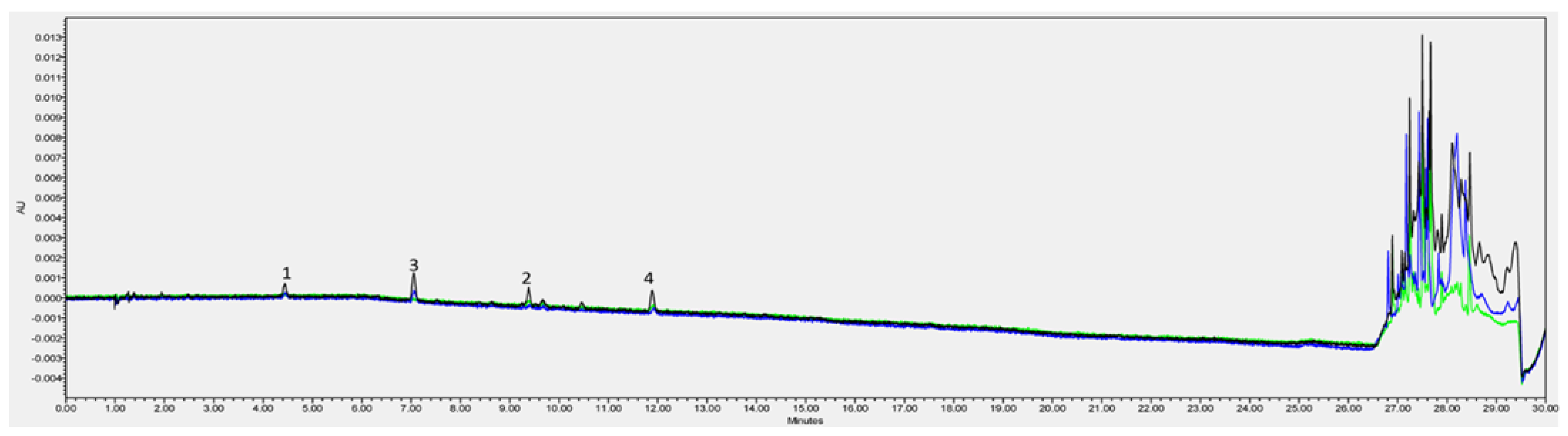

{kind=link}
{kind=link}
{kind=link}
| No | Species | Isolate No. | Isolate Code |
|---|---|---|---|
| 1. | Sulfur tuft (Hypholoma fasciculare) | 96,032 | Hf |
| 2. | Smoky polypore (Bjerkandera adusta) | 322 | Ba |
| 3. | Chaga (Inonotus obliquus) | 110,421.3 | Io |
| 4. | Sheathed woodtuft (Kuehneromyces mutabilis) | 96,040 | Km |
| 5. | Turkey tail (Trametes versicolor) | 230,318 | Tv |
| 6. | Oyster mushroom (Pleurotus ostreatus) | 20,080 | Po |
| 7. | Shaggy scalycap (Pholiota squarrosa) | 206,021 | Ps |
| Phenolic Acids | Hf | Ps | Ba | Io | Po | Tv | Km |
|---|---|---|---|---|---|---|---|
| Protocatechuic | 1.21a | 0.12c | 0.5b | 1.08a | 0.95a | 0.5b | 1.69a |
| Pyroglutaminic | 0.27ab | 0.09b | nd | 0.77a | nd | nd | 0.45a |
| Pyruvic | 1.34c | 0.12d | nd | 7.2b | nd | 10.95a | 6.56b |
| Gallic | 26.57a | 9.55b | 9.82b | 32.13a | 8.43b | 9.77b | 28.73a |
| 2.5-dihydroxybenzoic | 9.46a | 2.07b | 3.0b | 9.76a | 2.8b | 2.9b | 8.98a |
| 4-hydroxybenzoic | 11.22a | 1.45b | 2.0b | 9.45a | 1.71b | 1.78b | 12.69a |
| Caffeic | 31.73a | 3.31b | 0.01c | 31.62a | 0.02c | 0.01c | 32.58a |
| Syryngic | 28.84a | 2.92b | 0.09c | 30.64a | 0.08c | 0.06c | 29.82a |
| p-coumaric | 92.75a | 6.12b | 6.92b | 84.12a | 6.45b | 6.27b | 94.79a |
| Ferulic | 87.87a | 10.7b | 10.39b | 98.43a | 9.95b | 9.14b | 98.38a |
| Chlorogenic | 18.63b | 6.56c | 108.49a | 27.91b | 9.55c | 9.13c | 19.83b |
| Sinapic | 53.01b | 1.24c | 0.14c | 51.56b | 100.72a | 0.12c | 56.31b |
| t-cinnamic | 43.07a | 2.16b | 2.16b | 50.85a | 1.16b | 1.98b | 41.5a |
| Sum of an identified phenolic acids | 405.97a | 46.41c | 143.52b | 435.52a | 141.82b | 52.61c | 432.31a |
| Isolate Code | Flavonoids | |||||
|---|---|---|---|---|---|---|
| Naringenin | Vitexin | Rutin | Quercetin | Apigenin | Kaempferol | |
| Hf | 25.66a | 5.84a | 16.72a | 22.99b | 16.35a | 20.88a |
| Ps | 0.32c | nd | nd | nd | nd | nd |
| Ba | 0.07c | nd | nd | nd | nd | nd |
