Abstract
Environments are ubiquitously heterogeneous in nature, and clonal plants commonly benefit from both clonal integration and foraging responses in heterogeneous environments. While many studies have examined clonal integration and foraging responses separately, few have tested the effect of clonal integration on the foraging response of clonal plants to environmental heterogeneity. We grew offspring ramets of each of three clonal plants (Hydrocotyle vulgaris, Duchesnea indica, and Glechoma longituba) in both homogeneous and heterogenous soil environments and severed their stem connection to a mother ramet (to prevent clonal integration from the mother ramet) or kept it intact (to allow clonal integration). Without clonal integration from the mother ramet, soil heterogeneity had no effect on biomass or number of ramets for any of the three species. With clonal integration, soil heterogeneity also had no effect on biomass or number of ramets of D. indica and G. longituba, but significantly decreased biomass and marginally significantly decreased number of ramets of H. vulgaris. Without clonal integration, offspring ramets did not demonstrate either shoot or root foraging responses in terms of total, shoot and root biomass and ramet number in the heterogeneous soil environment in any of the three species. With integration, offspring ramets of H. vulgaris also did not demonstrate either root or shoot foraging responses, but offspring ramets of G. longituba demonstrated both root and shoot foraging responses, and those of D. indica demonstrated a root foraging response when they grew in the heterogeneous soil environment. We conclude that clonal integration can alter the foraging response of clonal plants, but this effect is species-specific. Our results also suggest that foraging responses of clonal plants in heterogeneous soil environments may not necessarily benefit the growth of clonal plants.
1. Introduction
Clonal plants, such as those capable of clonal growth or vegetive propagation, are abundant in nature, and play a substantial role in diverse habitats [1,2,3,4]. Vegetatively produced offspring ramets commonly remain physically connected to one another and to their mother ramets via horizontally growing roots or stems (stolons, creeping stems, and rhizomes) [5,6,7,8,9,10]. These physical connections allow clonal integration, i.e., translocation of resources (carbohydrates, water, and minimal nutrients) and signals from mother (or developmentally older) ramets to offspring (or developmentally younger) ramets, and from ramets growing in high-quality patches to those growing in low-quality ones in heterogeneous environments [11,12,13,14,15,16]. A large body of studies have suggested that clonal integration can promote the establishment of newly produced offspring ramets, ramets growing in low-quality patches, and the whole clone [10,17,18,19,20].
Environments are ubiquitously heterogeneous, and such environmental heterogeneity can occur at different spatial scales relevant to plant growth, development, and reproduction [21,22,23,24,25]. When clonal plants grow in heterogeneous environments consisting of high- and low-resource patches, they may demonstrate foraging responses by placing more roots (root foraging), shoots (shoot foraging), and/or whole ramets (both root and shoot foraging) in high-resource patches than in low-resource ones [26,27,28,29,30]. Because of such foraging responses, clonal plants can grow better in heterogeneous environments than in homogenous ones even if the total amount of resources (i.e., light, water, and nutrients) is the same in the two environments [10,31,32,33,34,35]. Due to interspecific differences in foraging responses, environmental heterogeneity may potentially influence competitive interactions between plants [36,37,38,39,40] and alter species composition and community productivity [25,41,42,43,44,45,46].
While numerous studies have separately tested the roles of clonal integration [16,20,47,48,49,50,51,52,53] and foraging responses of clonal plants in heterogeneous environments [26,35,54,55], to our best knowledge, few studies have assessed them simultaneously. As both clonal integration and foraging responses can potentially promote the growth of clonal plants [7,13,19,20,31,35,38], we hypothesize that the foraging responses of offspring ramets of clonal plants in heterogeneous environments will be more significant when clonal integration from their connected mother ramets is prevented than when it is allowed, i.e., clonal integration will alter the foraging responses of offspring ramets of clonal plants growing in heterogeneous environments. Since foraging responses are frequently associated with growth promotion [35,56,57,58,59], we hypothesize further that the positive effect of environmental heterogeneity on the growth of offspring ramets of clonal plants will be stronger when clonal integration from their connected mother ramets is prevented than when it is allowed, i.e., clonal integration will alter the effect of environmental heterogeneity on the growth of offspring ramets.
To test these hypotheses, we grew offspring ramets of each of three clonal plants (Hydrocotyle vulgaris, Duchesnea indica, and Glechoma longituba) in both homogeneous and heterogenous soil environments and severed their physical (stolon or creeping stem) connection to a mother ramet (to prevent clonal integration from the mother ramet) or kept it intact (to allow clonal integration). The total amount of nutrients and substrates in the homogeneous and heterogeneous environments remained the same. Specifically, we addressed the following questions: (1) Do the three clonal plants demonstrate foraging responses in terms of shoot, root, and total biomass and number of ramets in the heterogeneous soil environment? (2) Do such foraging responses in the heterogeneous soil environment, if any, result in a growth promotion of the offspring ramets compared to the homogeneous soil environment? (3) Does clonal integration from the connected mother ramet affect the foraging responses of the offspring ramets in the heterogeneous soil environment? (4) Does clonal integration alter the effect of soil heterogeneity on the growth of the offspring ramets?
2. Materials and Methods
2.1. The Species
The three study species, i.e., Duchesnea indica (Andr.) Focke (Rosaceae), Glechoma longituba Nakai Kupr. (Lamiaceae), and Hydrocotyle vulgaris L. (Umbelliferae), are all perennial herbs capable of vigorous clonal growth and widely distributed in China [60,61]. All three species can be distributed at forest margins, and G. longituba and D. indica can also occur under forests. Duchesnea indica and G. longituba are stoloniferous plants and H. vulgaris produces creeping stems that can root at each node [62,63,64]. A ramet of D. indica is a rosette, consisting of a vertical stem with highly compressed internodes, some leaves, and roots [62]. Each leaf is composed of a slender petiole with three leaflets and is borne on the vertical stem. The axillary buds on the vertical stem can develop into stolons. A ramet of G. longituba commonly consists of two single leaves, a node and some roots, and the two axillary buds at each stolon node can grow out to form two stolons [63]. A ramet of H. vulgaris consists of only one single leaf, one node, and some roots, and the axillary bud at each node can form a creeping stem [64].
2.2. Sampling and Cultivation
Ramets of all three species were collected at the Jiaojiang campus of Taizhou University in the Jiaojiang District, Taizhou, Zhejiang Province, China. They were vegetatively propagated in a greenhouse at Taizhou University. For each species, 35 similar-sized ramets were selected and planted individually in pots (14 cm in diameter and 13 cm high) filled with an equal mixture of sand and commercial compost (Meishimei Bio-Tech Co. Ltd., Beijing, China). After 30 days, for each species we selected, out of the 35 ramets, 20 which produced at least one stolon or creeping stem with at least one ramet. For each ramet selected, one stolon of similar size was kept and the remaining stolons or creeping stems, if any, were removed.
