Abstract
Thuja occidentalis L. wood is desirabl, e for wooden structures that require wood density uniformity. Wood density is a wood quality indicator related to numerous morphological, mechanical, physiological, and ecological properties. This study aimed to investigate the regional, site, and tree-to-tree variations of T. occidentalis wood density and growth components through the analysis of X-ray densitometer data. A total of 287 trees were randomly sampled from 11 sites in the Abitibi-Témiscamingue regions in Québec, Canada. The average ring density at breast height was 356 kg m−3, with a small difference between earlywood and latewood (167 kg m−3), indicating a relatively uniform wood. Ring density and width varied significantly between the Abitibi and the Témiscamingue regions, mainly in the juvenile wood. Trees from the Abitibi region showed higher ring density than those in the Témiscamingue region. In contrast, the ring width was higher in the Témiscamingue region. Site, tree, and cambial age significantly (p < 0.001) affected wood density and growth components. However, the largest variation is due to the tree-to-tree variation, accounting for about 15%–31% of the total variation. Compared to ring widths, ring density components showed a considerably smaller tree-to-tree variation and higher variation (7.1%) with cambial age than ring widths (0.6%). Ring width correlated positively and significantly (p < 0.001) with average temperature and annual precipitation, while ring density correlated negatively and significantly (p < 0.001) with average temperature and annual precipitation.
1. Introduction
Eastern white cedar (EWC) (Thuja occidentalis L.), one of the arborvitae species native to North America, is widely distributed from the Gulf of Saint Lawrence in the east to southeastern Manitoba in the west and from southern James Bay in the north to the Lake States in the south [1]. Due to its low density and high natural durability [2], EWC wood is highly prized for indoor joinery, outdoor furniture, and patios. It has excellent dimensional stability, increasing its value for wooden structures subjected to moisture fluctuations [3]. For example, the average service life of an untreated EWC heartwood post is 27 years, compared to just 5 years for an untreated black spruce post [1]. Hence, products such as shakes, shingles, fence posts, and mulch made from EWC have considerable potential market value [2,4]. Therefore, EWC wood improves the ability of medium-density fiberboard (MDF) [5] and oriented strand board (OSB) [4] to withstand decay and termites [6,7]. Moreover, restrictions on treated wood provide additional opportunities for EWC wood. However, the wood quality variation of this species is least investigated among the Canadian eastern commercial species.
The biological nature of wood makes it a highly variable material [8,9]. For a given species, wood properties show genetic, intra-tree, inter-tree, and inter-site variation [10,11,12]. The wood formation and properties within trees vary with both time of growth and tree age [13,14]. The wide variability in wood characteristics makes it difficult to evaluate wood performance precisely. Therefore, a better understanding of wood variability within a species would be useful for both wood quality research and efficient wood use.
Wood density is a key quality criterion due to its high correlation with mechanical strength and end-use performance [9]. It measures the amount of cell wall substance and predicts physical and mechanical properties such as wood strength, shrinkage, and elasticity [8,9]. Wood density is mainly influenced by genotype, site, aging of the cambium, and growth rate [15,16,17]. Intra-ring wood density variation is indicative of wood uniformity [9]. It determines the suitability of wood for specific end-uses, especially for high-value-added applications. For instance, uniform wood density is recommended for slicing and veneer peeling.
Generally, tree growth is positively related to site index quality. The site index measures the site’s productivity and reports the height of dominant and co-dominant trees in a stand at a base age [18,19,20]. However, the effects of environmental factors on the wood density of conifers are inconsistent. For instance, Muller-Landau [21] showed that the wood density of tropical species differed significantly among sites and with soil fertility but was independent of rainfall, seasonality, and temperature. Wood density and tree mortality were higher in poor than in rich soils. According to Ackermann [22] and Guilley et al. [23], the wood density hardly changed with changing geographic location and site quality in pedunculate oak (Quercus robur L.) and sessile oak (Quercus petraea Liebl.), respectively. However, Bergès et al. [18] reported that the ecological factors affected wood density independent of ring width, but their effect was unimportant in sessile oak. Though, other studies assumed that the effects of ecological factors on wood density are included in the effects on tree rings in quejigo oak (Quercus faginea Lam) [24], in balsam fir (Abies balsamea (L.) Mill) [11] and in European oak (pedunculate and sessile oak) [10]. The problem often lies in high inter-tree variability, which is not considered by studying the relationship between the site and ring density, masking the true relationship [10,18]. Cambial age is also considered the main factor in the variation of ring density in Laricio pine (Pinus nigra Subsp. Salzmannii) [19], but most studies did not give enough importance to this parameter by studying the relationship between the site and the ring density, which can influence the results.
The relationship between radial growth and climate in EWC has been documented in many studies [25,26,27,28]. Tardif and Bergeron [26] reported that abundant summer precipitation, associated with cool temperatures, produced maximum EWC growth and that drought strongly affected the growth of trees from xeric sites. Trees from xeric sites also responded more intensely to drought than those from mesic sites. Housset et al. [27] revealed that radial growth of marginal populations in America was limited by short growing seasons and summer droughts, autumn, and spring precipitation negatively impacted growth, especially in wet soil stands. In contrast, studies of on-site variation in ring density and growth in this species were very limited. Maeglin [29] reported a little variation in the ring density of trees grown in Wisconsin regions. In a previous study, Bouslimi et al. [30] studied the radial and longitudinal variations of selected wood properties and the relationship to biological processes in EWC in three boreal stands. However, no study has investigated this species’ site or inter-tree variations in ring widths and density components. Thus, this study investigates the regional, site, tree-to-tree, and cambial age variations of ring density components and ring widths in T. occidentalis in the Quebec boreal forest (Abitibi-Témiscamingue).
