
Towards Continuous Stem Water Content and Sap Flux Density Monitoring: IoT-Based Solution for Detecting Changes in Stem Water Dynamics

 , ,
, ,  , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Material and Methods
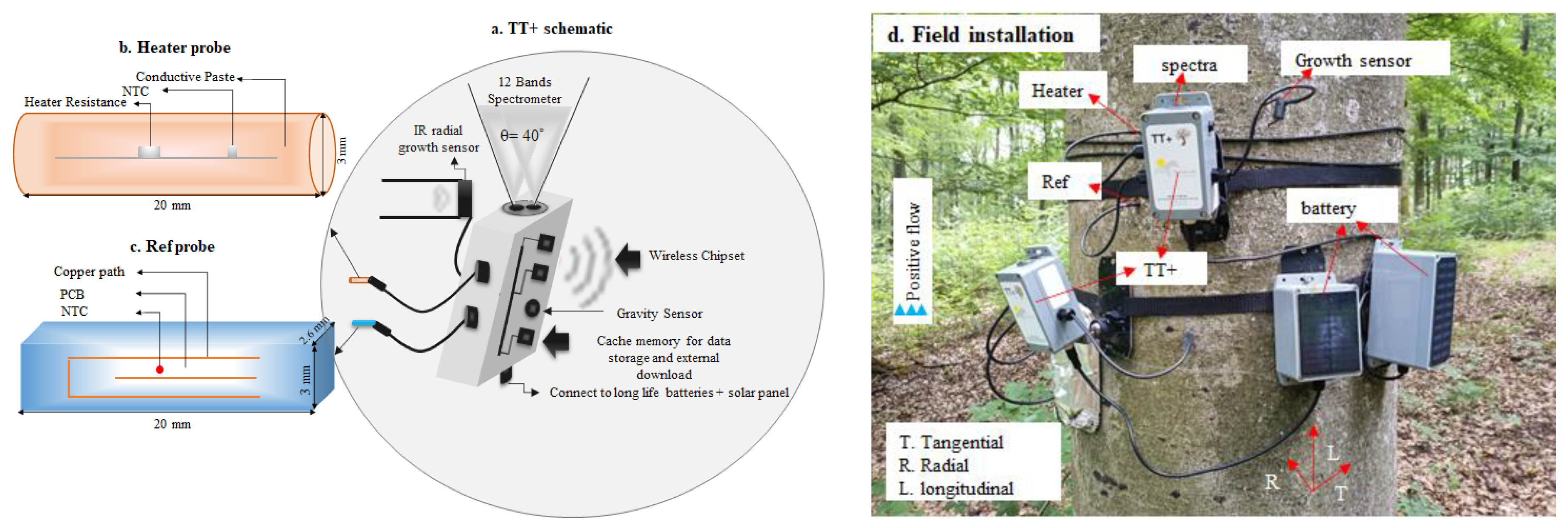
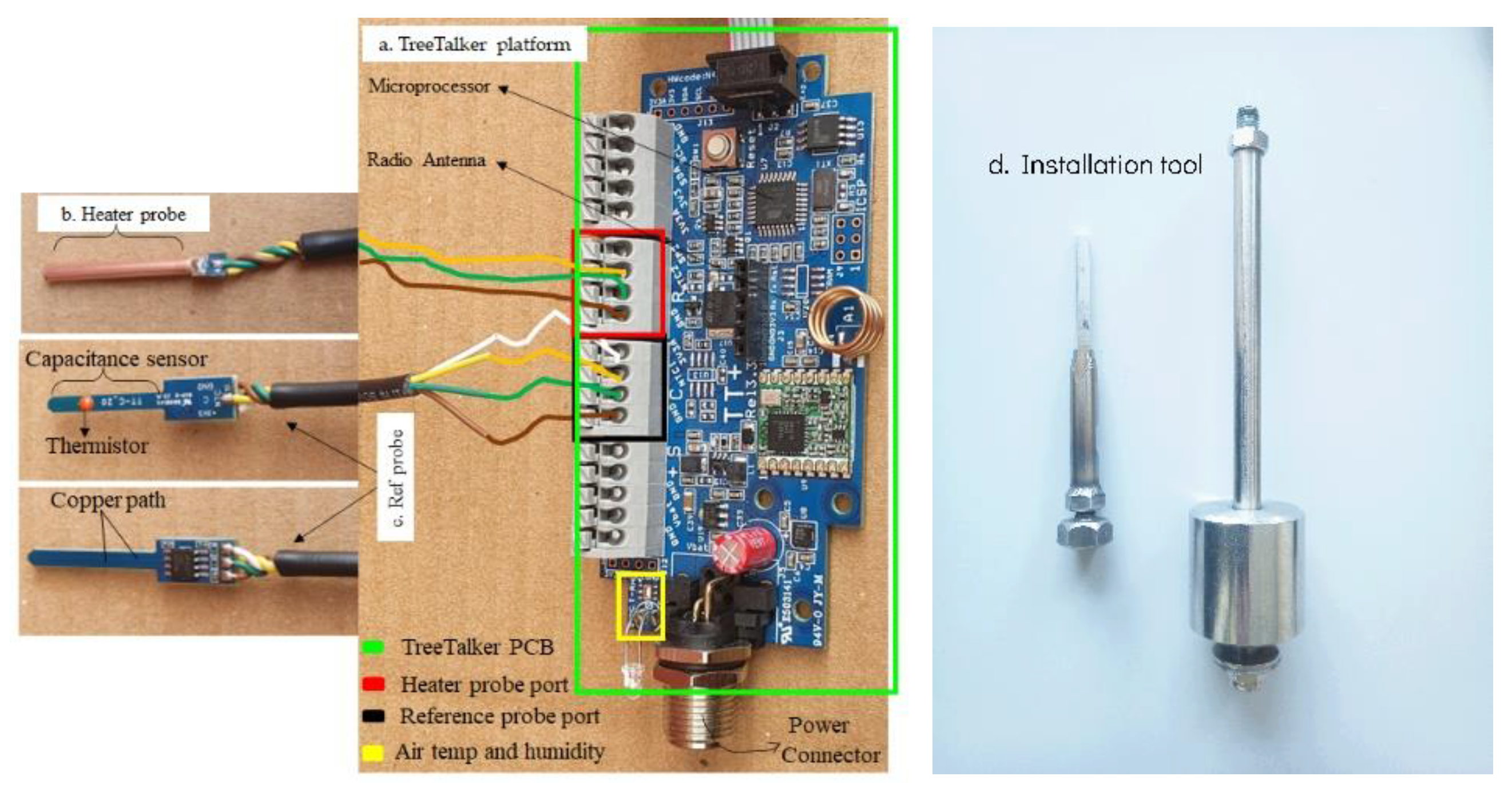