2.3. Experimental Design
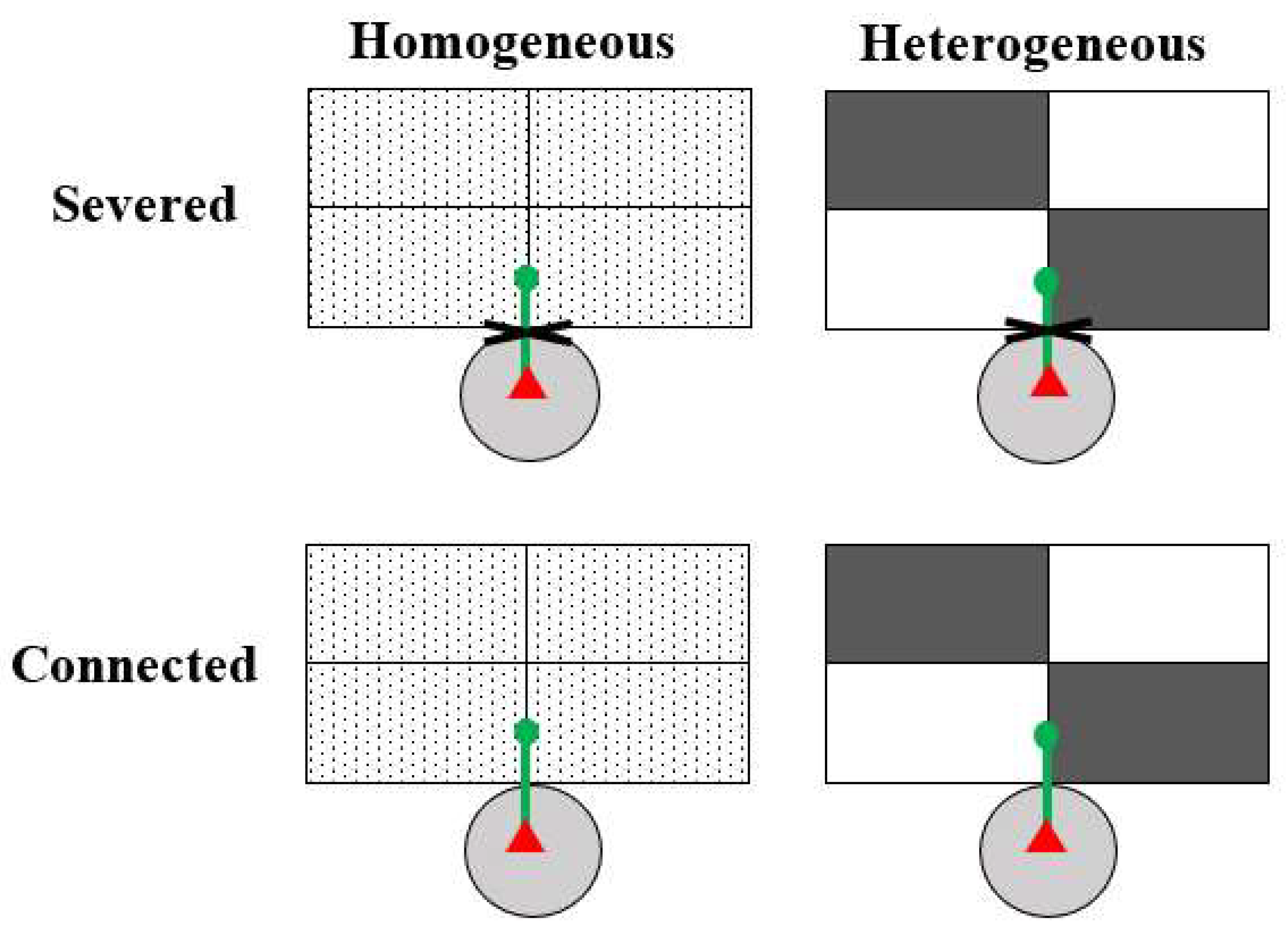
For each species, we set up four treatments, which were two soil heterogeneity treatments (homogeneous vs. heterogeneous) crossed with two clonal integration treatments (connection vs. severance, i.e., with vs. without integration; Figure 1). The 20 ramets of each species were randomly assigned to one of the four treatments, and each treatment was replicated five times. Each box (39 cm long × 28 cm wide × 14 cm deep) was divided into four equal patches (19.5 cm long × 14 cm wide). For the heterogeneous treatment, two patches in each box were filled with a high-nutrient soil and the other two with low-nutrient soil, arranged in a checkboard manner (Figure 1). For the homogeneous treatment, each of the four patches was filled with an even mixture of the high- and low-nutrient soil at a volume ratio of 1:1. Thus, the total amount of soil nutrients in each box was the same for all treatments.
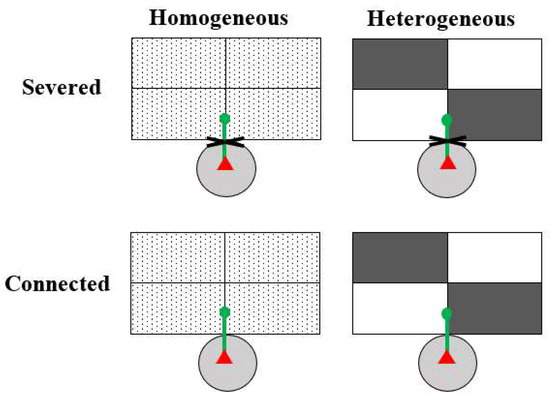
Figure 1.
Schematic representation of the experimental design. Each box consisted of four equal patches. In the heterogeneous treatment, two patches in a box were filled with a high-nutrient soil (gray rectangles) and the other two with a low-nutrient soil (white rectangles); in the homogeneous treatment, all four patches in a box were filled with an even mixture of the high- and the low-nutrient soils at a volume ratio of 1:1. A mother ramet (red triangle) was grown in a pot (light gray circle) and its first daughter ramet (green dot) was grown at the short edge of the two adjacent patches. The internode (thick green lines) between the mother and the first daughter ramet was either severed to prevent clonal integration or kept connected to allow clonal integration.
The high- and low-nutrient soils were a mixture of sand and commercial compost (5.59 g kg−1 total N, 3.99 g kg−1 total P and 146.2 g kg−1 organic matter; Hebei Dewoduo Fertilizer Co., Ltd, Beijing, China) at volume ratios of 1:9 and 9:1 ratio, respectively. As sand commonly contains very low amounts of N, P, and K, the amounts of N, P, and K in the sand–compost mixtures roughly equal to those in compost. Therefore, the total amounts of N, P, and K in the high-nutrient soil were about nine times higher than those in the low-nutrient soil.
One pot with a mother ramet was then placed near a box and the stolon or creeping stem of the ramet was forced to grow along the short edge of the adjacent soil patches (Figure 1). Two weeks after the first daughter ramet rooted, the stolon/creeping stem internode connecting the mother ramet and its first daughter ramet was either severed to prevent clonal integration or kept intact (connected) to allow clonal integration (Figure 1). Then we removed the apex of the stolon/creeping stem to encourage the production of secondary and tertiary stolons and ramets in the box.
The experiment started on 1 September and ended on 1 December 2019, and was conducted in the same greenhouse as for material cultivation. During the experiment, we removed the additional stolons/creeping stems produced by the mother ramet. The mean temperature and mean relative humidity in the greenhouse were 27.1 °C and 86.3%, respectively. Water was added to the boxes and pots when soil surface became dry. To avoid mass flow of water and thus soil nutrients between adjacent patches within the box, water was added slowly to soil. The amount of water added each time varied depending on the weather conditions. During the experiment, the boxes with the pots were randomly repositioned three times to avoid potential differences in microclimatic conditions in the greenhouse.