2. Materials and Methods
2.1. Study Material
The study material was sampled in eleven sites (or stands) within the natural range of T. occidentalis in the Abitibi-Témiscamingue in the province of Quebec, Canada (Figure 1). Sites were selected along a South–North latitudinal gradient (from 46°42′ N to 48°52′ N) to cover the full distribution of the species in this region and a wide range of ecological conditions (Figure 1): six stands in the Abitibi region (A1, A2, A3, A4, A5, and A6) and five stands in the Témiscamingue region (T1, T2, T3, T4, and T5). The study area is divided into two bioclimatic zones based on the abundance of EWC (Figure 1). The continuous zone (CZ) falls into the balsam fir and yellow birch (Betula alleghaniensis Britton) bioclimatic domain and represents an area where cedar is common [31,32]. The discontinuous zone (DZ) is in the balsam fir and white birch (Betula papyrifera Marsh.) bioclimatic domain. It marks the northern edge of the continuous distribution, where cedar becomes less common in the forest matrix. The site occupation rates by EWC along the gradient were estimated to be 55%–87% and 9%–33% in the continuous and discontinuous zones, respectively [27,32].
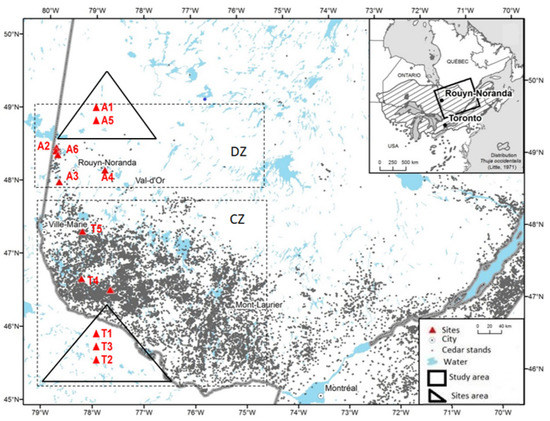
Figure 1.
Locations of Thuja occidentalis sample sites (doted in red) in the Abitibi-Témiscamingue, Québec, Canada. Sampling sites were stratified between the continuous zone of distribution (CZ) and the discontinuous zone (DZ) [32].
Stand and tree characteristics are summarized in Table 1. Climatic data for each site were obtained from weather data from 1930 to 2007 [33]. Mean annual temperatures tend to decrease towards the northeast of the study area, while total annual precipitation decreases towards the northwest. In the Abitibi region, stands are dominated by balsam fir and cedar, except site A1, which is dominated by spruce and cedar. However, in the Témiscamingue region, stands are almost mixed, and generally, cedar is associated with yellow birch, except for site T5, where cedar is associated with balsam fir and spruce. Site A2 is naturally regenerated from fires between 1717 and 1760 [34]. The soil in site T5 has a high moisture content (37.2%) [35], which explains the lower growth rate of EWC trees (Table 1).

Table 1.
Stand and T. occidentalis tree characteristics.
Forty trees were randomly selected from each site, and total height and diameter at breast height (DHP) were measured using steel tape (Table 1). The age of selected trees was also determined from the increment cores sampled at breast height using a QTRS-01X, a Tree-Ring X-Ray Scanner (Quintek Measurement Systems, Knoxville, Tennessee) according to the procedure described in Koubaa et al. [36].
2.2. Collection and Preparation of Wood Samples
In this study, one 6-mm increment core was collected from each sampled tree at breast height in the North–South direction (bark to bark passing through the pith). Increment cores were wrapped individually in a plastic bag and frozen to prevent decay until preparation. After air-drying, cores were sawn longitudinally to approximately 1.57 mm thick, the thickness required for accurate Tree-ring X-Ray Scanner analysis. The strips (one strip for each increment core) were extracted using a cyclohexane/ethanol solution 2:1 (v/v) for 24 h and distilled water for another 24 h to remove resinous substances and water-soluble carbohydrates. Extractives can account for from 1% to 20% of the oven-dry weight of wood of various tree species and can appreciably influence wood density [37]. Extractives are unequally distributed from the pith to the bark and can be considerably influenced by tree age, genetics, and site conditions [38]. Accordingly, by removing the extractives, we controlled their effect on wood density, which permits more accurate comparisons of change in wood density with site and tree. Samples were then conditioned to 8% equilibrium moisture content before measurement.
2.3. Ring Width and Wood Density Measurement
Ring widths and wood density components were measured using a QTRS-01X, a Tree-Ring X-ray Scanner (QMC, Knoxville, Tennessee), as described in Bouslimi et al. [39]. The mass attenuation coefficient (cm2 g−1) required to calculate the density was determined using the maximum moisture content method [40]. A linear resolution of 20 µm was used for X-ray densitometry measurements. After conditioning, rings from the pith to the bark were scanned in air-dry condition.
From the wood density profiles, average (RD), earlywood (EWD), and latewood (LWD) densities and average (RW), earlywood (EWW), and latewood (LWW) widths were calculated for each annual ring. The demarcation between earlywood and latewood was determined for each annual ring using a six-degree polynomial by the maximum derivative method [36]. The density of the demarcation point of the polynomial was defined as the transition density (TD). Latewood proportion (LP) was calculated as the ratio of latewood width to annual ring width. Cross-dating ring width chronologies detected false and missing rings. Cross-dating was statistically verified using COFECHA software [41]. During scanning, precautions were also taken to eliminate rings with compression wood or branch traces. Decayed strips were discarded. Accordingly, 287 strips were retained for this analysis. The number of analyzed trees for each site is given in Table 1.