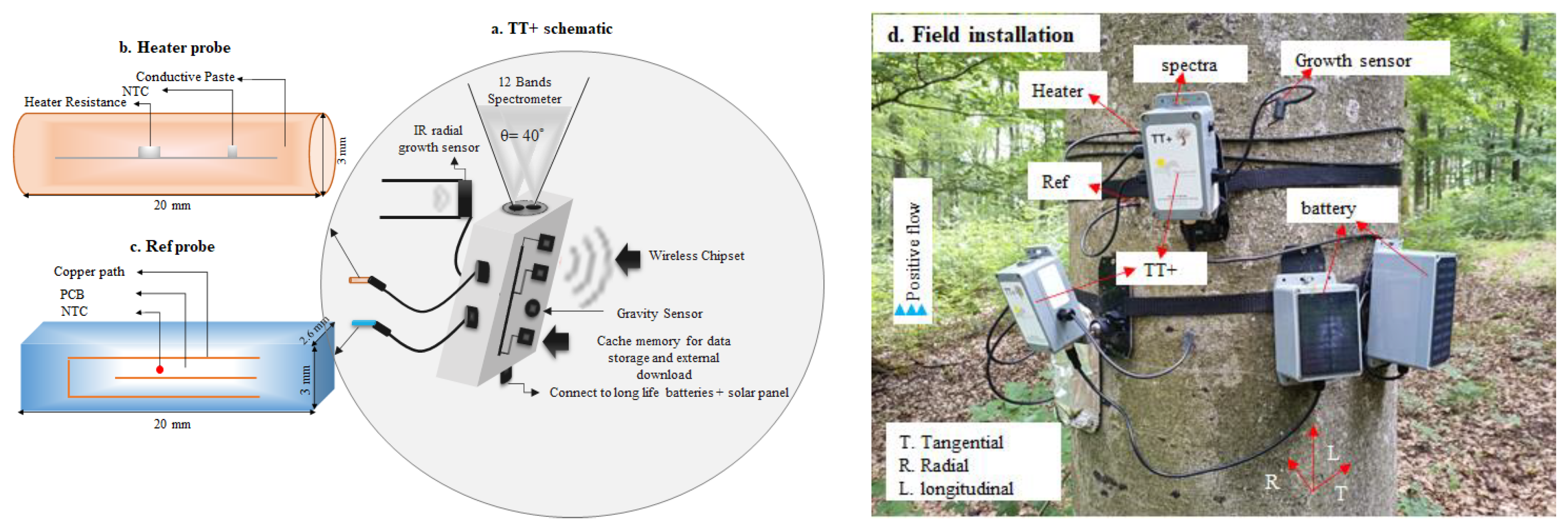
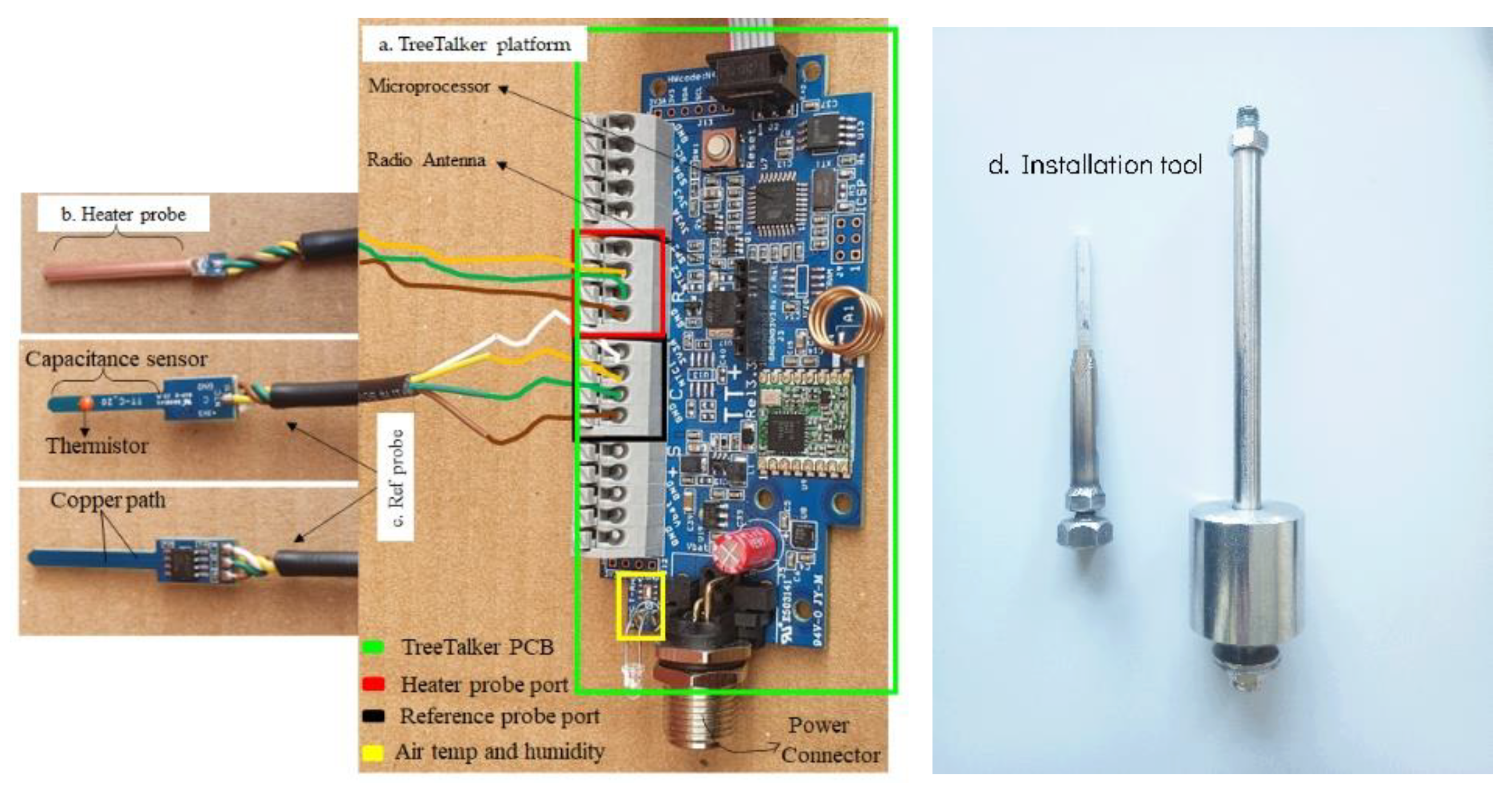
2.1. The TreeTalker Platform
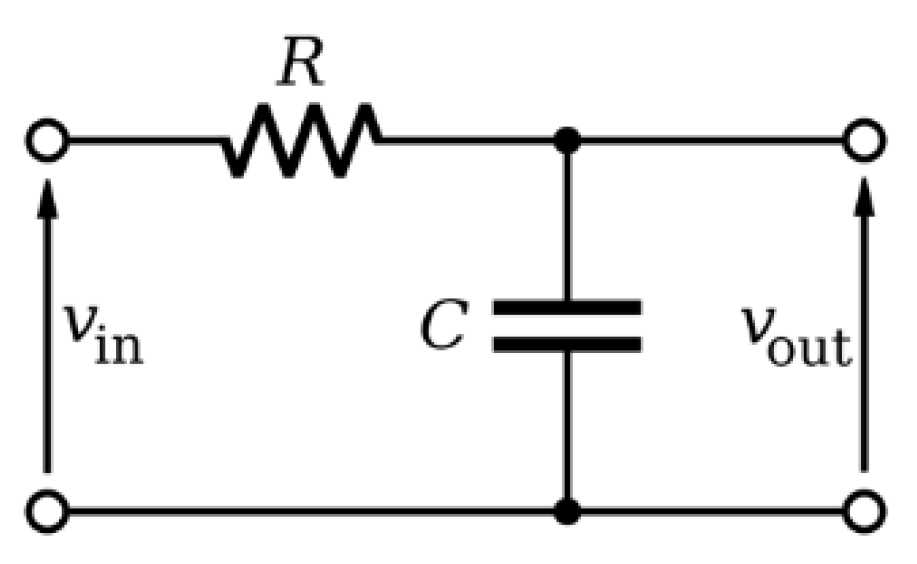
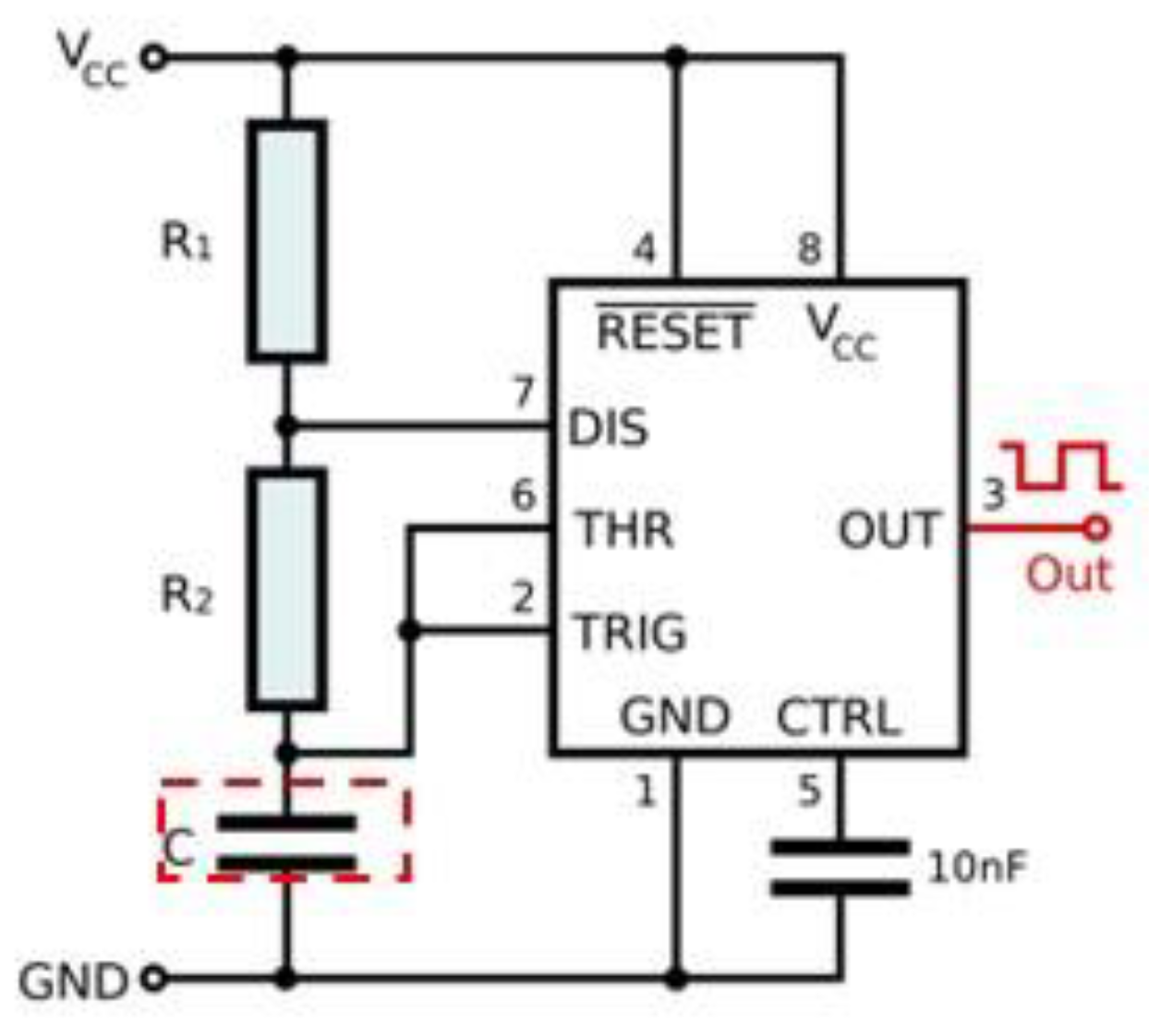
2.2. Sensor Design for Stem Water Content Monitoring
2.3. Lab Experiments
2.3.1. Temperature Dependence of Ref Probe