2.4. Measurements
For the heterogeneous treatments, we counted the total number of new offspring ramets in the high- and low-nutrient patches separately for each box. Then, we harvested the first daughter ramet, all offspring ramets in the high-nutrient patches, and all offspring ramets in the low-nutrient patches in each box separately. For the homogeneous treatments, the plant in each box was harvested in a similar fashion, i.e., the first daughter ramet and offspring ramets in the imagined low- and high-nutrient patches were harvested separately. We also harvested the mother ramet in each pot. The plants were then separated into shoots (leaves plus stolons/creeping stems) and roots, dried at 70 °C for 72 h, and weighed.
2.5. Data Analysis
During the experiment, the first daughter ramet of H. vulgaris in one replicate of the severed homogeneous treatment died unexpectedly, and thus the data for this plant were excluded from analysis. For each species, we used two-way ANOVA to test the effects of soil heterogeneity (homogeneous vs. heterogeneous) and clonal integration (connected vs. severed, i.e., with vs. without integration) on biomass (total, shoot, and root) and the number of offspring ramets in the whole box and on biomass of the mother ramet in the pot. When a significant effect was observed, Tukey HSD tests were used for multiple comparisons. We also employed three-way ANOVA with repeated measures to test the effects of patch quality (low- vs. high-nutrient patches), soil heterogeneity, and clonal integration on biomass and number of the offspring ramets in patches within the box for each species. In this model, patch quality was treated as a repeated variable because the low- and high-nutrient patches in each box were not independent [65]. Following this model, planed linear contrasts were used to test whether the mean values of each variable differed between the low- and high-nutrient patches within each of the four combinations of clonal integration and soil heterogeneity. Before analysis, data on number of the offspring ramets of H. vulgaris in the box were transformed to square root and data on total biomass, root biomass, and number of offspring ramets of D. indica, and data on root biomass of the offspring ramets of G. longituba were log-transformed to satisfy the assumption of ANOVA. Also, data on root biomass of the mother ramet of H. vulgaris in the pot and data on total and shoot biomass of the mother ramet of D. indica were log-transformed. All analyses were conducted using SPSS 23.0 (IBM Corp., Armonk, NY, USA).
3. Results
3.1. Effects of Clonal Integration and Soil Heterogeneity on Biomass of the Mother in the Pot
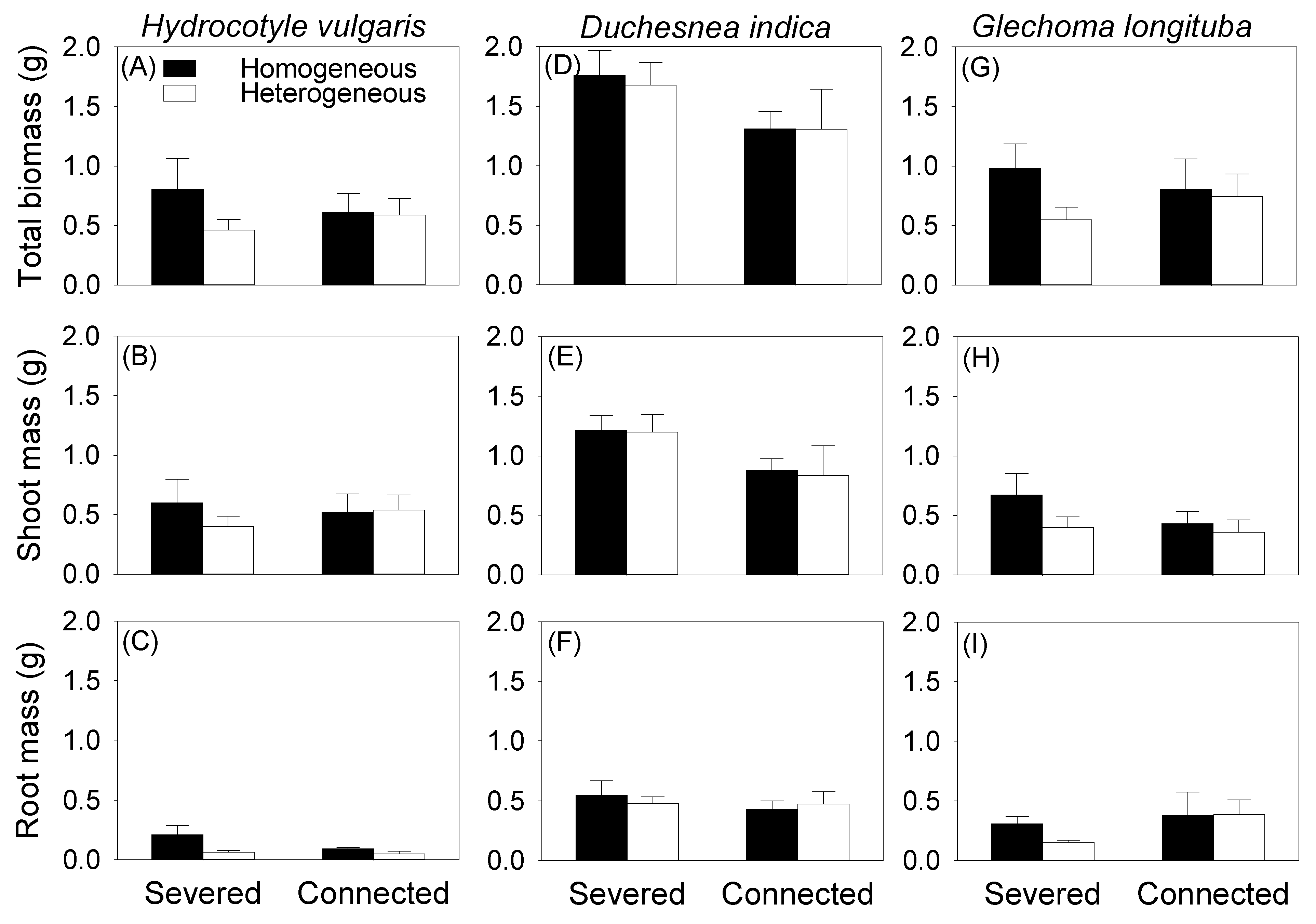
Clonal integration did not significantly affect total and root biomass of the mother ramet growing in the pot for any of the three species, and had no effect on shoot biomass of the mother ramet of H. vulgaris and G. longituba (Table 1, Figure 2). While the ANOVA result produced a significant effect of clonal integration on shoot biomass of D. indica (Table 1B), post hoc analysis demonstrated no significant difference (Figure 2E). Soil heterogeneity in the box had no significant effect on total, shoot, and root biomass of the mother ramet in the pot for any of the three species (Table 1, Figure 2).

Table 1.
ANOVA results for effects of clonal integration (I), soil heterogeneity (H), and their interaction on the growth of the mother ramet of each species.
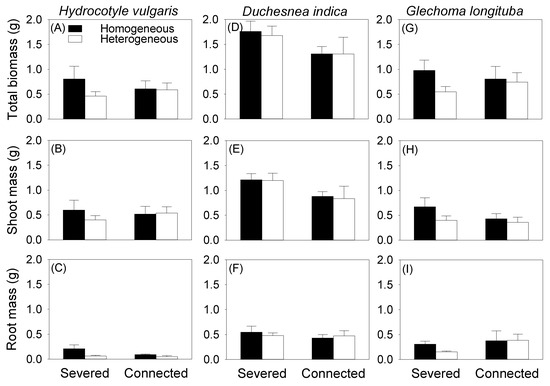
Figure 2.
Effects of soil heterogeneity (homogeneous vs. heterogeneous) and clonal integration (severed vs. connected) on total mass (A–C), shoot mass (D–F), and root mass (G–I) of the mother ramet of each species.