The radial variation in wood traits was frequently used to estimate the transition age from juvenile to mature wood. Considering the radial pattern of EWC wood traits (wood density components, ring widths, and tracheid length and width), the wood produced from the pith up to the 30th ring is considered juvenile wood, and the remaining mature wood [30].
2.4. Statistical Analysis
Wood density components and ring widths were subjected to variance analyses (ANOVA) using a mixed-model approach, with cambial age as the repeated measure [40]. The site and cambial age were considered fixed effects, and the tree was considered random. The hierarchical effects of individual trees and sites were accounted for using two nested levels, with the tree effect nested within the site effect, as follows:
where Y is the dependent variable, µ is the grand mean, αi is the fixed effect of cambial age, βj is the fixed site effect, γk is the random tree effect tree, and ε is the residual error. The analysis did not consider data beyond the 70th annual ring from the pith. Because the analysis excluded the first annual ring, data included 69 repeated measures (rings 2 from the pith to 70).
Yijk = µ + αi + βj + γk + ε
The SAS mixed-model procedure (PROC MIXED) was used to fit the models using restricted maximum likelihood (REML) [42,43]. Instead of assuming independent errors, we imposed a first-order autoregressive correlation structure AR (1). The Kenward–Roger method [40] determined the degrees of freedom. Homoscedasticity and residual normality were verified after data log transformation. The F-test determined the statistical significance of the fixed effects, while the Z-tests determined the significance of the random effects. Tukey’s multiple comparison test investigated the significant statistical differences in wood properties between sites. Differences were considered statistically significant at p ≤ 0.05. Variance components were estimated as a percentage of all effects’ total variation (VAR) using the VARCOMP procedure [42].
For each annual ring, the mean and coefficient of variation (CV) of wood density properties at breast height for 287 EWC trees in the Abitibi-Témiscamingue were calculated. The CV shows the extent of variability in the population’s mean [9]. Furthermore, to determine the effect of region, the means of each variable were subjected to analysis of variance (ANOVA). Tukey’s multiple comparison method investigated the significance of the difference (p ≤ 0.05) between the wood properties of the Abitibi and Témiscamingue regions [42]. Correlation analyses established relationships between ecological factors (latitude, longitude, elevation, average annual temperature, total annual precipitation, stand density, tree height, and DHP) and wood density properties. Average annual temperature and total annual precipitation values (from 1930 to 2007) were determined according to Régnière and St-Amant [33].
3. Results and Discussions
3.1. T. occidentalis Wood Density Characteristics in the Québec Forest
The percentage of the total T. occidentalis stems is calculated for each stand [30], and it ranges from 41 to 81 (%) (Table 1). The Témiscamingue stands have a higher growth rate than the Abitibi stands. The total height and DHP were more important in the Témiscamingue stands. The sampled trees ranged from 40 to 198 years, with an average age of 66–95 for the sampled stands (Table 1).
EWC trees grown in Abitibi-Témiscamingue (Table 2) have higher annual ring density (RD) (356 kg m−3) than those grown in Wisconsin (281–324 kg m−3) [29]. Latewood density (LWD) was 35% higher than earlywood density (EWD) (Table 2), owing to the structural differences between the tracheid cell walls of latewood and earlywood [30]. A thick cell wall characterizes the latewood tracheids of this species compared to earlywood tracheids [30]. However, the difference between LWD and EWD in the studied trees was low (only 167 kg m−3) (Table 2) compared to that reported in other conifers such as jack pine (Pinus banksiana Lamb) (383 kg m−3) [44] and black spruce (Picea mariana (Mill.) B.S.P.) (243–383 kg m−3) [15,45], which are the most commercial species in North America. Furthermore, the difference in RD between juvenile and mature wood was minor in EWC (18 kg m−3), which could increase the value of this species for use in wooden structures that require wood uniformity, as veneer peeling and slicing are an excellent end-use where a high degree of uniformity is very beneficial [46].
Table 2.
Means and coefficients of variation (CV) (in parentheses (%)) of ring widths and ring density components at breast height in Thuja occidentalis in the Québec Boreal Forest and as a function of the site.
In agreement with a previous study on EWC [30], the pith-to-bark variation in studied wood traits revealed that the transition from juvenile to mature wood is gradual in this species (Figure 2). The RD was higher near the pith (402 kg m−3) and decreased slowly to reach the minimum (350 kg m−3) in the juvenile-mature wood transition zone (at about age 30) and remained constant or decreased steadily thereafter (Figure 2a). The coefficients of variation (Figure 2b) are higher from the pith up to age 30 (14%–20%), at which point the variation lessened (at around 10%). The larger variation observed within the first 30 rings is linked to the juvenile wood characteristics [9,47]. According to Alteyrac et al. [46], the proportion of juvenile wood tends to be higher in a suppressed (as T. occidentalis) than in a dominant tree. EWD followed a similar pattern to that of RD (Figure 2a). In contrast, LWD showed a completely different radial pattern (Figure 2a). It was low (441 kg m−3) and almost constant near the pith up to the 10th ring, where it increased to reach a maximum (500 kg m−3) in mature wood and remained constant over the tree age thereafter (Figure 2a). The variation coefficients related to cambial age were greater in LWD than in EWD and RD (Figure 2b). Transition density (TD) followed a similar pattern to that of LWD (Figure 2a). The radial pattern variation is related to the age of the cambium and changes in tree structure and function, as discussed in previous findings [30].