2.3.2. Species-Specific Calibration
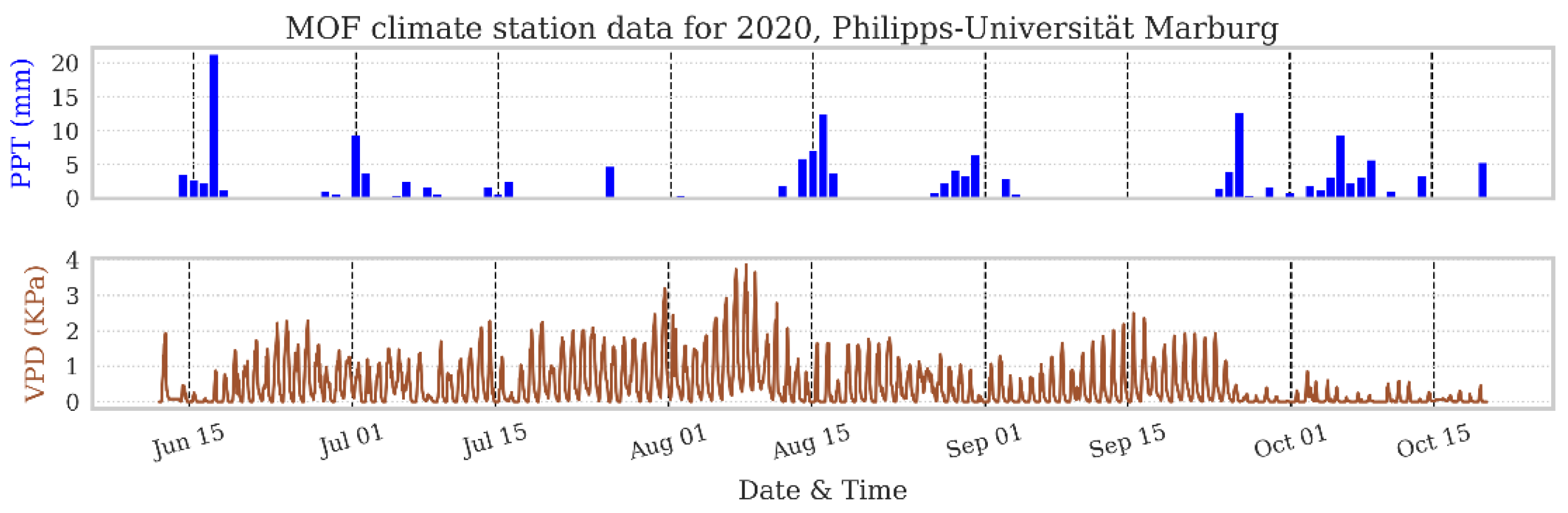
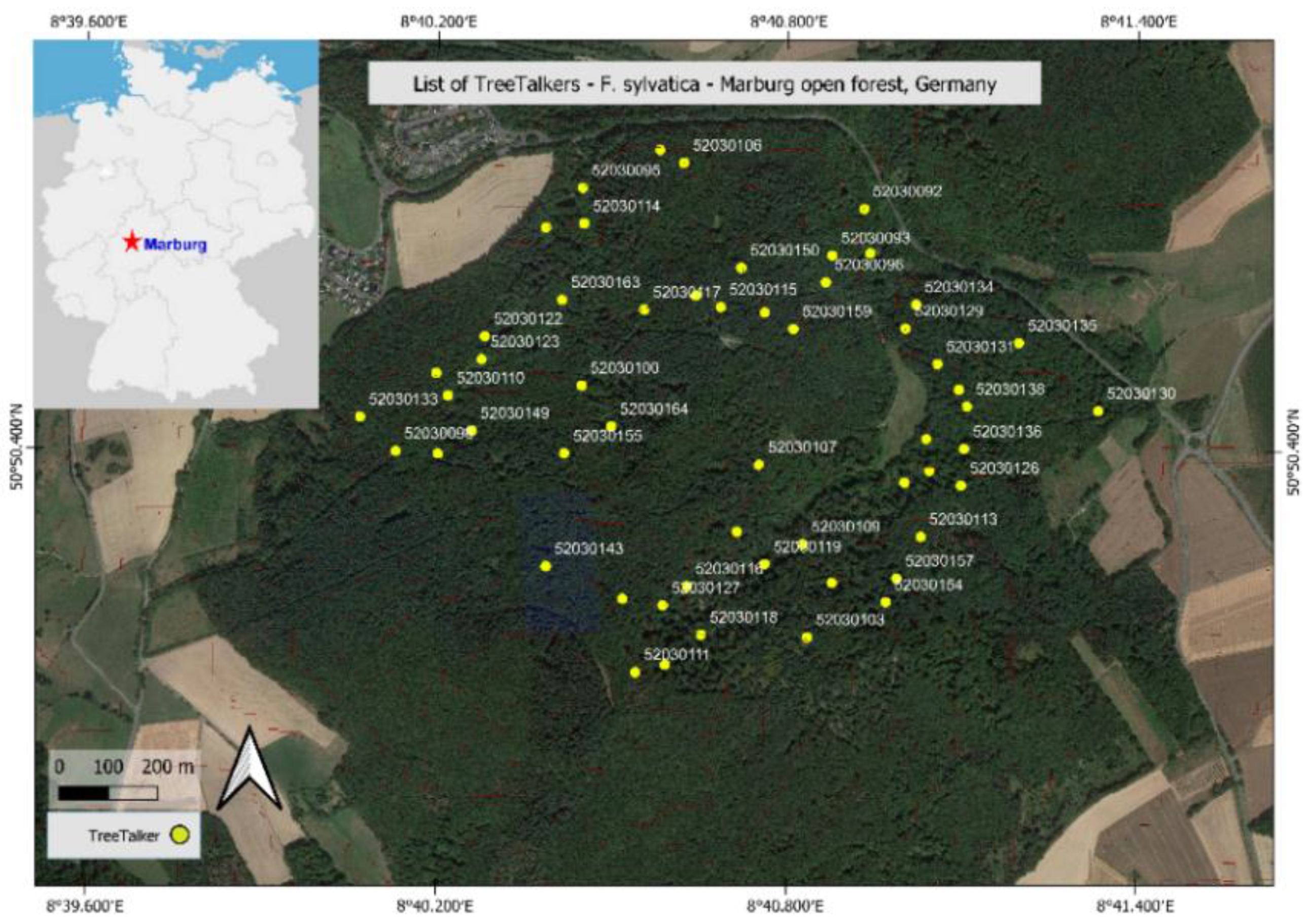
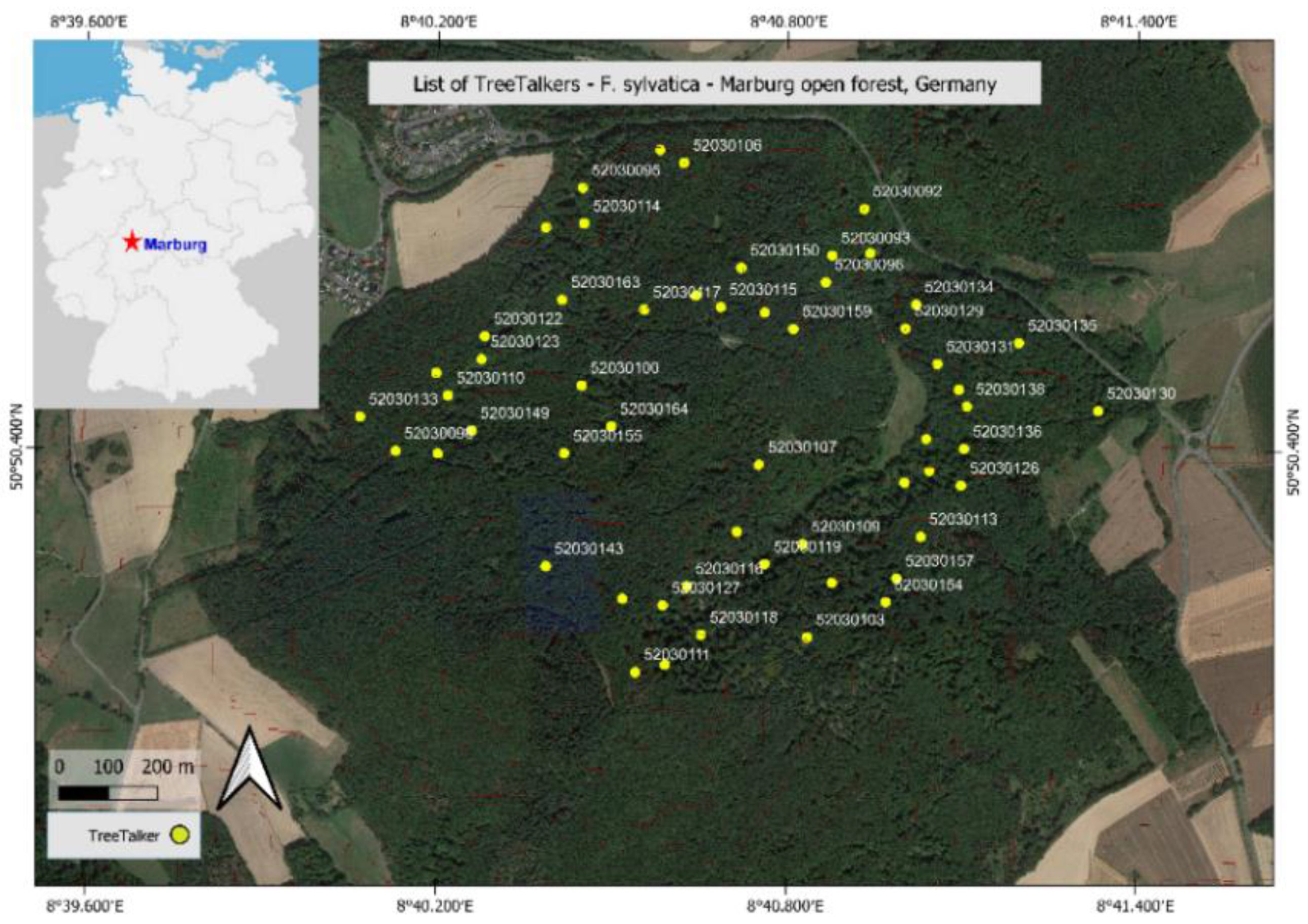
2.4. Study Site and Field Experiments
3. Results and Discussion
3.1. Lab Experiments
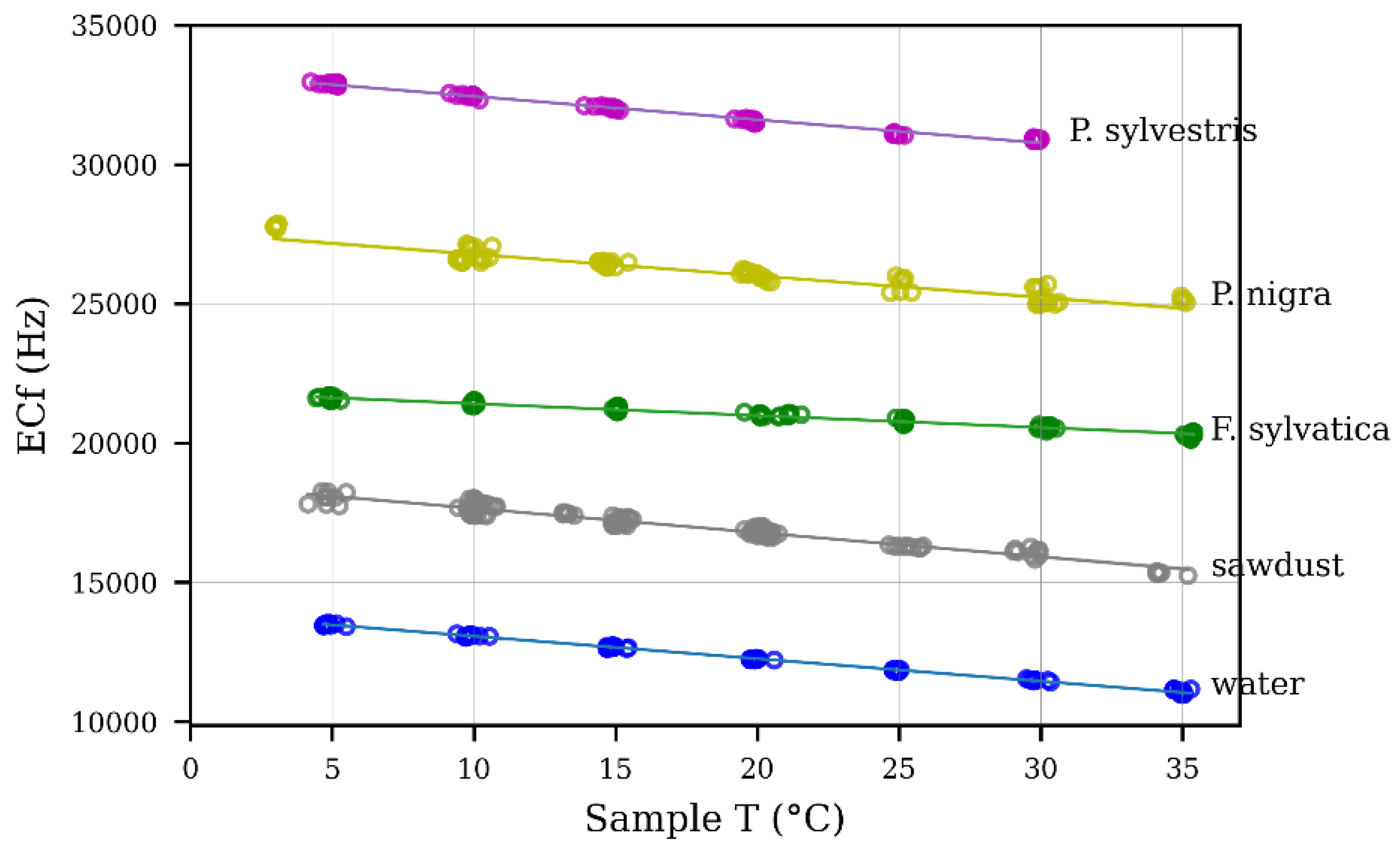
3.1.1. Temperature Sensitivity Analysis of Ref Probe