3.2. Effects of Clonal Integration and Soil Heterogeneity on Offspring Growth in the Box
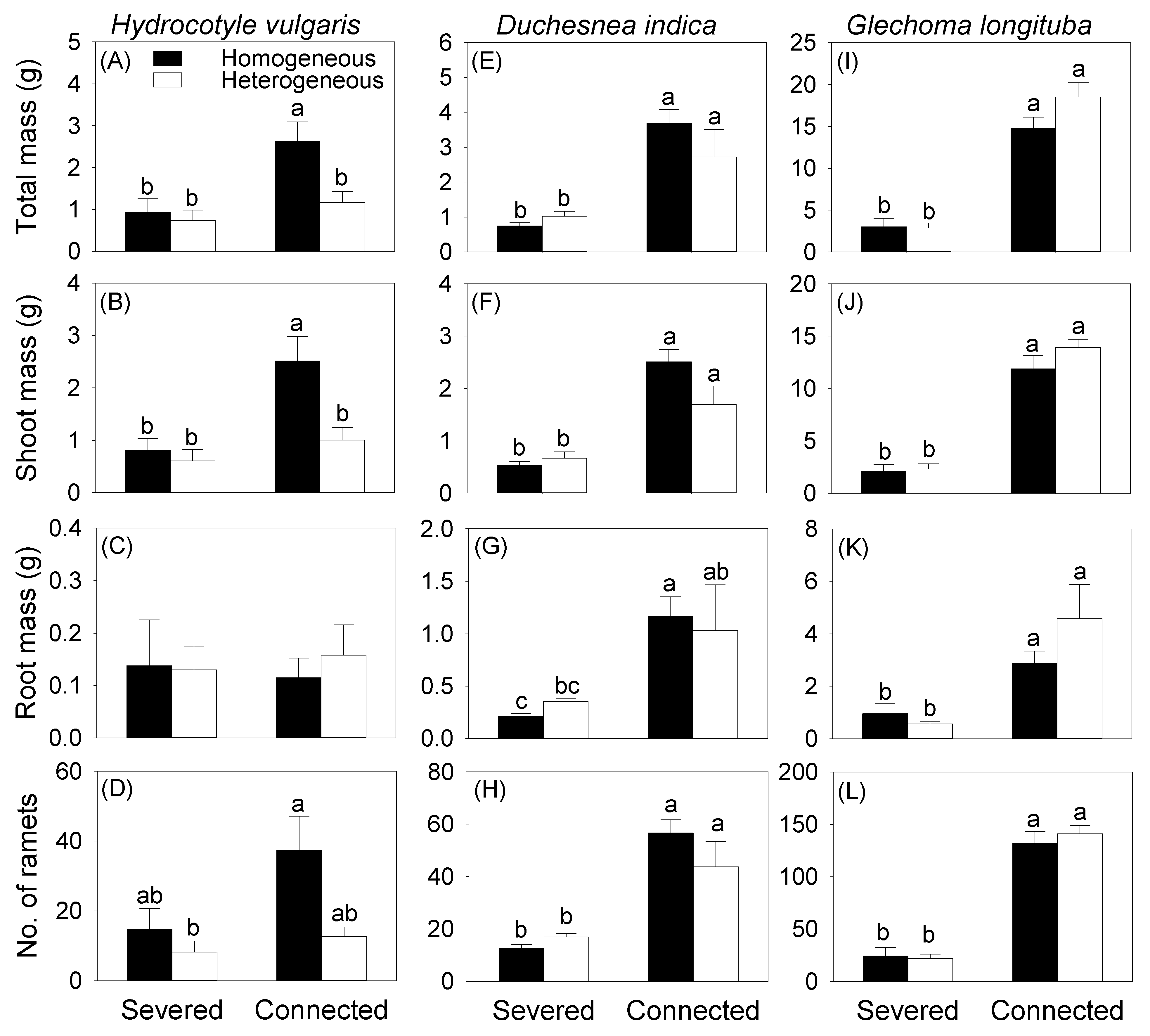
Generally, averaged across the two soil treatments, clonal integration from the mother ramet growing in the pot increased all growth measures of the offspring ramets growing in the box for all three species, except root biomass of H. vulgaris (Table 2, Figure 3). Without clonal integration from the mother ramet, soil heterogeneity had no effect on total, root, and shoot biomass and the number of the offspring ramets growing in the whole box for any of the three species (Figure 3). With clonal integration, soil heterogeneity also had no significant effect on either biomass or number of the offspring ramets growing in the whole box for D. indica (Figure 3E–H) and G. longituba (Figure 3I–L), but significantly decreased total and shoot biomass of the offspring ramets of H. vulgaris (Figure 3A,B).
Table 2.
ANOVA results for effects of clonal integration from the mother ramet growing in the pot (I), soil heterogeneity in the box (H), and their interaction on the growth of the offspring ramets of each species growing in the box.
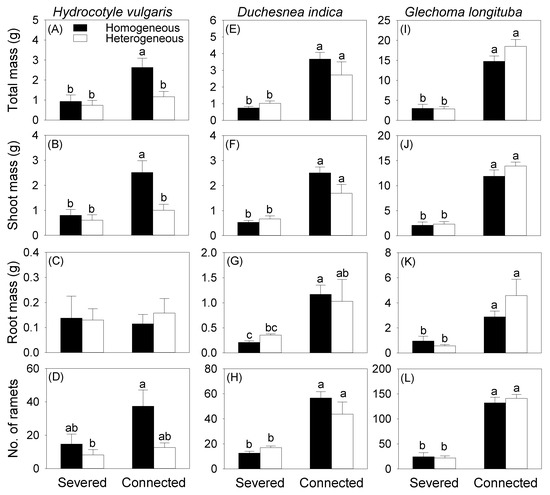
Figure 3.
Effects of soil heterogeneity (homogeneous vs. heterogeneous) and clonal integration (severed vs. connected) on biomass (A–C,E–G,I–K) and number (D,H,L) of the offspring ramets of each species growing in the box. Error bars indicate +1 SE. Different letters indicate significant differences (Tukey HSD tests).
3.3. Effects of Clonal Integration on Foraging Responses
Patch quality or its two-way and three-way interactions with clonal integration and soil heterogeneity had no significant effects on the growth of D. indica (Table 3B). There were significant interactive effects of patch quality × clonal integration and patch quality × clonal integration × soil heterogeneity on total biomass, shoot biomass, and number of ramets, and a significant effect of patch quality × clonal integration × soil heterogeneity on total stem length of the offspring ramets of H. vulgaris at the patch level (Table 3A). There were also significant interactive effects of patch quality × soil heterogeneity and patch quality × clonal integration × soil heterogeneity on total and shoot biomass, and a significant effect of patch quality × soil heterogeneity on number of the offspring ramets of G. longituba at the patch level (Table 3C).
Table 3.
ANOVA results for effects of clonal integration from the mother ramet growing in the pot (S), soil heterogeneity in the box (H), patch quality (Q), and their interactions on the growth of the offspring ramets of each species growing in the box.