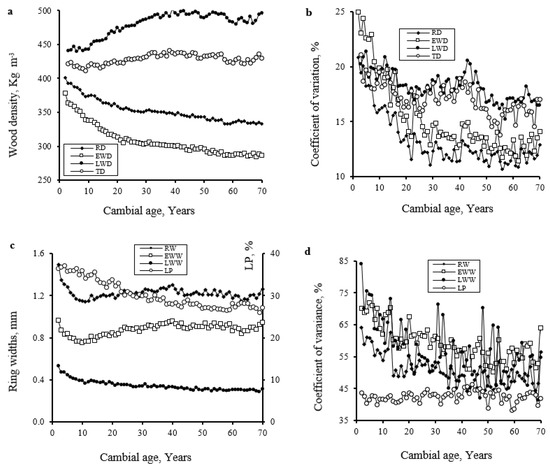
Figure 2.
Means (a,c) and coefficient of variations (b,d) of annual ring density (RD, kg m−3), earlywood density (EWD, kg m−3), latewood density (LWD, kg m−3), transition density (TD, kg m−3), ring (RW, mm), earlywood (EWW, mm), latewood width (LWW, mm), and latewood proportion (LP, %) as related to cambial age (from the pith) at breast height in 287 Thuja occidentalis trees growing in Abitibi Témiscamingue, Québec, Canada.
EWC has a low growth rate (1.28 mm) (Table 2) compared to other species, such as black spruce (Picea mariana (Mill.) B.S.P.) (2.51 to 3.34 mm) [48,49] and balsam fir (2 mm) [50]. T. occidentalis was always associated with late successional stages [51], which limited resource acquisition and negatively affected its growth compared to dominant trees [30,52,53,54]. The annual ring width (RW) showed a slight variation with cambial age (Figure 2c), decreasing from a maximum near the pith (1.38 mm) to a minimum at the 10th ring (1.15 mm), increasing gradually thereafter to reach a maximum (1.30 mm) in the transition zone and remaining relatively constant with age afterward (Figure 2c). Earlywood width (EWW) showed a similar pattern to RW (Figure 2c). In contrast to RW and EWW, latewood width (LWW) was relatively constant (Figure 2c) over tree age (0.4 mm). Accordingly, the distribution of latewood (LP) was also relatively uniform within the tree (Figure 2c), accounting for over 31% of the wood (Table 2). Thus, the EWC wood is more uniform, which results in high dimensional stability but low strength. This could make the wood easier to machine and produce a more uniform veneer which would be an advantage for applications that require wood uniformity [4,7]. The LP value of this species is higher than previously reported with most commercial species [44,45,50]. For instance, this value ranged from 23.9 to 25.2% in jack pine [44] and 11.8% in balsam fir [11,50].
Bouslimi et al. [30] showed that in the first years of growth (the first 10 years), RW decreases owing to increased competition from other undergrowth in the same stand, causing an increase in the LP near the pith (Figure 2c). This could explain the higher RD value near the pith (Figure 2a). When subsequent growth conditions are good, RW increases, which increases the earlywood proportion but decreases the latewood proportion (Figure 2c). RW increased sharply after cambial age 10 (Figure 2c), probably because the competition with undergrowth decreased with increasing years, and tree seedling establishment resumed [30].
In Abitibi-Témiscamingue, RD components showed a considerably smaller variation due to the tree effect than RW components (Table 2). These results agree with earlier conifers’ studies [11,49,50,55]. Among studied traits, RD, EWD, and LWD showed the smallest variation (<19%), followed by LP (44%), while RW, EWW, and LWW showed remarkably larger variation (52%–62%) (Figure 2, Table 2). Furthermore, the between-tree variation related to cambial age was greater in latewood than in earlywood (Figure 2b,d). A similar result was noted by Zhang [15] in black spruce and Koga and Zhang [50] in balsam fir.
3.2. Regional Variation of Ring Widths and Ring Density Components
The variation in average ring width and density components of the whole tree and juvenile and mature wood with the region in T. occidentalis are shown in Table 3. The regional variation in wood density traits related to cambial age is also presented in Figure 3. The analysis of variance (Table 3) showed that the region significantly affected most wood density traits (p < 0.001). The variation due to region was more important in juvenile than in mature wood, as indicated by the lower F-value for mature than for juvenile wood (Table 3).
Table 3.
Regional comparison of ring density components and ring widths for the whole tree and juvenile and mature wood at breast height in Thuja occidentalis in the Abitibi-Témiscamingue, Quebec, Canada.
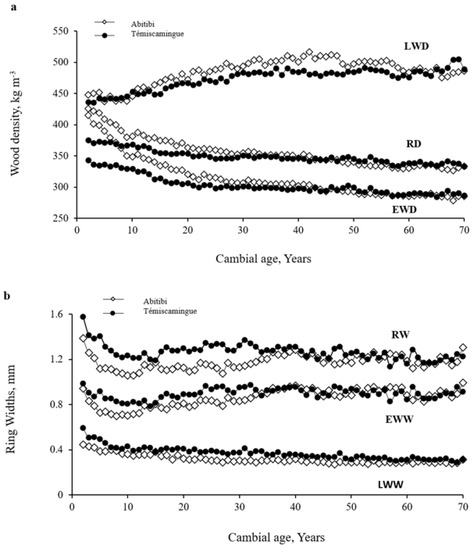
Figure 3.