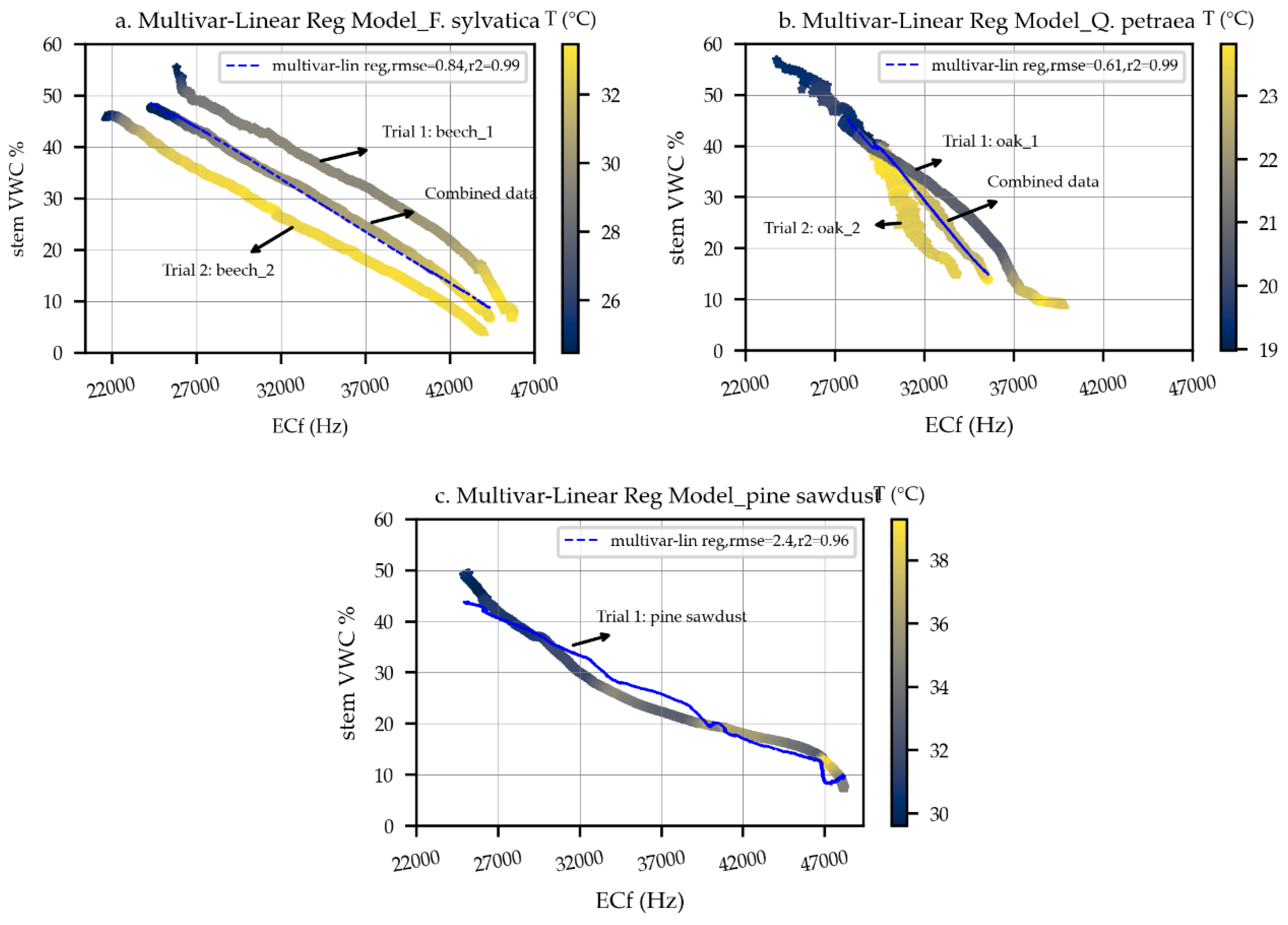
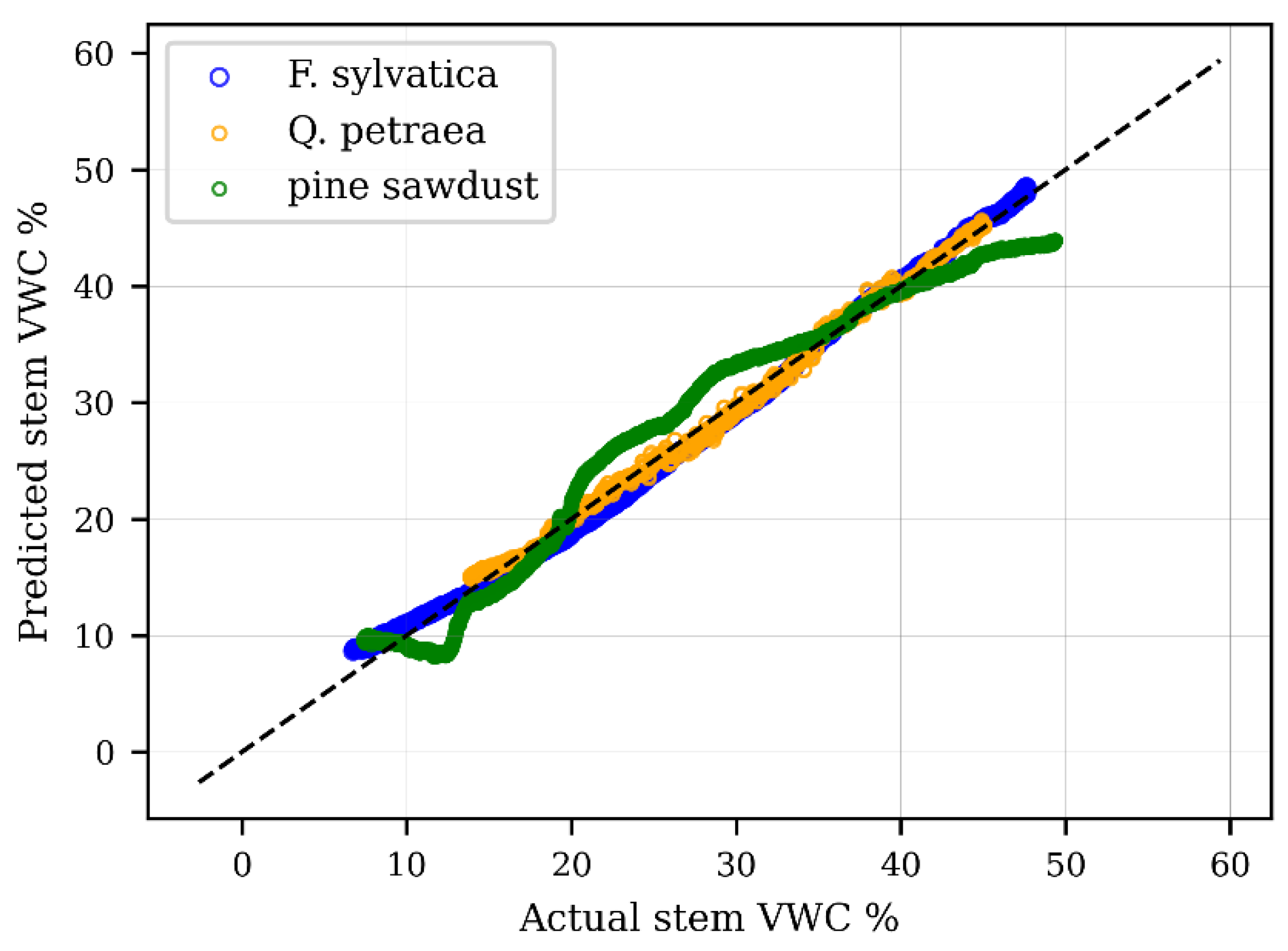
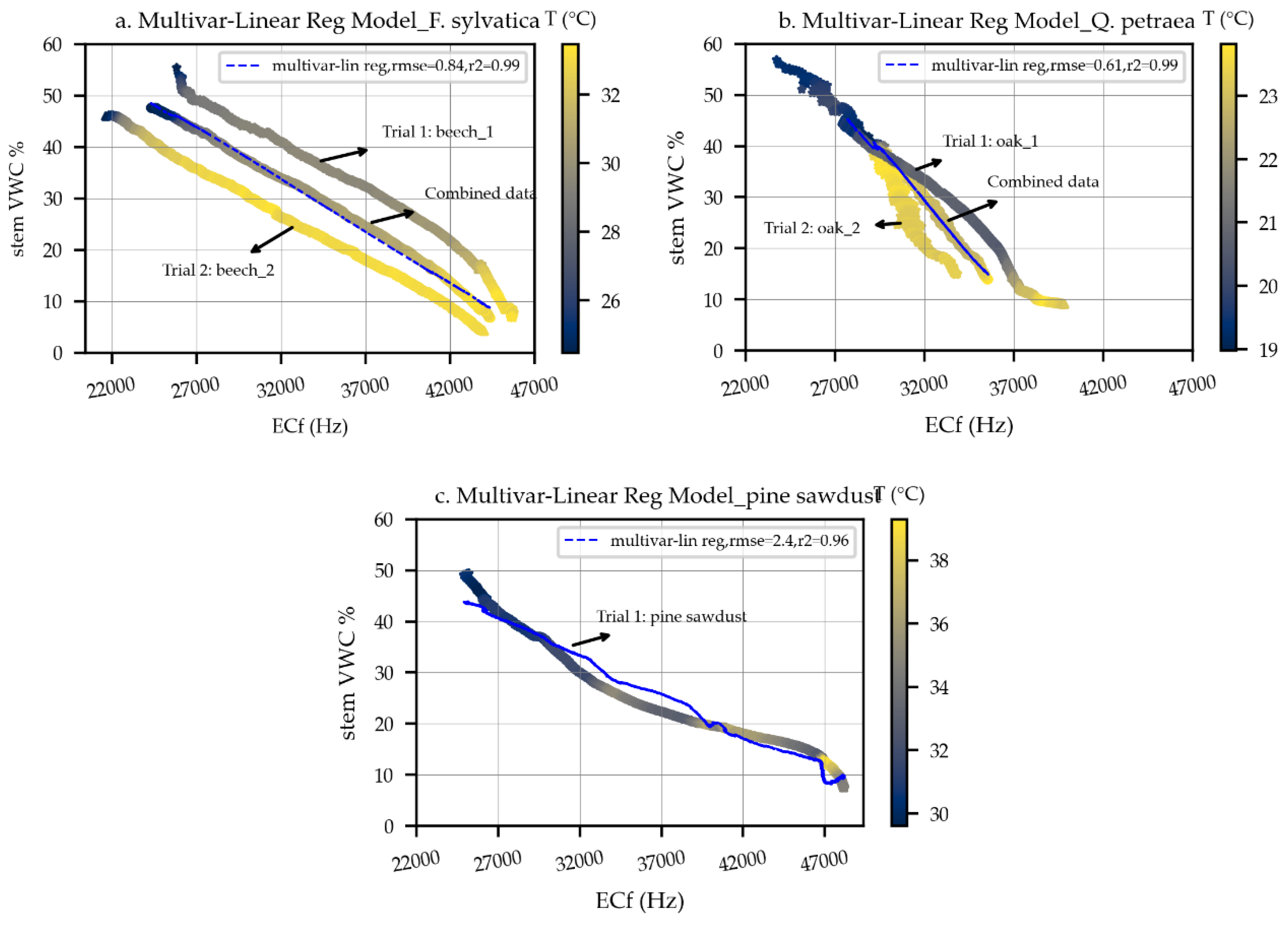
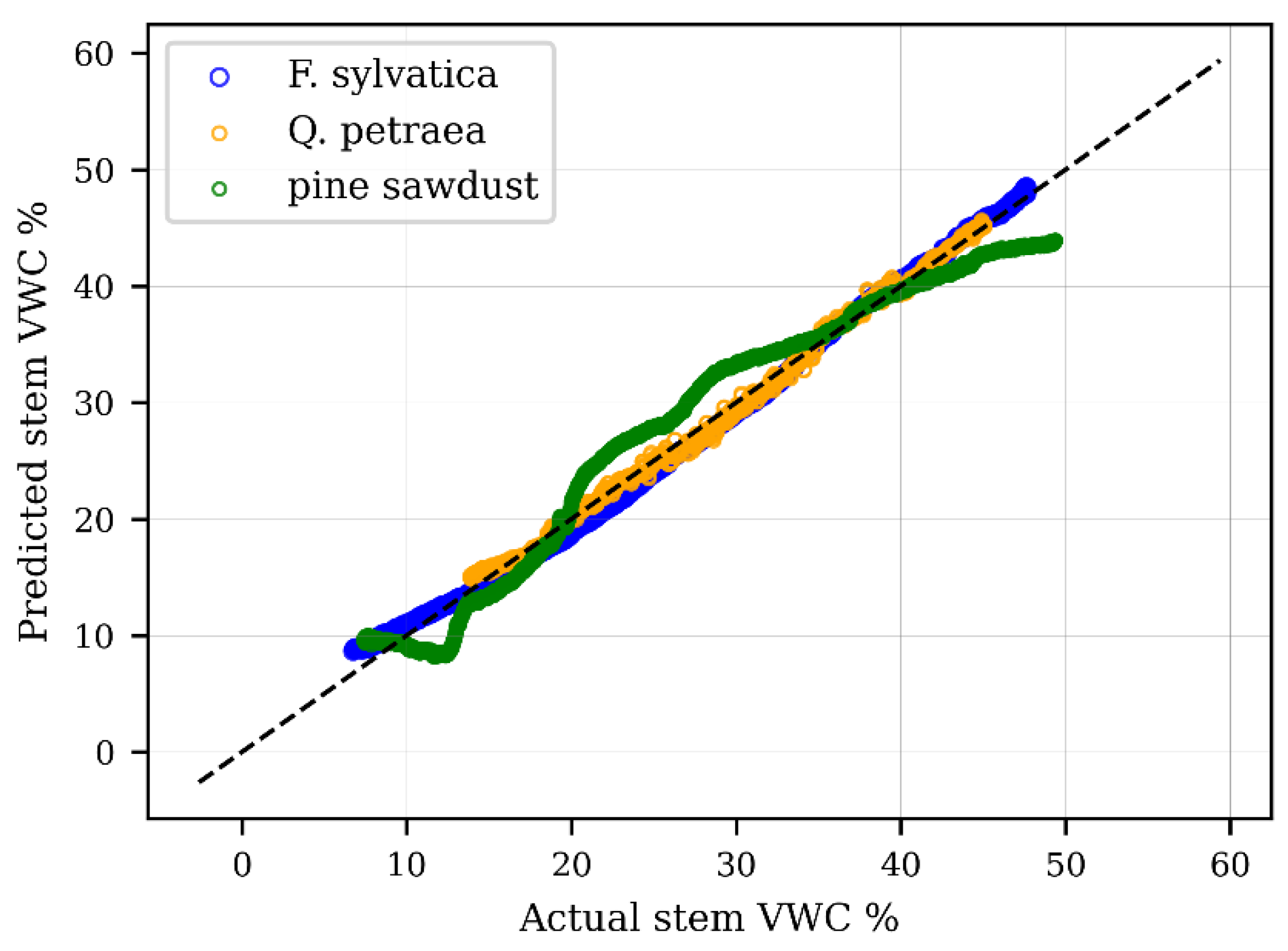
3.1.2. Species-Specific Calibration
3.1.3. Relative Saturation Index
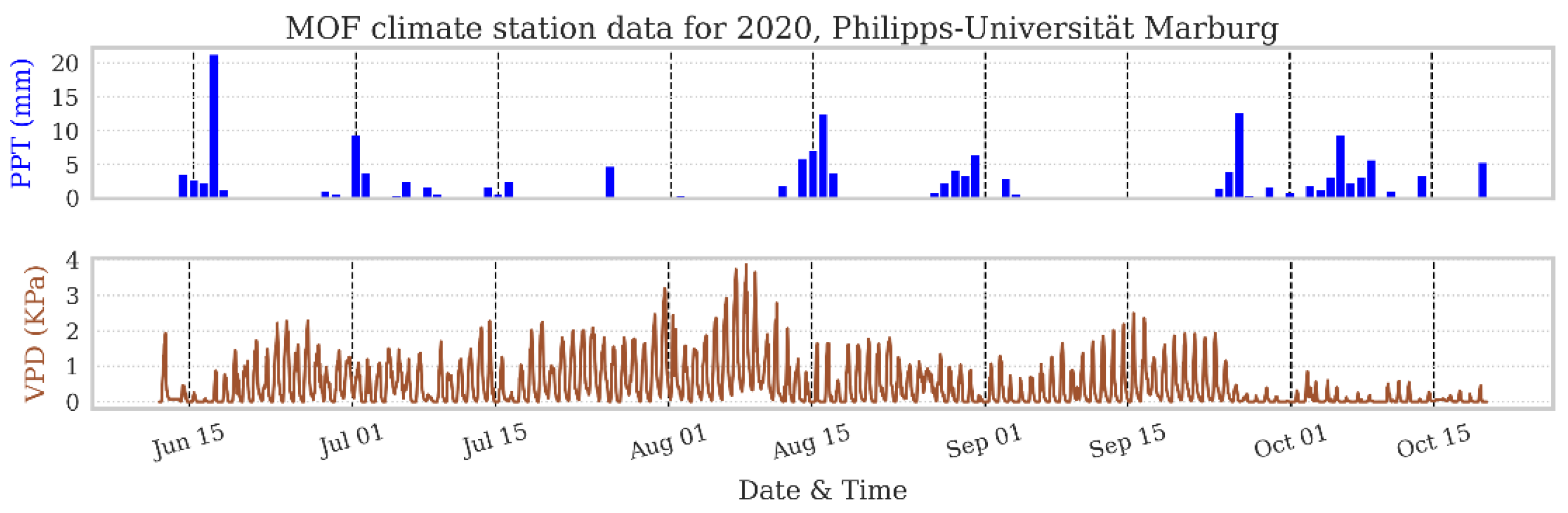
3.2. Field Results