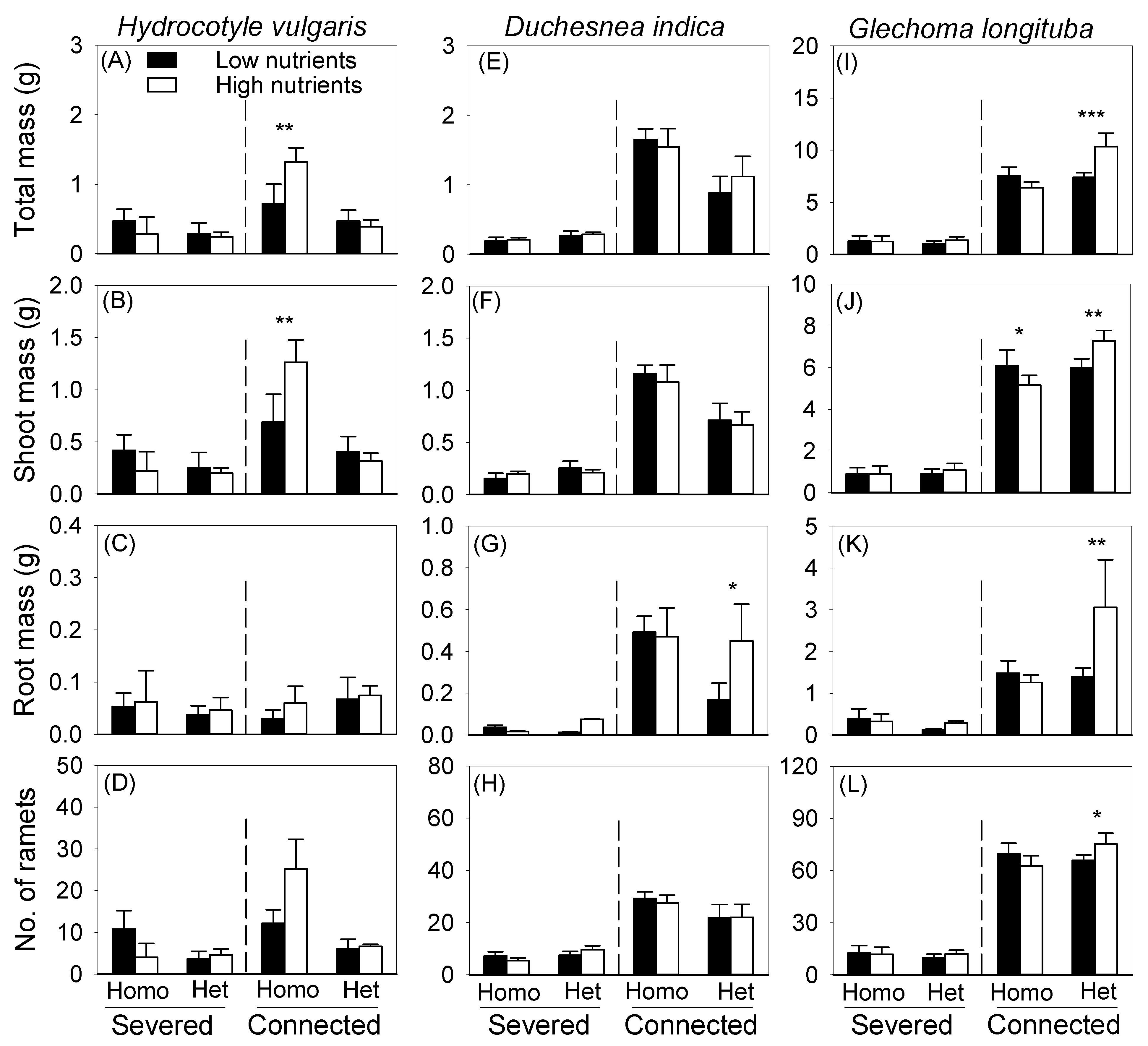
Without clonal integration from the mother ramet growing in the pot, the offspring ramets growing in the box did not display significant differences in total, shoot, and root biomass and number of ramets between the high- and low-nutrient patches in the heterogeneous treatment (Figure 4). This was true for all three species (Figure 4). With integration, the offspring ramets of H. vulgaris growing in the box also did not display significant differences in total, shoot, and root biomass and number of ramets between the high- and low-nutrient patches in the heterogeneous treatments (Figure 4A–D). However, with clonal integration, the offspring ramets of D. indica produced more root mass in the high- than in the low-nutrient patches in the heterogeneous treatment (Figure 4G), and those of G. longituba produced more total, root, and shoot biomass and ramets in the high-nutrient patches than in the low-nutrient patches in the heterogeneous treatment (Figure 4I–L). Unexpectedly, in the homogeneous treatment, total and shoot biomass of H. vulgaris and shoot biomass of G. longituba displayed significant differences between the two types of imagined patches (Figure 4A,B,J).
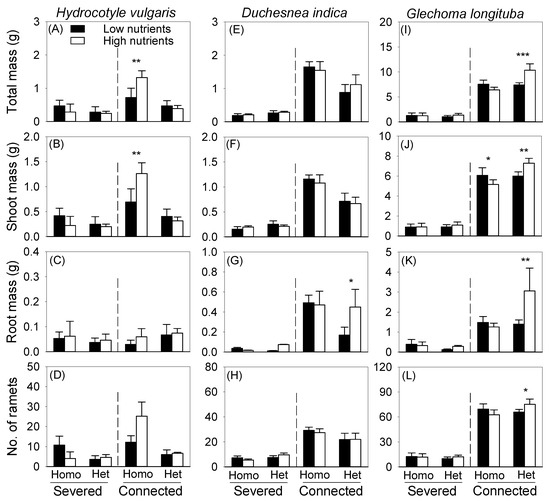
Figure 4.
Effects of soil heterogeneity (homogeneous vs. heterogeneous), clonal integration (severed vs. connected) and patch quality (high- vs. low-nutrient patches) on biomass (A–C,E–G,I–K) and number (D,H,L) of the offspring ramets of each species growing in the box. Error bars indicate +1 SE. Symbols (* p < 0.05, ** p < 0.01 and *** p < 0.001) indicate significant differences between the high- and low-nutrient patches within the box (by linear contrasts).
4. Discussion
As expected, clonal integration promoted the growth of the offspring ramets in both the homogeneous and heterogeneous soil environments for all three clonal plant species at no cost to their connected mother ramets (Table 1 and Table 2, Figure 2 and Figure 3). These results are consistent with previous findings of many experimental studies [10,15,18,20,66] and also two meta-analytical studies [18,19]. The promoted growth of the offspring ramets is likely due to the importation of carbohydrates, nutrients, and/or water translocated from their connected mother ramet, as reported previously [9,10,16,20,66]. Also, likely only surplus resources were exported from the mother ramet to the offspring ramets so that the loss of these amounts of resources could not reduce the growth of the mother ramet and thus result in a significant cost of clonal integration [10,67,68,69].
Without clonal integration from the mother ramet, the offspring ramets in the heterogeneous soil environment produced statistically the same amount of biomass and ramets in the two types of soil patches (Figure 4), suggesting that none of the three clonal species demonstrated either root- or shoot-foraging responses in the heterogeneous soil environment without the support from their mother ramet. With clonal integration, however, the offspring ramets of D. indica demonstrated root-foraging responses by producing more root biomass in the high- than in the low-nutrient patches in the heterogeneous soil environment, and those of G. longituba demonstrated both root and shoot foraging (Figure 4). In previous studies, the root-sprouting clonal plant Ajuga genevensis and the stoloniferous clonal plant Potentilla anserina did not demonstrate foraging responses [70,71]. However, foraging responses in heterogeneous environments have been demonstrated in many other clonal species, including Hierochloe glabra [58], Fragaria vesca [72], Potentilla reptans [40], and Glechoma hederacea [73]. Additionally, in a study with six pairs of invasive and native clonal plant species, Kesser et al. [59] found that the invasive species generally displayed root-foraging responses in heterogeneous soil environments, whereas the native species did not. Therefore, clonal species differ in the ability of root- and shoot-foraging responses in heterogeneous environments. Our results also suggest that clonal integration can modify the foraging responses of clonal plants in heterogeneous environments, but such an effect can vary with species.