Regional variation in (a) annual ring density (RD, kg m−3), earlywood density (EWD, kg m−3), latewood density (LWD, kg m−3), (b) ring width (RW, mm), earlywood width (EWW, mm) and latewood width (LWW, mm) in Thuja occidentalis in the Abitibi-Témiscamingue, Quebec, Canada.
The average RD varied significantly across regions (F= 60.80; p < 0.001) in juvenile wood (Table 3, Figure 3a). Trees in Abitibi have a higher average RD (378 kg m−3) than those in Témiscaminge (359 kg m−3). EWD and LWD were also higher in Abitibi than in Témiscamingue (Table 3, Figure 3a). In contrast, the average RW, EWW, and LWW were relatively higher in Témiscamingue (Table 3, Figure 3b). There are several causes for the variation of growth between both regions, such as ecological, edaphic, and environmental conditions. However, the climatic characteristics of each area seem to be the main factor influencing the growth of this species [27,31,56,57,58]. Abundant summer precipitation, associated with cool temperatures, produced maximum EWC growth; in contrast, radial growth was negatively correlated with warm summer temperatures (R = −0.3 to −0.5) and was limited by short growing seasons [26]. Overall, the wood formation was influenced by temperature and the quantity and timing of precipitation [26,27,57,58,59,60]. Housset and al. [27] reported a negative correlation with the June temperature of the current year of ring formation was also observed (r = −0.5) in this species. Radial growth was also positively correlated with precipitation in June preceding ring formation. In contrast, excess precipitation during autumn preceding ring formation negatively correlated with EWC growth.
According to Fromm [59], trees grow synchronously with the seasons and can endure periods unfavorable for growth by dormancy. In trees of boreal forests with very cold winters, changes in photoperiod and temperature are the dominant environmental signals regulating seasonal growth cycling. Thus, favorable growth conditions could induce cambial divisions and consequently positively influence tree growth [59]. In Témiscamingue, the average temperature and the annual precipitation were more important than in Abitibi (Table 1). According to Regnière and St-Amant [33], the frost-free period was approximately 150 days in Témiscamingue compared to only 80 days in the Abitibi region. The combination of these factors could stimulate tree growth, increasing the width of the ring in Témiscamingue [61,62]. The increase in the RW (Table 3, Figure 3) in the Témiscamingue site could negatively influence the RD [49,50,63] because of changes in cellular structure [30]. The proportion of earlywood and latewood in the growing ring determines the overall density of the ring [30]. As the RW in T. occidentalis increases, the width of the earlywood increases without a corresponding increase in the amount of latewood (Figure 2c), causing lower RD (Figure 2a). For T. occidentalis, cell and lumen diameters decreased from early- to latewood, while earlywood cells have a thin wall layer, which is one of the reasons for the lower density [39]. These could explain the decrease in RD in Témiscamingue compared to the Abitibi region (Figure 3).
Allowing to Larocque [60], the environment is a relevant factor in wood density determinism via the effects of ring width and cambial age on wood density. According to Zhang et al. [49], the decrease in RD combined with the increase in RW in conifers was more pronounced in species that showed a gradual transition from earlywood to latewood (as T. occidentalis) than in species that made abrupt transitions. Furthermore, Koga and Zhang [50] reported that, in conifers, an increased growth rate usually leads to a greater increase in earlywood than in latewood formation and delays the transition from juvenile to mature wood [49].
The difference between both regions for most wood density properties, except for LWD, was more important in juvenile wood than mature wood (Figure 3, Table 3). This difference was higher near the pith and tended to decrease with cambial age up to age 30–35 years (Figure 3), where the variation between Abitibi and Témiscamingue regions was small afterward (Table 3). For instance, the difference in RD between both regions was 61 kg m−3 at age 2, then decreased progressively to about 6 kg m−3 at age 30 and remained relatively constant thereafter (Figure 3a).
The high difference in RD observed near the pith between both regions could be linked to the variation of compression wood occurrence [9,64], associated with soils and environmental characteristics of each region, and, to some extent, to the seedling origin. In Abitibi, EWC is associated with late successional stages (nondominant tree); thus, it grows more slowly owing to more limited resource acquisition than the dominant tree. In the study sites, EWC is mainly associated with balsam fir and spruce (Table 1). These species showed a more rapid growth rate than EWC [26,51,65]. Thus, the competition understory associates could negatively affect the cambial activity and consequently favors the production of compression wood, especially when the layering accounts for a considerable amount of cedar reproduction in this area [32,51,66]. Compression wood is characterized by higher density and latewood proportion than normal wood [8,47]. According to Rosner [67], high wood density is supposed to be a common strategy to guarantee low vulnerability to cavitation. Thus, species with higher wood density are better able to resist drought-induced embolism [68]. The topology of the xylem network could also affect water transport and growth [69]. Under adverse conditions, EWC tended to sectorize its radial architecture to almost isolate the hydraulic pathways completely to different parts of the stem and crown, thereby increasing the odds of survival but significantly decreasing its growth rate [52,70]. The hydraulic sectorial in branches, stems, and taproots may be regarded as an adaptive trait to cope with environmental stress [53].