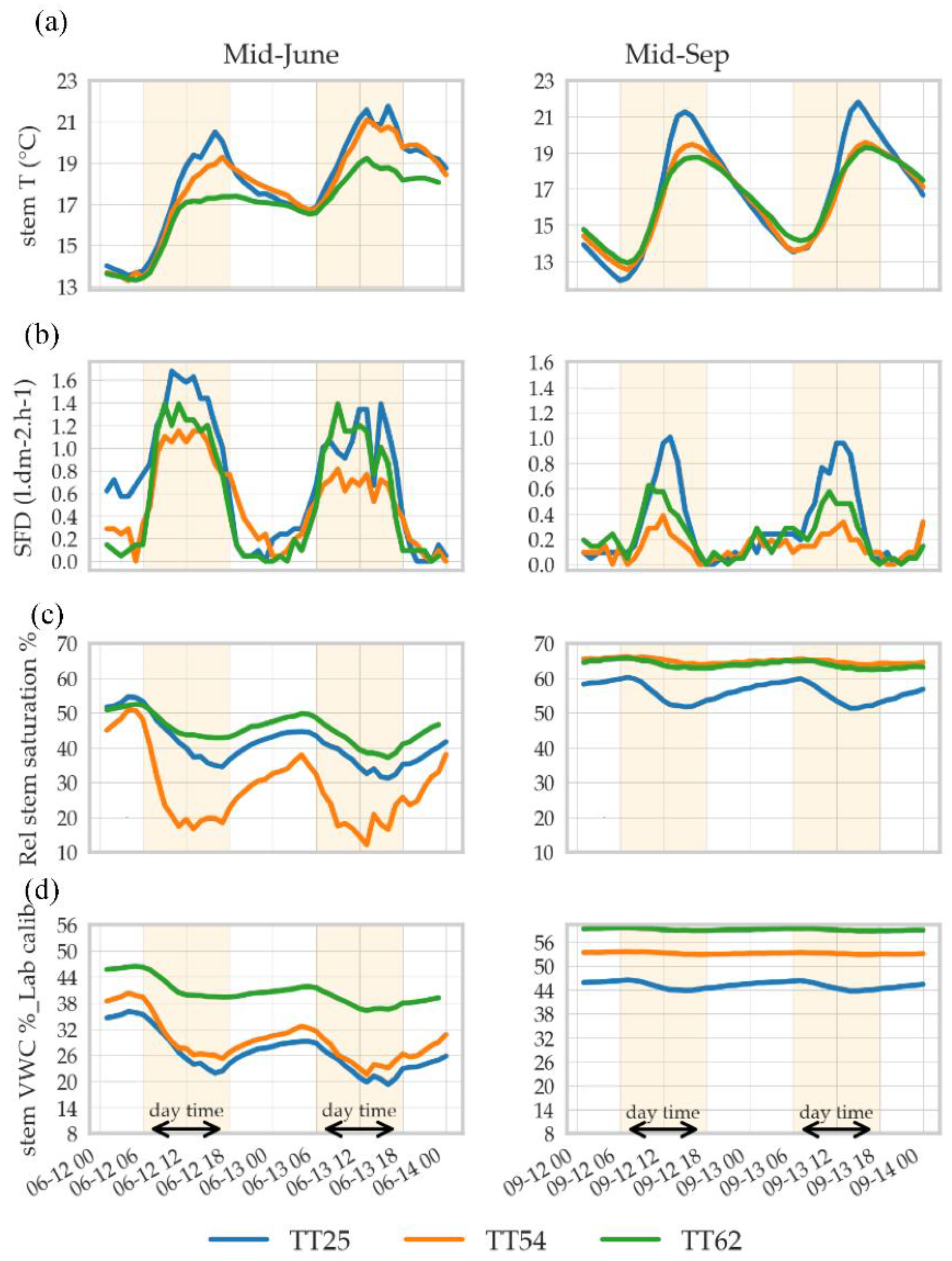
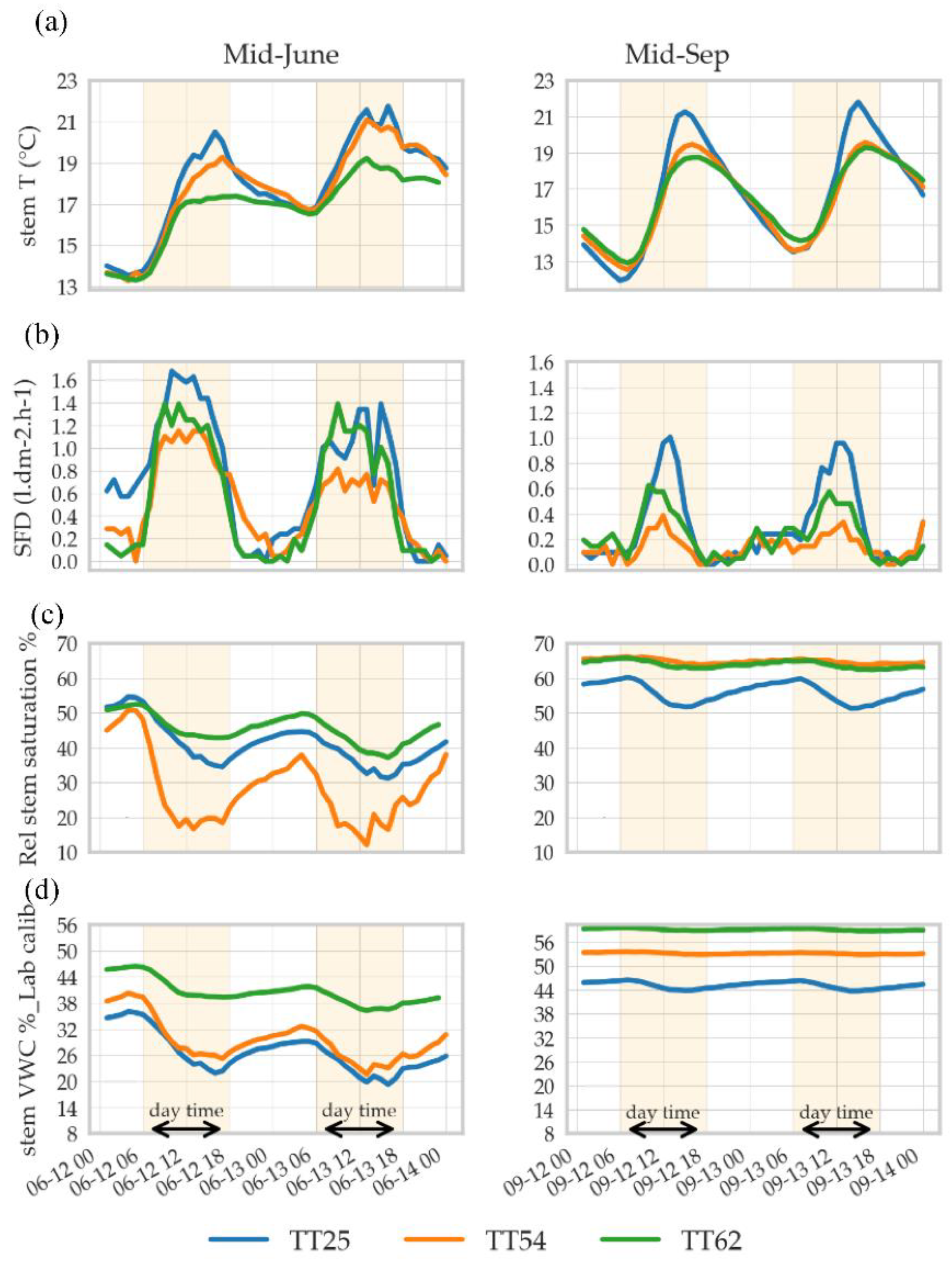
3.2.1. Individual Variability in Sap Flow Rate and Stem VWC in Beech
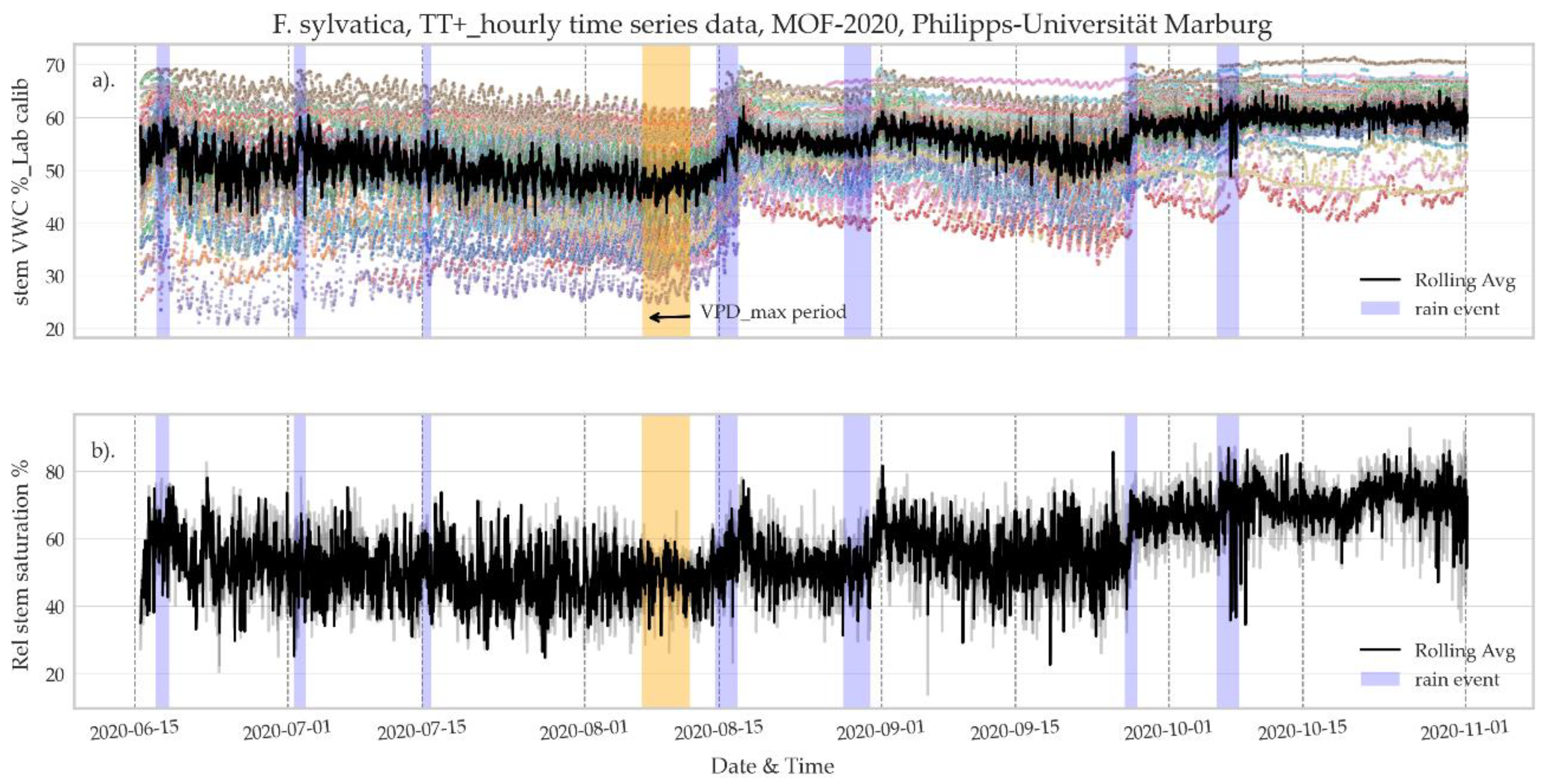
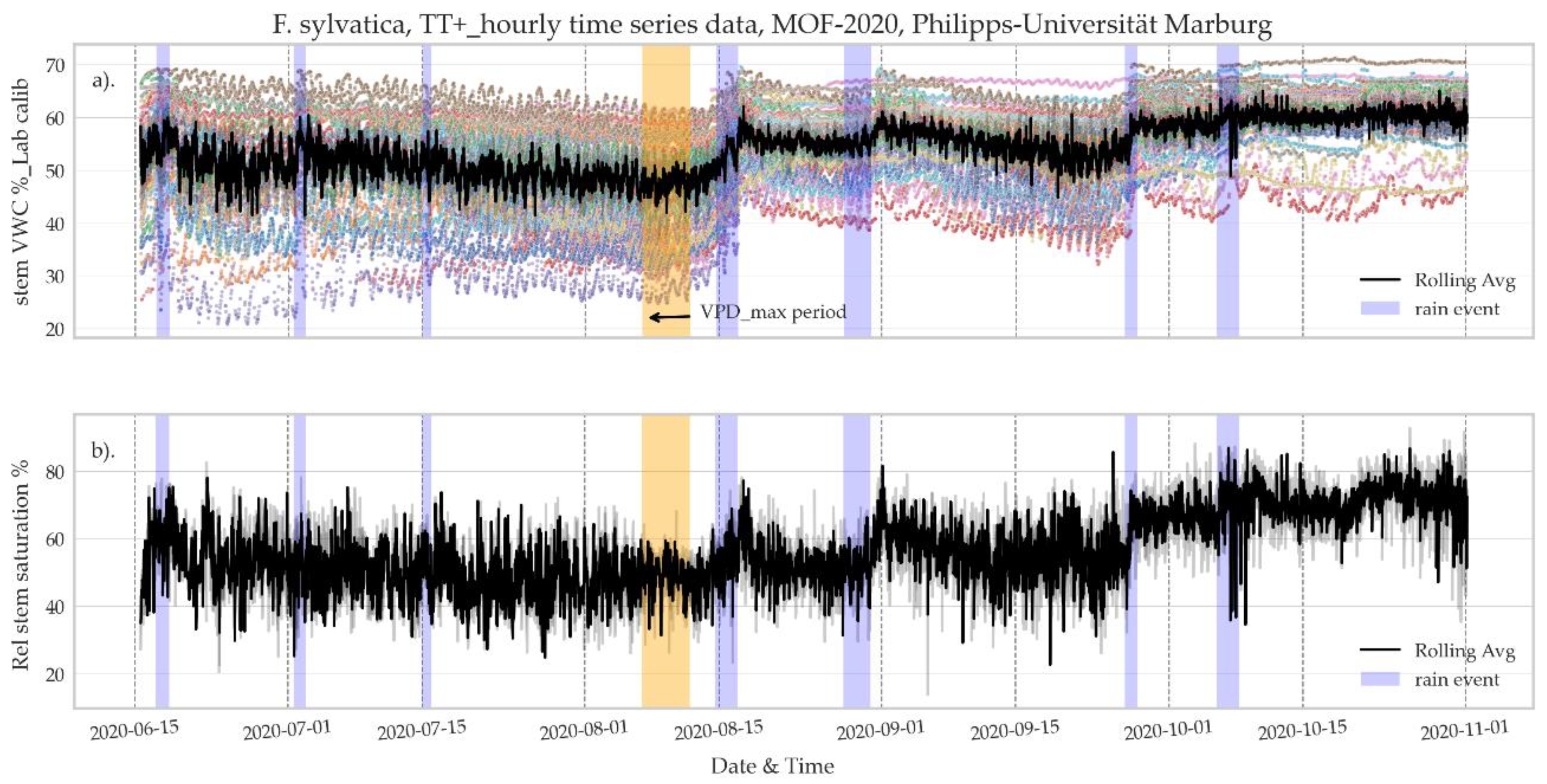
3.2.2. Seasonal Rhythm of Stem VWC and Relative Saturation Index
4. Conclusions
- The TreeTalker, having a capacitive sensor, is an effective IoT-based device for real-time and continuous estimation of relative stem saturation at a single tree level, furthermore, demonstrating a reasonable capacity for capturing a single tree’s ecophysiological behavior under differing environmental conditions.