No matter whether clonal integration was allowed or not, soil heterogeneity significantly promoted the growth of the offspring ramets in none of the three clonal species (Figure 3). This is despite that fact that in the presence of clonal integration, D. indica demonstrated root foraging and G. longituba displayed both root and shoot foraging. Many previous studies have suggested that soil heterogeneity can promote the growth of clonal plants [25,36,38,42,55,65,71,74]. For instance, Birch and Hutchings (1994) found that Glechoma hederaceae produced 2.5 times higher biomass in the heterogeneous soil environment than in the homogeneous environment with the same amount of nutrients. Similarly, in a recent study, Si et al. (2021) reported that two dwarf bamboo species (Sasa argenteosriata and Sasaella glabra) produced significantly more biomass in the heterogeneous than in the homogeneous soil environment even if the two environments contained the same amount of nutrients. However, there are also studies suggesting no effect of soil heterogeneity on the growth of clonal plants [32,71,75,76]. Thus, in response to soil heterogeneity, a significant foraging response of clonal plants may not necessarily result in their growth promotion, as also suggested in other studies [33,58,71].
Surprisingly, soil heterogeneity decreased the growth of one clonal species (H. vulgaris) in the presence of clonal integration (Figure 3A,B,D). Additionally and unexpectedly, in the homogeneous treatment total and shoot biomass of H. vulgaris and shoot biomass of G. longituba displayed significant differences between the two types of imagined patches (Figure 4A,B,J). The exact reasons for these unexpected results are unknown. It is likely that these came from some undetected disturbance noise during the experiment.
5. Conclusions
We conclude that clonal integration can alter root and/or foraging responses of clonal plants when they grow in heterogeneous environments, but this effect of clonal integration depends on the plant species. However, root- and/or shoot-foraging responses in heterogeneous environments may not necessarily be adaptative and thus benefit the growth of clonal plants. Our results do not support the idea that clonal integration can modify the effect of environmental heterogeneity on the growth of clonal plants. It should be noted that we considered only clonal integration between a mother ramet growing in a pot and their offspring ramets growing in heterogeneous environments. Future studies could consider the effect of clonal integration between ramets growing in heterogeneous environments on their foraging responses and potential growth benefits.
Author Contributions
X.-X.C. performed the experiment and wrote the first draft of the manuscript; W.X. and N.-F.L. designed the experiment and helped with data analysis; F.-H.Y. conceptualized the study and rewrote the manuscript based on the first draft. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by NSFC (grants 31870610, 32071527, 31761123001).
Institutional Review Board Statement
No applicable.
Informed Consent Statement
No applicable.
Data Availability Statement
The data are available on request.
Acknowledgments
We thank three anonymous reviewers for their comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pan, J.J.; Price, J.S. Fitness and evolution in clonal plants: The impact of clonal growth. Evol. Ecol. 2002, 15, 583–600. [Google Scholar] [CrossRef]
- Eckert, C.G. The loss of sex in clonal plants. Evol. Ecol. 2002, 15, 501–520. [Google Scholar] [CrossRef]
- Klimes, L.; Klimesova, J.; Hendriks, R.; Groenendael, J.V. Clonal Plant Architecture: A Comparative Analysis of Form and Function. In The Ecology and Evolution of Clonal Plants; De Kroon, H., Van Groenendael, J., Eds.; Backhuys: Leiden, The Netherlands, 1997; pp. 1–29. [Google Scholar]
- Jackson, J.B.; Buss, L.W.; Cook, R.E.; Ashmun, J.W. Population Biology and Evolution of Clonal Organisms; Yale University Press: New Haven, CT, USA, 1985. [Google Scholar]
- Xu, L.; Yu, F.-H.; Van Drunen, E.; Schieving, F.; Dong, M.; Anten, N.P. Trampling, defoliation and physiological integration affect growth, morphological and mechanical properties of a root-suckering clonal tree. Ann. Bot. 2012, 109, 1001–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adomako, M.O.; Alpert, P.; Du, D.-L.; Yu, F.-H. Effects of clonal integration, nutrients and cadmium on growth of the aquatic macrophyte Pistia stratiotes. J. Plant Ecol. 2020, 13, 765–772. [Google Scholar] [CrossRef]
- Wang, P.; Alpert, P.; Yu, F.-H. Physiological integration can increase competitive ability in clonal plants if competition is patchy. Oecologia 2021, 195, 199–212. [Google Scholar] [CrossRef]
- Gao, F.L.; Alpert, P.; Yu, F.H. Parasitism induces negative effects of physiological integration in a clonal plant. New Phytol. 2021, 229, 585–592. [Google Scholar] [CrossRef]
- Pennings, S.C.; Callaway, R.M. The advantages of clonal integration under different ecological conditions: A community-wide test. Ecology 2000, 81, 709–716. [Google Scholar] [CrossRef]
- Stuefer, J.F.; During, H.J.; De Kroon, H. High benefits of clonal integration in two stoloniferous species, in response to heterogeneous light environments. J. Ecol. 1994, 82, 511–518. [Google Scholar] [CrossRef]
- Si, C.; Alpert, P.; Zhang, J.-F.; Lin, J.; Wang, Y.-Y.; Hong, M.-M.; Roiloa, S.R.; Yu, F.-H. Capacity for clonal integration in introduced versus native clones of the invasive plant Hydrocotyle vulgaris. Sci. Total Environ. 2020, 745, 141056. [Google Scholar] [CrossRef]
- Gómez, S.; Onoda, Y.; Ossipov, V.; Stuefer, J.F. Systemic induced resistance: A risk-spreading strategy in clonal plant networks? New Phytol. 2008, 179, 1142–1153. [Google Scholar] [CrossRef]
- Lu, H.Z.; Brooker, R.; Song, L.; Liu, W.Y.; Sack, L.; Zhang, J.L.; Yu, F.H. When facilitation meets clonal integration in forest canopies. New Phytol. 2020, 225, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.-P.; Wei, G.-W.; Luo, F.-L.; Li, C.-Y.; Dong, B.-C.; Ji, J.-S.; Yu, F.-H. Effects of salinity and clonal integration on the amphibious plant Paspalum paspaloides: Growth, photosynthesis and tissue ion regulation. J. Plant Ecol. 2019, 12, 45–55. [Google Scholar] [CrossRef]
- Amsberry, L.; Baker, M.A.; Ewanchuk, P.J.; Bertness, M.D. Clonal integration and the expansion of Phragmites australis. Ecol. Appl. 2000, 10, 1110–1118. [Google Scholar] [CrossRef]
- Oborny, B.; Kun, Á.; Czárán, T.; Bokros, S. The effect of clonal integration on plant competition for mosaic habitat space. Ecology 2000, 81, 3291–3304. [Google Scholar] [CrossRef]