3.3. Inter-Site, Among-Tree, and with Cambial Age Variations of Ring Width and Wood Density Components
The variation in ring density components and ring widths with site, tree, and cambial age in T. occidentalis are presented in Table 2. The radial variation in RD and RW at selected sites is also shown in Figure 4. Considering all data (rings: 2–70), the analysis of variance (Table 4) showed that site, as well as tree and cambial age, significantly (p < 0.001) affected wood density traits. All properties showed a higher variation with cambial age, between sites, and among trees within the same site (Table 4, Figure 4). However, the largest variation is due to the tree, accounting for about 15%–32% of the total variation (Table 4), linked to the age difference of studied trees and consequently to tree circumference. RD components (RD, EWD, and LWD) showed a higher variation with cambial age than RW components (RW, EWW, LWW, and LP) (Table 4), which is mainly attributable to the juvenile and mature wood characteristics [9]. Compared to mature wood, T. occidentalis juvenile wood is characterized by a higher variation in cell dimensions and cell wall formation [30]. The LP (Figure 2c) was higher near the pith, decreasing gradually to reach a minimum in the transition zone and remaining constant thereafter. Accordingly, the EWC density variation is less important in mature wood than in juvenile wood. In contrast, RW components showed a larger tree-to-tree variation within the same site than RD components (Table 2 and Table 4). According to Fujiwara and Yang [71], the circumferential growth rate is the key to understanding the between-tree variation in RW in the same stand. The competition between trees in the same stand due to differences in crown structure and canopy position could considerably affect xylem development in EWC [72]. This might explain the larger between-tree variation in RW in this species. Furthermore, many studies suggest that the underlying cause of between-tree variation in RW is the relationship of crown ratio to the length of the stem devoid of branches, which is tree-age dependent [73,74,75]. As shown in Table 1, the tree age showed a significant variation in the same and among sampling sites, which could explain the high variation of RW between trees.
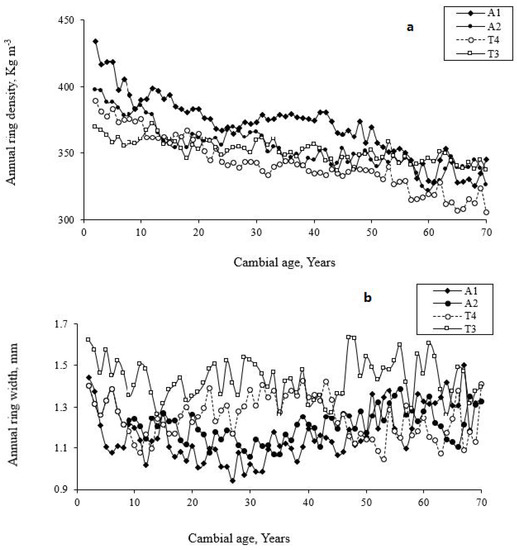
Figure 4.
Variation of (a) annual ring density (RD, kg m−3) and (b) annual ring width (RW, mm) in Thuja occidentalis at selected sites (A1, A2, T3, and T4) in the Abitibi-Témiscamingue, Quebec, Canada.
Table 4.
Linear mixed model analysis of variance, with F-value for fixed effects, Z-value for random effects and their significance for each source of variation, and variance components (VAR) for wood density components and ring widths in Thuja occidentalis.
It is essential to note that in Abitibi-Témiscamingue, the variation due to tree, cambial age, and site in wood traits were less important in EWC compared to those reported in many other commercial species such as jack pine, Brutia pine (Pinus brutia Ten), white spruce (Picea glauca (Moench) Voss), Scots pine (Pinus sylvestris L.), and cork oaks (Quercus suber) [12,44,76,77,78,79].
The variation due to cambial age was more important in juvenile than in mature wood, as indicated by the lower F-value for mature than for juvenile wood (Table 4). In juvenile wood, the cambial age effect was highly significant (p < 0.001) for most properties, accounting for up to 14.5% of the total variation. However, in mature wood, it was only significant on RD (p < 0.001), EWD (p < 0.001), and LP (p < 0.05), accounting for only 1.1%–3.2% of the total variation. This effect was associated with RD and RW components variation between juvenile and mature wood (Figure 2). In mature wood, RW components and LWD were relatively constant over the tree age (Figure 2); therefore, these traits’ cambial age effect was nonsignificant (Table 4).
The cambial age effect on RD was highly significant (p < 0.001), accounting for 12.7% of the total variation (Table 4). This effect was associated with RD variation between juvenile and mature wood (Table 4, Figure 2a). EWD and LWD showed similar results (Table 4, Figure 2a). The site effect on RD was also highly significant (p < 0.001), although it accounts for only 3.1% of the total variation (Table 4; Figure 4a). This could be linked to the cambial age effect and the high tree-to-tree variation.
The tree effect on EWD and LWD was highly significant (p < 0.001), accounting for 14.3% and 31.6% of the total variation, respectively (Table 4). For EWD, the low tree-to-tree variation could be explained by the large EWD variation in the juvenile wood phase, while the LWD is characterized by constancy over the cambial age (Figure 2a). Thus, the cambial age effect was greater in EWD than in LWD (Table 4), accounting for 18.4% and 6.2% of the EWD and LWD variation, respectively. When considering juvenile and mature wood separately, the variation due to trees was more important (Table 4). It accounted for 23% and 34% of the EWD variation in juvenile and mature wood, respectively, and about 43% of the LWD variation in both kinds of wood. The tree-to-tree variation of RD, EWD, and LWD could be explained by several factors, including vegetative competition among trees, tree age, and soil conditions [19,61,80,81,82]. In mature wood, tree circumference increased with increasing age [71], which could positively influence RW (Figure 4b) and consequently negatively affect the RD (Figure 4a) [50].