- The application of continuous and simultaneous monitoring of several trees is showing a remarkable coherent response to climate events (rain and drought), where precipitation has a positive impact, with increased stem VWC, and decreases are evident in dry periods.
- Stem water content varies regularly according to diurnal and nocturnal fluctuation, following the overall trend of rising during the night and falling during the day. This observed trend is coherent with stem recharge and increased transpiration rates are driven by photosynthesis.
- The accuracy of the sensor, however, seems not yet to be adequate for diurnal trend estimation under small water content variations. We are continuously working on improving the sensor with a high frequency of oscillation (100–500 MHz) to increase the sensitivity to the level requested by daily trends.
- Individual tree ecophysiological and geometrical characteristics demonstrate a plausible connection to the pattern and amount of stem water content.
- For further optimization of species-specific calibrations of the Ref probe, we suggest a seasonal sampling campaign to collect wood core samples.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sevanto, S.; Mcdowell, N.G.; Dickman, L.T.; Pangle, R.; Pockman, W.T. How do trees die? A test of the hydraulic failure and carbon starvation hypotheses. Plant Cell Environ. 2014, 37, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Nadezhdina, N. Sap flow index as an indicator of plant water status. Tree Physiol. 1999, 19, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Salomón, R.L.; Steppe, K.; Ourcival, J.M.; Villers, S.; Rodríguez-Calcerrada, J.; Schapman, R.; Limousin, J.M. Hydraulic acclimation in a Mediterranean oak subjected to permanent throughfall exclusion results in increased stem hydraulic capacitance. Plant Cell Environ. 2020, 43, 1528–1544. [Google Scholar] [CrossRef]
- Waring, R.H.; Running, S.W. Sapwood water storage: Its contribution to transpiration and effect upon water conductance through the stems of old-growth Douglas-fir. Plant Cell Environ. 1978, 1, 131–140. [Google Scholar] [CrossRef]
- Čermák, J.; Kučera, J.; Bauerle, W.L.; Phillips, N.; Hinckley, T.M. Tree water storage and its diurnal dynamics related to sap flow and changes in stem volume in old-growth Douglas-fir trees. Tree Physiol. 2007, 27, 181–198. [Google Scholar] [CrossRef]
- Dzikiti, S.; Steppe, K.; Lemeur, R.; Milford, J.R. Whole-tree level water balance and its implications on stomatal oscillations in orange trees [Citrus sinensis (L.) Osbeck] under natural climatic conditions. J. Exp. Bot. 2007, 58, 1893–1901. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, G.; Andrade, J.L.; Meinzer, F.C.; Holbrook, N.M.; Cavelier, J.; Jackson, P.; Celis, A. Stem water storage and diurnal patterns of water use in tropical forest canopy trees. Plant Cell Environ. 1998, 21, 397–406. [Google Scholar] [CrossRef]
- Hernández-Santana, V.; Martínez-Fernández, J.; Morán, C.; Cano, A. Response of Quercus pyrenaica (melojo oak) to soil water deficit: A case study in Spain. Eur. J. For. Res. 2008, 127, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Malavasi, U.C.; Davis, A.S.; de Matos Malavasi, M. Estimativa de água em caule lenhosos vivos—Uma revisão. Cerne 2016, 22, 415–422. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.G. Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef] [Green Version]
- Nadler, A.; Tyree, M.T. Substituting stem’s water content by electrical conductivity for monitoring water status changes. Soil Sci. Soc. Am. J. 2008, 72, 1006–1013. [Google Scholar] [CrossRef]
- López-Bernal, Á.; Testi, L.; Villalobos, F.J. Using the compensated heat pulse method to monitor trends in stem water content in standing trees. Tree Physiol. 2012, 32, 1420–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matheny, A.M.; Bohrer, G.; Garrity, S.R.; Morin, T.H.; Howard, C.J.; Vogel, C.S. Observations of stem water storage in trees of opposing Hydraulic strategies. Ecosphere 2015, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wullschleger, S.D.; Hanson, P.J.; Todd, D.E. Measuring stem water content in four deciduous hardwoods with a time-domain reflectometer. Tree Physiol. 1996, 16, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Constantz, J.; Murphy, F. Monitoring moisture storage in trees using time domain reflectometry. J. Hydrol. 1990, 119, 31–42. [Google Scholar] [CrossRef]
- Irvine, J.; Grace, J. Non-destructive measurement of stem water content by time domain reflectometry using short probes. J. Exp. Bot. 1997, 48, 813–818. [Google Scholar] [CrossRef]
- Boone, R.S.S.; Wengert, E.M.M. Guide for Using the Oven-Dry Method for Determining the Moisture Content of Wood. For. Facts 1998, 89, 1–4. [Google Scholar]
- Edwards, W.R.N.; Jarvis, P.G. A method for measuring radial differences in water content of intact tree stems by attenuation of gamma radiation. Plant Cell Environ. 1983, 6, 255–260. [Google Scholar] [CrossRef]
- Byrne, G.F.; Fenn, M.D.; Burgar, M.I. Nuclear magnetic resonance studies of water in tree sections. Agric. For. Meteorol. 1986, 38, 307–317. [Google Scholar] [CrossRef]
- van As, H.; Scheenen, T.; Vergeldt, F.J. MRI of intact plants. Photosynth. Res. 2009, 102, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Lane, P.N.J.; Mackenzie, D.H.; Nadler, A.D. Note of clarification about: Field and laboratory calibration and test of TDR and capacitance techniques for indirect measurement of soil water content, by P.N.J. Lane and D.H. Mackenzie, Vol. 39, pp. 1371–1386. Soil Res. 2002, 40, 555. [Google Scholar] [CrossRef]
- Nadler, A.; Raveh, E.; Yermiyahu, U.; Green, S.R. Evaluation of TDR use to monitor water content in stem of lemon trees and soil and their response to water stress. Soil Sci. Soc. Am. J. 2003, 67, 437–448. [Google Scholar] [CrossRef]
- Nadler, A.; Raveh, E.; Yermiyahu, U.; Green, S. Stress Induced Water Content Variations in Mango Stem by Time Domain Reflectometry. Soil Sci. Soc. Am. J. 2006, 70, 510–520. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Turner, N.C.; Aogu, K.; Dyck, M.; Feng, H.; Si, B.; Wang, J.; Lv, J. Time and frequency domain reflectometry for the measurement of tree stem water content: A review, evaluation, and future perspectives. Agric. For. Meteorol. 2021, 306, 108442. [Google Scholar] [CrossRef]
- Vandegehuchte, M.W.; Steppe, K. Sapflow+: A four-needle heat-pulse sap flow sensor enabling nonempirical sap flux density and water content measurements. New Phytol. 2012, 196, 306–317. [Google Scholar] [CrossRef] [PubMed]
- Vergeynst, L.L.; Vandegehuchte, M.W.; McGuire, M.A.; Teskey, R.O.; Steppe, K. Changes in stem water content influence sap flux density measurements with thermal dissipation probes. Trees 2014, 28, 949–955. [Google Scholar] [CrossRef]
- Dietsch, P.; Franke, S.; Franke, B.; Gamper, A.; Winter, S. Methods to determine wood moisture content and their applicability in monitoring concepts. J. Civ. Struct. Health Monit. 2015, 5, 115–127. [Google Scholar] [CrossRef]
- Hardie, M. Review of Novel and Emerging Proximal Soil Moisture Sensors for Use in Agriculture. Sensors 2020, 20, 6934. [Google Scholar] [CrossRef]
- Stott, L.V.; Black, B.; Bugbee, B. Quantifying Tree Hydration Using Electromagnetic Sensors. Horticulturae 2020, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh, A.; Toudeshki, A.; Ehsani, R.; Migliaccio, K.; Wang, D. Detecting tree water stress using a trunk relative water content measurement sensor. Smart Agric. Technol. 2021, 1, 100003. [Google Scholar] [CrossRef]
- Looker, N.; Martin, J.; Jencso, K.; Hu, J. Contribution of sapwood traits to uncertainty in conifer sap flow as estimated with the heat-ratio method. Agric. For. Meteorol. 2016, 223, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Chetpattananondh, P.; Thongpull, K.; Chetpattananondh, K. Interdigital capacitance sensing of moisture content in rubber wood. Comput. Electron. Agric. 2017, 142, 545–551. [Google Scholar] [CrossRef]
- Fares, A.; Polyakov, V. Advances in Crop Water Management Using Capacitive Water Sensors. Adv. Agron. 2006, 90, 43–77. [Google Scholar] [CrossRef]
- Fuchs, A.; Moser, M.J.; Zangl, H.; Bretterklieber, T. Using Capacitive Sensing To Determine the Moisture Content of Wood Pellets—Investigations and Application. Int. J. Smart Sens. Intell. Syst. 2009, 2, 293–308. [Google Scholar] [CrossRef] [Green Version]
- Korkua, S.K.; Sakphrom, S. Low-cost capacitive sensor for detecting palm-wood moisture content in real-time. Heliyon 2020, 6, e04555. [Google Scholar] [CrossRef]
- Matheny, A.M.; Garrity, S.R.; Bohrer, G. The calibration and use of capacitance sensors to monitor stem water content in trees. J. Vis. Exp. 2017, 2017, 1–10. [Google Scholar] [CrossRef]
- Liang, H.; Zhang, M.; Wang, H.; Gao, C.; Zhao, Y. Variation Characteristics of Stem Water Content in Lagerstroemia indica and Its Response to Environmental Factors. J. Sens. 2020, 2020, 8689272. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Sun, Y.; Tyree, M.T.; Sheng, W.; Cheng, Q.; Xue, X.; Schumann, H.; Schulze Lammers, P. An improved sensor for precision detection of in situ stem water content using a frequency domain fringing capacitor. New Phytol. 2015, 206, 471–481. [Google Scholar] [CrossRef]
- Hao, G.Y.; Wheeler, J.K.; Holbrook, N.M.; Goldstein, G. Investigating xylem embolism formation, refilling and water storage in tree trunks using frequency domain reflectometry. J. Exp. Bot. 2013, 64, 2321–2332. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Yan, X.; Grantz, D.A.; Xiang, Y.; de Oliveira, R.F.; Huang, L.; Wang, Z.; Du, T.; Cheng, Q. In-situ and non-invasive measurement of stem water content of trees using an innovative interdigitated-electrodes dielectric sensor less susceptible to stem diameter variation. Agric. For. Meteorol. 2021, 307, 108473. [Google Scholar] [CrossRef]