- Hutchings, M.J.; Wijesinghe, D.K. Performance of a clonal species in patchy environments: Effects of environmental context on yield at local and whole-plant scales. Evol. Ecol. 2008, 22, 313–324. [Google Scholar] [CrossRef]
- Song, Y.-B.; Yu, F.-H.; Keser, L.H.; Dawson, W.; Fischer, M.; Dong, M.; Van Kleunen, M. United we stand, divided we fall: A meta-analysis of experiments on clonal integration and its relationship to invasiveness. Oecologia 2013, 171, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xu, T.; Wang, Y.; Li, G.; Abdullah, I.; Zhong, Z.; Liu, J.; Zhu, W.; Wang, L.; Wang, D.; et al. A meta-analysis of effects of physiological integration in clonal plants under homogeneous vs. heterogeneous environments. Funct. Ecol. 2021, 35, 578–589. [Google Scholar] [CrossRef]
- Stuefer, J.F.; Gómez, S.; Mölken, T.V. Clonal integration beyond resource sharing: Implications for defence signalling and disease transmission in clonal plant networks. Evol. Ecol. 2004, 18, 647–667. [Google Scholar] [CrossRef]
- Jackson, R.; Caldwell, M. Geostatistical patterns of soil heterogeneity around individual perennial plants. J. Ecol. 1993, 81, 683–692. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, X.; Yu, F.; Dong, M.; Zhang, S.; Wang, R. Spatial heterogeneity of Ulmus pumila open forest ecosystem in Otindag sandy land. Environ. Sci. 2003, 24, 29–34. [Google Scholar]
- Alpert, P.; Mooney, H.A. Resource heterogeneity generated by shrubs and topography on coastal sand dunes. Vegetatio 1996, 122, 83–93. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Environmental heterogeneity and plant species diversity: A hypothesis. Am. Nat. 1977, 111, 376–381. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Birch, C.; Hutchings, M. Exploitation of patchily distributed soil resources by the clonal herb Glechoma hederacea. J. Ecol. 1994, 82, 653–664. [Google Scholar] [CrossRef]
- Adomako, M.O.; Xue, W.; Roiloa, S.; Zhang, Q.; Du, D.-L.; Yu, F.-H. Earthworms modulate impacts of soil heterogeneity on plant growth at different spatial scales. Front. Plant Sci. 2021, 12, 735495. [Google Scholar] [CrossRef]
- Adomako, M.O.; Xue, W.; Du, D.-L.; Yu, F.-H. Soil biota and soil substrates influence responses of the rhizomatous clonal grass Leymus chinensis to nutrient heterogeneity. Plant Soil 2021, 465, 19–29. [Google Scholar] [CrossRef]
- Hutchings, M.; De Kroon, H. Foraging in plants: The role of morphological plasticity in resource acquisition. Adv. Ecol. Res. 1994, 25, 159–238. [Google Scholar]
- De Kroons, H.; Hutchings, M.J. Morphological plasticity in clonal plants: The foraging concept reconsidered. J. Ecol. 1995, 83, 143–152. [Google Scholar] [CrossRef]
- Si, C.; Xue, W.; Guo, Z.-W.; Zhang, J.-F.; Hong, M.-M.; Wang, Y.-Y.; Lin, J.; Yu, F.-H. Soil heterogeneity and earthworms independently promote growth of two bamboo species. Ecol. Indic. 2021, 130, 108068. [Google Scholar] [CrossRef]
- Wijesinghe, D.K.; Hutchings, M.J. The effects of environmental heterogeneity on the performance of Glechoma hederacea: The interactions between patch contrast and patch scale. J. Ecol. 1999, 87, 860–872. [Google Scholar] [CrossRef]
- Liu, L.; Alpert, P.; Dong, B.-C.; Yu, F.-H. Modification by earthworms of effects of soil heterogeneity and root foraging in eight species of grass. Sci. Total Environ. 2020, 708, 134941. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Alpert, P.; Dong, B.-C.; Li, J.-M.; Yu, F.-H. Combined effects of soil heterogeneity, herbivory and detritivory on growth of the clonal plant Hydrocotyle vulgaris. Plant Soil 2017, 421, 429–437. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Fischer, M. Adaptive evolution of plastic foraging responses in a clonal plant. Ecology 2001, 82, 3309–3319. [Google Scholar] [CrossRef]
- Liang, J.-F.; Yuan, W.-Y.; Gao, J.-Q.; Roiloa, S.R.; Song, M.-H.; Zhang, X.-Y.; Yu, F.-H. Soil resource heterogeneity competitively favors an invasive clonal plant over a native one. Oecologia 2020, 193, 155–165. [Google Scholar] [CrossRef]
- Fransen, B.; De Kroon, H.; Berendse, F. Soil nutrient heterogeneity alters competition between two perennial grass species. Ecology 2001, 82, 2534–2546. [Google Scholar] [CrossRef]
- Xue, W.; Huang, L.; Yu, F.-H.; Bezemer, T.M. Intraspecific aggregation and soil heterogeneity: Competitive interactions of two clonal plants with contrasting spatial architecture. Plant Soil 2018, 425, 231–240. [Google Scholar] [CrossRef]
- Liu, L.; Dong, B.-C.; Alpert, P.; Yu, F.-H. Effects of soil substrate heterogeneity and moisture on interspecific competition between Alternanthera philoxeroides and four native species. J. Plant Ecol. 2017, 10, 528–537. [Google Scholar]
- Louâpre, P.; Bittebière, A.-K.; Clément, B.; Pierre, J.-S.; Mony, C. How past and present influence the foraging of clonal plants? PLoS ONE 2012, 7, e38288. [Google Scholar] [CrossRef] [Green Version]
- Yao, S.; Yu, J.; Zhang, L.; Lei, N.; Xue, W.; Chen, J.; Zhang, Q.; Yu, F. Effects of soil nutrient heterogeneity and earthworms on aboveground biomass of experimental plant communities. Phyton 2021, 90, 1259. [Google Scholar] [CrossRef]
- Xue, W.; Huang, L.; Yu, F.-H. Increasing soil configurational heterogeneity promotes plant community evenness through equalizing differences in competitive ability. Sci. Total Environ. 2021, 750, 142308. [Google Scholar] [CrossRef]
- Hutchings, M.J.; John, E.A.; Wijesinghe, D.K. Toward understanding the consequences of soil heterogeneity for plant populations and communities. Ecology 2003, 84, 2322–2334. [Google Scholar] [CrossRef]
- Adomako, M.O.; Gao, F.-L.; Li, J.-M.; Du, D.-L.; Xue, W.; Yu, F.-H. Effects of soil nutrient heterogeneity and parasitic plant infection on an experimental grassland community. Flora 2020, 271, 151666. [Google Scholar] [CrossRef]
- Wijesinghe, D.K.; John, E.A.; Hutchings, M.J. Does pattern of soil resource heterogeneity determine plant community structure? An experimental investigation. J. Ecol. 2005, 93, 99–112. [Google Scholar] [CrossRef]
- Geng, Y.; Van Klinken, R.D.; Sosa, A.; Li, B.; Chen, J.; Xu, C.-Y. The relative importance of genetic diversity and phenotypic plasticity in determining invasion success of a clonal weed in the USA and China. Front. Plant Sci. 2016, 7, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Q.-Y.; Alpert, P.; An, J.; Gao, J.-Q.; Han, G.-X.; Yu, F.-H. Clonal integration in Phagmites australis mitigates effects of oil pollution on greenhouse gas emissions in a coastal wetland. Sci. Total Environ. 2020, 739, 140007. [Google Scholar] [CrossRef]