The site effect on RW, EWW, and LWW was highly significant (p < 0.001); however, it accounted for only a small proportion (3.8–9.9%) of the total variation (Table 4) and could be explained by the large tree-to-tree variation of these traits within the same stand (Table 4 and Figure 2d). For example, the RW showed a remarkably larger tree-to-tree variation in this study, as indicated by the large coefficients of variation (42%–56%) (Table 2 and Figure 2d). The tree effect on RW accounted for 27.3% of the total variation (Table 4). The variation of RW between trees is mainly linked to the variation of crown and canopy structures with tree and age [13,73]. According to Gartner et al. [83], the removal of 50% of the live crown in young trees in Douglas-fir (Pseudotsuga menziesii (Mirb.)) caused a decline in RW.
The tree effect on EWW, LWW, and LP was highly significant (p < 0.001) and accounted for 28%–44% of the total variation (Table 4). The between-tree variation was lower in LWW than in EWW. The site variation, however, was greater in LWW than in EWW (Table 4). It accounted for 9.9% and 3.8% of the LWW and EWW variation, respectively, and depended on the wood type (Table 4). For EWW, the low site variation could be explained by the greater tree-to-tree variation (27%–43% of the total variation) and the age effect in the juvenile wood phase (Figure 2c, Table 4). In contrast to EWW, LWW was constant over tree age (Figure 2c).
The highly significant effect of cambial age on RW (p < 0.001) accounts for only 0.6% of the total variation (Table 4). In EWC’s mature wood, the RW is almost constant over the cambial age; therefore, the cambial age effect on RW was negligible (0% of the total variation) (Table 4). The error term accounted for a high percentage of the total variation (57%–69%) of all ring widths and density components (Table 4). This result suggests that other factors not considered in this analysis may have influenced the variation in these properties, such as tree age, diameter at breast height, stand density, tree origin (sexual or vegetative), competition from other undergrowth in the same stand, and climatic conditions (annual growing degree-days and seasonal precipitation) [22,27,60]. The competition between trees in the same stand could significantly affect xylem development due to differences in crown structure and canopy position [72]. The topology of the xylem network could also affect water transport and EWC growth [69]. Under adverse conditions (such as drought), EWC tended to sectorize its radial architecture to almost isolate the hydraulic pathways completely to different parts of the stem and crown, thereby increasing the odds of survival but significantly decreasing its growth rate [52,53]. The hydraulic sectoriality in branches, stems, and taproots may be regarded as an adaptive trait to cope with environmental stress [54].
3.4. Relationships of Wood Density and Ring Width with Ecological Factors and Their Variation along a South–North Latitudinal Gradient in Abitibi Témiscamingue
As shown in Figure 5a, the RD increased along a South–North latitudinal gradient, with increasing latitude. The correlation coefficients between RD and RW and ecological factors are shown in Table 5. Most correlations were statistically significant (p < 0.001). RD correlated negatively with average temperature and annual precipitation and positively with latitude (Table 5). RD ranges from 346 to 379 kg m−3 from the southeast, central, northeast and northwest locations in Abitibi-Témiscamingue. LWD and EWD increased as well with the increase in latitude.
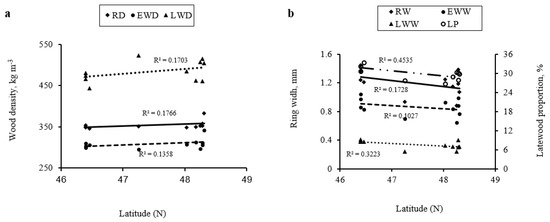
Figure 5.
Variations of (a) average ring density (RD, kg m−3), earlywood density (EWD, kg m−3), latewood density (LWD, kg m−3), (b) annual ring width (RW, mm), earlywood width (EWW, mm), latewood width (LWW, mm), and latewood proportion (LP, %) along a South–North latitudinal gradient in Abitibi Témiscamingue, Québec, Canada.
Table 5.
Correlation coefficients between annual ring density (RD) and annual ring width (RW) and ecological factors in Thuja occidentalis.
The between-tree variation in RD is more important among Abitibi sites (CV = 17%) compared to that in Témiscamingue sites (CV = 12%) (Table 2). The RD ranged from 351 to 379 kg m−3 in Abitibi sites and 348 to 354 kg m−3 in Témiscamingue sites. These values were higher compared to that reported in Wisconsin locations in this species [29], where the RD ranged from 314 to 324 kg m−3 from northeast, northwest, southeast, and southwest locations. The RD of trees from the central region was lower and averaged 281 kg m−3.
In the Témiscamingue region, the average RD of the different sites was not statistically significant. In contrast, in the Abitibi region, there were significant differences between the average RD of the studied sites (Table 2). The low variability in the Témiscamingue sites across the region could be explained by similar climatic, soil, and topological conditions (Table 1). These results reflect that the RD largely depends on ecological and environmental site conditions and supports previous findings on Pinus nigra [19] and Quercus petraea Liebl [18]. The climatic characteristics of each region appear to be the main factors affecting the RD components in T. occidentalis [27]. Mean annual temperatures and annual precipitation varied from 1.39 to 1.95 °C and 840 to 892 mm, respectively, in the Abitibi region (Table 1). In the Témiscamingue region, these values ranged from 2.33 to 3.63 °C and 874 to 932 mm, respectively. According to Housset et al. [27], annual growing degree days (GDD) and seasonal precipitation significantly explained the inter-site variability of EWC growth and, consequently, the wood density. The inter-annual growth variations in this species were positively correlated with spring temperature across the same study area [27]. In contrast, radial growth was negatively correlated with warm summer temperatures and positively correlated with precipitation in August. However, excess precipitation during autumn (October) negatively correlated with growth in the Témiscamingue region. Sites with greater annual GDD appeared slightly more sensitive to summer moisture stress. In addition, the site’s hydrological regime and tree size influenced the species’ response to summer thermal stress and moisture variability. Archambault, S., and Bergeron, Y. [28] reported that precipitation in June and low temperature in June or July negatively influenced growth.