- Saito, T.; Yasuda, H.; Sakurai, M.; Acharya, K.; Sueki, S.; Inosako, K.; Yoda, K.; Fujimaki, H.; Abd Elbasit, M.A.M.; Eldoma, A.M.; et al. Monitoring of Stem Water Content of Native and Invasive Trees in Arid Environments Using GS3 Soil Moisture Sensors. Vadose Zone J. 2016, 15, vzj2015.04.0061. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Sun, Y.; Shan, G.; Grantz, D.A.; Cheng, Q.; Schulze Lammers, P.; Damerow, L.; Wen, B.; Xue, X.; Chen, B. In situ measurement of stem water content and diurnal storage of an apricot tree with a high frequency inner fringing dielectric sensor. Agric. For. Meteorol. 2018, 250–251, 35–46. [Google Scholar] [CrossRef]
- Araújo, G.P.; Vellame, L.M.; Costa, J.A.; Costa, C.A.G. A low-cost monitoring system of stem water content: Development and application to Brazilian forest species. Smart Agric. Technol. 2021, 1, 100012. [Google Scholar] [CrossRef]
- Ren, R.; von der Crone, J.; Horton, R.; Liu, G.; Steppe, K. An improved single probe method for sap flow measurements using finite heating duration. Agric. For. Meteorol. 2020, 280, 107788. [Google Scholar] [CrossRef]
- Windt, C.W.; Blümler, P. A portable NMR sensor to measure dynamic changes in the amount of water in living stems or fruit and its potential to measure sap flow. Tree Physiol. 2015, 35, 366–375. [Google Scholar] [CrossRef] [Green Version]
- Matasov, V.; Marchesini, L.B.; Yaroslavtsev, A.; Sala, G.; Fareeva, O.; Seregin, I.; Castaldi, S.; Vasenev, V.; Valentini, R. IoT monitoring of urban tree ecosystem services: Possibilities and challenges. Forests 2020, 11, 775. [Google Scholar] [CrossRef]
- Valentini, R.; Marchesini, L.B.; Gianelle, D.; Sala, G.; Yarovslavtsev, A.; Vasenev, V.I.; Castaldi, S. New tree monitoring systems: From industry 4.0 to nature 4. Ann. Silvic. Res. 2019, 43, 84–88. [Google Scholar] [CrossRef]
- Zorzi, I.; Francini, S.; Chirici, G.; Cocozza, C. The TreeTalkersCheck R package: An automatic daily routine to check physiological traits of trees in the forest. Ecol. Inform. 2021, 66, 101433. [Google Scholar] [CrossRef]
- Granier, A. Une nouvelle méthode pour la mesure du flux de sève brute dans le tronc des arbres. Ann. Sci. For. 1985, 42, 193–200. [Google Scholar] [CrossRef]
- Nhean, S.; Isarangkool Na Ayutthaya, S.; Rocheteau, A.; Do, F.C.; Tognetti, R. Multi-species test and calibration of an improved transient thermal dissipation system of sap flow measurement with a single probe. Tree Physiol. 2019, 39, 1061–1070. [Google Scholar] [CrossRef]
- Masmoudi, M.M.; Mahjoub, I.; Lhomme, J.P.; Ben Mechlia, N. Sap flow measurement by a single thermal dissipation probe in transient regime: Implementation of the method and test under field conditions. Ann. For. Sci. 2012, 69, 773–781. [Google Scholar] [CrossRef]
- Isarangkool Na Ayutthaya, S.; Do, F.C.; Pannengpetch, K.; Junjittakarn, J.; Maeght, J.L.; Rocheteau, A.; Cochard, H. Transient thermal dissipation method of xylem sap flow measurement: Multi-species calibration and field evaluation. Tree Physiol. 2010, 30, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Do, F.C.; Isarangkool Na Ayutthaya, S.; Rocheteau, A. Transient thermal dissipation method for xylem sap flow measurement: Implementation with a single probe. Tree Physiol. 2011, 31, 369–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, F.C.; Puangjumpa, N.; Rocheteau, A.; Duthoit, M.; Nhean, S.; Isarangkool Na Ayutthaya, S. Towards reduced heating duration in the transient thermal dissipation system of sap flow measurements. Acta Hortic. 2018, 1222, 229–235. [Google Scholar] [CrossRef]
- Whalley, W.R.; Dean, T.J.; Izzard, P. Evaluation of the capacitance technique as a method for dynamically measuring soil water content. J. Agric. Eng. Res. 1992, 52, 147–155. [Google Scholar] [CrossRef]
- Zhu, Y.; Irmak, S.; Jhala, A.J.; Vuran, M.C.; Diotto, A. Time-domain and frequency-domain reflectometry type soil moisture sensor performance and soil temperature effects in fine- And coarse-textured soils. Appl. Eng. Agric. 2019, 35, 117–134. [Google Scholar] [CrossRef]
- Dean, T.J.; Bell, J.P.; Baty, A.J.B. Soil moisture measurement by an improved capacitance technique, Part I. Sensor design and performance. J. Hydrol. 1987, 93, 67–78. [Google Scholar] [CrossRef]
- Kizito, F.; Campbell, C.S.; Campbell, G.S.; Cobos, D.R.; Teare, B.L.; Carter, B.; Hopmans, J.W. Frequency, electrical conductivity and temperature analysis of a low-cost capacitance soil moisture sensor. J. Hydrol. 2008, 352, 367–378. [Google Scholar] [CrossRef]
- Fares, A.; Safeeq, M.; Awal, R.; Fares, S.; Dogan, A. Temperature and Probe-to-Probe Variability Effects on the Performance of Capacitance Soil Moisture Sensors in an Oxisol. Vadose Zone J. 2016, 15, vzj2015.07.0098. [Google Scholar] [CrossRef] [Green Version]
- Friess, N.; Bendix, J.; Brändle, M.; Brandl, R.; Dahlke, S.; Farwig, N.; Freisleben, B.; Holzmann, H.; Meyer, H.; Müller, T.; et al. Introducing Nature 4.0: A sensor network for environmental monitoring in the Marburg Open Forest. Biodivers. Inf. Sci. Stand. 2019, 3, 3–4. [Google Scholar] [CrossRef]
- Wiedemann, A.; Marañón-Jiménez, S.; Rebmann, C.; Herbst, M.; Cuntz, M. An empirical study of the wound effect on sap flux density measured with thermal dissipation probes. Tree Physiol. 2016, 36, 1471–1484. [Google Scholar] [CrossRef] [PubMed]
- Wolf, H. Determination of water density: Limitations at the uncertainty level of 1 × 10−6. Accredit. Qual. Assur. 2008, 13, 587–591. [Google Scholar] [CrossRef]
- Neuwirth, B.; Rabbel, I.; Bendix, J.; Bogena, H.R.; Thies, B. The european heat wave 2018: The dendroecological response of oak and spruce in western germany. Forests 2021, 12, 283. [Google Scholar] [CrossRef]
- Schermelleh-Engel, K.; Moosbrugger, H.; Müller, H. Evaluating the Fit of Structural Equation Models: Tests of Significance and Descriptive Goodness-of-Fit Measures. Methods Psychol. Res. Online 2003, 8, 23–74. [Google Scholar]
- Nola, P.; Bracco, F.; Assini, S.; Von Arx, G.; Castagneri, D. Xylem anatomy of Robinia pseudoacacia L. and Quercus robur L. is differently affected by climate in a temperate alluvial forest. Ann. For. Sci. 2020, 77, 1–16. [Google Scholar] [CrossRef]
- Bowman, W.P.P.; Barbour, M.M.M.; Turnbull, M.H.H.; Tissue, D.T.T.; Whitehead, D.; Griffin, K.L.L. Sap flow rates and sapwood density are critical factors in within- and between-tree variation in CO2 efflux from stems of mature Dacrydium cupressinum trees. New Phytol. 2005, 167, 815–828. [Google Scholar] [CrossRef]
- Barbour, M.M.; Whitehead, D. A demonstration of the theoretical prediction that sap velocity is related to wood density in the conifer Dacrydium cupressinum. New Phytol. 2003, 158, 477–488. [Google Scholar] [CrossRef] [Green Version]
- Roderick, M.L.; Berry, S.L. Linking wood density with tree growth and environment: A theoretical analysis based on the motion of water. New Phytol. 2001, 149, 473–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalevanková, P.; Sitková, Z.; Kučera, J.; Střelcová, K. Impact of water deficit on seasonal and diurnal dynamics of european beech transpiration and time-lag effect between stand transpiration and environmental drivers. Water 2020, 12, 3437. [Google Scholar] [CrossRef]
- Gebauer, T.; Horna, V.; Leuschner, C. Canopy transpiration of pure and mixed forest stands with variable abundance of European beech. J. Hydrol. 2012, 442–443, 2–14. [Google Scholar] [CrossRef]
- Hafner, B.D.; Tomasella, M.; Häberle, K.H.; Goebel, M.; Matyssek, R.; Grams, T.E.E. Hydraulic redistribution under moderate drought among English oak, European beech and Norway spruce determined by deuterium isotope labeling in a split-root experiment. Tree Physiol. 2017, 37, 950–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TT+ No. | Height (m) | DBH (cm) | Sapwood Area (cm2) | Canopy Volume (m3) |
|---|---|---|---|---|
| TT25 | 29.9 | 53 | 685 | 879 |
| TT54 | 32.7 | 56 | 806 | 1017 |
| TT62 | 29.5 | 53 | 615 | 1801 |
| Var | Water | Sawdust | F. sylvatica | P. nigra | P. sylvestris |
|---|---|---|---|---|---|
| Slope () | −81 | −87 | −43 | −84 | −78 |
| Intercept () | 13,867 | 18,519 | 21,834 | 33,283 | 27,555 |
| R2 | 0.98 | 0.88 | 0.94 | 0.98 | 0.81 |
| p-value | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| stderr | 0.53 | 0.92 | 0.45 | 0.41 | 1.28 |
| Scheme 2 | R2 | p > |t| | RMSE | No. Obs | |||
|---|---|---|---|---|---|---|---|
| Beech | −0.23 | −0.0021 | 93 | 0.996 | <0.001 | 0.86 | 1430 |
| Oak | −0.56 | −0.0042 | 150 | 0.996 | <0.001 | 0.61 | 889 |
| Pine sawdust | −0.61 | −0.0013 | 96 | 0.969 | <0.001 | 2.4 | 8263 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asgharinia, S.; Leberecht, M.; Belelli Marchesini, L.; Friess, N.; Gianelle, D.; Nauss, T.; Opgenoorth, L.; Yates, J.; Valentini, R. Towards Continuous Stem Water Content and Sap Flux Density Monitoring: IoT-Based Solution for Detecting Changes in Stem Water Dynamics. Forests 2022, 13, 1040. https://doi.org/10.3390/f13071040
Asgharinia S, Leberecht M, Belelli Marchesini L, Friess N, Gianelle D, Nauss T, Opgenoorth L, Yates J, Valentini R. Towards Continuous Stem Water Content and Sap Flux Density Monitoring: IoT-Based Solution for Detecting Changes in Stem Water Dynamics. Forests. 2022; 13(7):1040. https://doi.org/10.3390/f13071040
Chicago/Turabian StyleAsgharinia, Shahla, Martin Leberecht, Luca Belelli Marchesini, Nicolas Friess, Damiano Gianelle, Thomas Nauss, Lars Opgenoorth, Jim Yates, and Riccardo Valentini. 2022. "Towards Continuous Stem Water Content and Sap Flux Density Monitoring: IoT-Based Solution for Detecting Changes in Stem Water Dynamics" Forests 13, no. 7: 1040. https://doi.org/10.3390/f13071040
APA StyleAsgharinia, S., Leberecht, M., Belelli Marchesini, L., Friess, N., Gianelle, D., Nauss, T., Opgenoorth, L., Yates, J., & Valentini, R. (2022). Towards Continuous Stem Water Content and Sap Flux Density Monitoring: IoT-Based Solution for Detecting Changes in Stem Water Dynamics. Forests, 13(7), 1040. https://doi.org/10.3390/f13071040