| Io | 27.11a | 5.18a | 19.05a | 24.03ab | 17.92a | 21.21a |
| Po | 0.14c | nd | nd | 0.33c | 6.99b | nd |
| Tv | 0.18c | 0.03b | nd | nd | nd | nd |
| Km | 16.82b | 6.62a | 19.02a | 27.50a | 19.57a | 24.63a |
| Isolate Code | FPA (mg GAE/g DM) | ABTS (TEAC) μM Trolox/g |
|---|---|---|
| Hf | 147.10b | 467.87b |
| Ps | 162.21c | 104.20a |
| Ba | 102.97a | 108.28a |
| Io | 188.99d | 351.41b |
| Po | 104.81a | 417.71b |
| Tv | 195.52d | 908.56c |
| Km | 171.62c | 534.27b |
| Isolate Code | Lutein | Zeaxanthin | β-Carotene | Astaxanthin | Sum of Identified Carotenoids |
|---|---|---|---|---|---|
| Hf | 0.15a | 0.05a | 0.37a | 0.10a | 0.90a |
| Ps | 0.11a | 0.04a | 0.19a | 0.05a | 0.65a |
| Ba | 0.11a | 0.08a | 0.30a | 0.10a | 0.99a |
| Io | 0.31b | 0.24b | 0.51ab | 0.21ab | 1.65b |
| Po | 0.57b | 0.37b | 0.70b | 0.21ab | 1.99b |
| Tv | 0.70c | 0.21b | 0.99b | 0.37b | 2.66c |
| Km | 0.03a | 0.07a | 0.26a | 0.10a | 0.46a |
| Isolate Code | Ergosterol |
|---|---|
| Hf | 772.50c |
| Ps | 300.63a |
| Ba | 317.92a |
| Io | 863.33c |
| Po | 342.18a |
| Tv | 223.30a |
| KM | 610.89b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szwajkowska-Michałek, L.; Stuper-Szablewska, K.; Krzyżaniak, M.; Łakomy, P. A Bioactive Compounds Profile Present in the Selected Wood Rot. Forests 2022, 13, 1242. https://doi.org/10.3390/f13081242
Szwajkowska-Michałek L, Stuper-Szablewska K, Krzyżaniak M, Łakomy P. A Bioactive Compounds Profile Present in the Selected Wood Rot. Forests. 2022; 13(8):1242. https://doi.org/10.3390/f13081242
Chicago/Turabian StyleSzwajkowska-Michałek, Lidia, Kinga Stuper-Szablewska, Michał Krzyżaniak, and Piotr Łakomy. 2022. "A Bioactive Compounds Profile Present in the Selected Wood Rot" Forests 13, no. 8: 1242. https://doi.org/10.3390/f13081242
APA StyleSzwajkowska-Michałek, L., Stuper-Szablewska, K., Krzyżaniak, M., & Łakomy, P. (2022). A Bioactive Compounds Profile Present in the Selected Wood Rot. Forests, 13(8), 1242. https://doi.org/10.3390/f13081242