- Xue, W.; Wang, W.-L.; Yuan, Q.-Y.; Yu, F.-H. Clonal integration in Phragmites australis alters soil microbial communities in an oil-contaminated wetland. Environ. Pollut. 2020, 265, 114828. [Google Scholar] [CrossRef]
- Zhang, L.-M.; Lu, H.-Z.; Alpert, P.; Song, L.; Liu, W.-Y.; Yu, F.-H. Higher benefits of clonal integration in rhizome-derived than in frond-derived ramets of the tropical fern Bolbitis heteroclita. Flora 2019, 257, 151415. [Google Scholar] [CrossRef]
- Dong, B.-C.; Zhang, L.-M.; Li, K.-Y.; Hu, X.-T.; Wang, P.; Wang, Y.-J.; Luo, F.-L.; Li, H.-L.; Yu, F.-H. Effects of clonal integration and nitrogen supply on responses of a clonal plant to short-term herbivory. J. Plant Ecol. 2019, 12, 624–635. [Google Scholar] [CrossRef]
- Portela, R.; Dong, B.-C.; Yu, F.-H.; Barreiro, R.; Roiloa, S.R. Effects of physiological integration on defense strategies against herbivory by the clonal plant Alternanthera philoxeroides. J. Plant Ecol. 2019, 12, 662–672. [Google Scholar] [CrossRef]
- Lin, H.-F.; Alpert, P.; Zhang, Q.; Yu, F.-H. Facilitation of amphibious habit by physiological integration in the clonal, perennial, climbing herb Ipomoea aquatica. Sci. Total Environ. 2018, 618, 262–268. [Google Scholar] [CrossRef]
- Wang, Y.J.; Müller-Schärer, H.; Van Kleunen, M.; Cai, A.M.; Zhang, P.; Yan, R.; Dong, B.C.; Yu, F.H. Invasive alien plants benefit more from clonal integration in heterogeneous environments than natives. New Phytol. 2017, 216, 1072–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Ali, A.; Yong, X.-H.; Lin, C.-G.; Niu, X.-H.; Cai, A.-M.; Dong, B.-C.; Zhou, Z.-X.; Wang, Y.-J.; Yu, F.-H. A multi-species comparison of selective placement patterns of ramets in invasive alien and native clonal plants to light, soil nutrient and water heterogeneity. Sci. Total Environ. 2019, 657, 1568–1577. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.C.; Wang, J.Z.; Liu, R.H.; Zhang, M.X.; Luo, F.L.; Yu, F.H. Soil heterogeneity affects ramet placement of Hydrocotyle vulgaris. J. Plant Ecol. 2014, 8, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Cain, M.L. Consequences of foraging in clonal plant species. Ecology 1994, 75, 933–944. [Google Scholar] [CrossRef]
- Liu, F.; Liu, J.; Dong, M. Ecological consequences of clonal integration in plants. Front. Plant Sci. 2016, 7, 770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Xing, F.; Jin, Y.; Nie, D.; Wang, Y. Foraging responses of clonal plants to multi-patch environmental heterogeneity: Spatial preference and temporal reversibility. Plant Soil 2012, 359, 137–147. [Google Scholar] [CrossRef]
- Keser, L.H.; Dawson, W.; Song, Y.-B.; Yu, F.-H.; Fischer, M.; Dong, M.; Van Kleunen, M. Invasive clonal plant species have a greater root-foraging plasticity than non-invasive ones. Oecologia 2014, 174, 1055–1064. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.-H.; Ji, M.-C.; Wang, Y.-Y.; Qiao, D.-D.; Chen, Y.-C. Study on invasion risk of Hydrocotyle vulgaris as an alien species in wetlands. J. Zhejiang Univ. Agric. Life Sci. 2011, 37, 425–431. [Google Scholar]
- Murphy, K.J.; Rørslett, B.; Springuel, I. Strategy analysis of submerged lake macrophyte communities: An international example. Aquat. Bot. 1990, 36, 303–323. [Google Scholar] [CrossRef]
- Wang, M.-Z.; Li, H.-L.; Liu, C.-X.; Dong, B.-C.; Yu, F.-H. Adaptive plasticity in response to light and nutrient availability in the clonal plant Duchesnea indica. J. Plant Ecol. 2022. [Google Scholar] [CrossRef]
- Liao, M.; Yu, F.; Song, M.; Zhang, S.; Zhang, J.; Dong, M. Plasticity in R/S ratio, morphology and fitness-related traits in response to reciprocal patchiness of light and nutrients in the stoloniferous herb, Glechoma longituba L. Acta Oecol. 2003, 24, 231–239. [Google Scholar] [CrossRef]
- Gao, J.-Q.; Duan, M.-Y.; Zhang, X.-Y.; Li, Q.-W.; Yu, F.-H. Effects of frequency and intensity of drying-rewetting cycles on Hydrocotyle vulgaris growth and greenhouse gas emissions from wetland microcosms. Catena 2018, 164, 44–49. [Google Scholar] [CrossRef]
- Roiloa, S.R.; Retuerto, R. Small-scale heterogeneity in soil quality influences photosynthetic efficiency and habitat selection in a clonal plant. Ann. Bot. 2006, 98, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.Y.; Schooler, S.S.; Van Klinken, R.D. Effects of clonal integration and light availability on the growth and physiology of two invasive herbs. J. Ecol. 2010, 98, 833–844. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Chen, Y.; Dong, M. Clonal integration enhances survival and performance of Potentilla anserina, suffering from partial sand burial on Ordos plateau, China. Evol. Ecol. 2002, 15, 303–318. [Google Scholar] [CrossRef]
- Estrada, J.A.; Wilson, C.H.; Flory, S.L. Clonal integration enhances performance of an invasive grass. Oikos 2020, 129, 1623–1631. [Google Scholar] [CrossRef]
- Pauliukonis, N.; Gough, L. Effects of the loss of clonal integration on four sedges that differ in ramet aggregation. Plant Ecol. 2004, 173, 1–15. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; During, H.J.; Li, L. Do clonal plants show greater division of labour morphologically and physiologically at higher patch contrasts? PLoS ONE 2011, 6, e25401. [Google Scholar] [CrossRef]
- Martínková, J.; Klimeš, A.; Klimešová, J. No evidence for nutrient foraging in root-sprouting clonal plants. Basic Appl. Ecol. 2018, 28, 27–36. [Google Scholar] [CrossRef]
- Waters, E.M.; Soini, H.A.; Novotny, M.V.; Watson, M.A. Volatile organic compounds (VOCs) drive nutrient foraging in the clonal woodland strawberry, Fragaria vesca. Plant Soil 2016, 407, 261–274. [Google Scholar] [CrossRef]
- Slade, A.; Hutchings, M. The effects of light intensity on foraging in the clonal herb Glechoma hederacea. J. Ecol. 1987, 75, 639–650. [Google Scholar] [CrossRef]
- Zhang, L.-M.; Alpert, P.; Yu, F.-H. Nutrient foraging ability promotes intraspecific competitiveness in the clonal plant Hydrocotyle vulgaris. Ecol. Indic. 2022, 138, 108862. [Google Scholar] [CrossRef]
- Wang, P.; Lei, J.-P.; Li, M.-H.; Yu, F.-H. Spatial heterogeneity in light supply affects intraspecific competition of a stoloniferous clonal plant. PLoS ONE 2012, 7, e39105. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, D.K.; Hutchings, M.J. The effects of spatial scale of environmental heterogeneity on the growth of a clonal plant: An experimental study with Glechoma hederacea. J. Ecol. 1997, 85, 17–28. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).