RD showed the highest values in the northwest locations in Abitibi-Témiscamingue (Figure 5 and Table 2). Site A1 showed the highest value (379 kg m−3), followed by sites A5 (361 kg m−3) and A6 (360 kg m−3); however, site T4 showed the lowest (348 kg m−3). The difference in RD between site A1 and almost all other sites, except sites A5 and A6, is statistically significant (Table 2). At site A1, where the value of RD was the highest, the soil is sandy, and EWC is associated with spruce (Table 1), which seems to influence the RD positively [3]. Cedar is an important component of understory and mid-story strata. In contrast, spruce has a faster growth rate, promoting canopy openness and thus cedar growth without understory associates’ competition, with could positively affect the cambial activity and, consequently, wood quality [3]. Site A2 showed a relatively lower RD value (Table 2). Multiple factors, such as climate change (drought stress) and fire suppression, limit growth and contribute to EWC’s decline in the same study area [27].
In contrast to RD, the RW decreased along a South–North latitudinal gradient (Figure 5b). The RW is negatively and significantly (p < 0.001) correlated to the latitude (r = −0.12) and the longitude (r = −0.12) (Table 5). As temperature and precipitation decreased with increasing latitude and longitude, their relationships were strongly and negatively correlated (Table 5). For example, latitude accounted for 82% of the average temperature variation and 50% of the annual precipitation variation (Table 5). Decreasing temperature and precipitation by increasing latitude and longitude could negatively influence the RW and, consequently, the diameter at breast height and tree height (Table 5). Accordingly, the latitude accounted for about 78% of the diameter at breast height and tree height variations. The relationship between longitude and tree height was negative and more important (r = −0.80) compared to that between longitude and diameter at breast height (r = −0.50).
RW was higher in the Témiscamingue sites compared to the Abitibi sites (Table 2). Site T3 showed the highest RW value (1.43 mm), followed by site T2 (1.37 mm). However, site A6, located in the northwest and where the annual temperature and the annual precipitation are lower (Table 1), showed the lowest value (Table 2). Site T5, where the growing conditions (Table 1) are good (high temperature and precipitation values), showed a lower RW (1.09 mm) (Table 1) compared to other sites in the Témiscamingue region. This site is very wet and has a high moisture content (37%) [30]. This could negatively affect the tree growth and could explain the low RW. According to Johnston [3], EWC did not develop well on extremely wet or dry sites. Housset et al. [27] also reported that autumn and spring precipitation negatively impacted growth, especially in wet soil stands. Cedar growth is affected by seasonal variability in precipitation and local soil hydrological conditions. The link between growth and soil hydrology depends on C/N ratio in the upper 50 cm. Sites with a high C/N ratio could have their soils saturated with water for longer periods reducing the mineralization of organic matter, thus increasing the risk of root asphyxia. A secondary constraint of wet sites is that the spring soil warm-up is delayed, which consequently slows growth initiation and could retard soil frost thawing, limiting EWC growth. The paradoxical shift from an excess water constraint during the spring to drought stress in the summer can be attributed to the superficial root system that cedar develops under hydric edaphic conditions [65]. During the summer lowering of the water table, the upper layer of organic soils becomes dry, and the cedar’s shallow roots cannot supply water, limiting the growth.
4. Conclusions
This study investigated the regional, site, inter-tree, and radial variations in ring density and ring width components in Thuja occidentalis in the Québec Boreal Forest. It reinforces the arguments that accounting for cambial age and inter-tree variability should be considered by studying the relationship between the site and the ring density and, for the first time, provides information about the interregional variation as related to cambial age of wood density properties. The average ring density in trees grown at Abitibi-Témiscamingue was 356 kg m−3, with a small variation between earlywood and latewood (167 kg m−3), which reflects a relatively uniform wood. The within-tree distribution of latewood was relatively uniform, accounting for over 31% of the wood.
Ring density and width components showed higher variation with cambial age between sites and among trees within the same site. However, the largest variation is due to tree-specific conditions. The ring density components showed higher variation with cambial age than ring widths. The variation due to cambial age was more important in juvenile (wood formed in the early stages of tree growth) than in mature wood. In contrast, the ring widths showed a considerably larger tree-to-tree variation within the same site than ring density components. The tree-to-tree variation is more important in earlywood width than in latewood width. The site effect on ring density components was much less important than ring widths and depended on wood type. The variation due to the site was considerable on latewood width. The regional variations in ring width and ring density components in the Abitibi-Témiscamingue indicate that trees grown in the Abitibi region have higher ring density and lower ring width than those grown in the Témiscamingue region, mainly linked to the climatic characteristics of each area. The variation due to region was considerable on latewood width. It was more significant in juvenile than in mature wood, mainly linked to the variation of compression wood occurrence in Thuja occidentalis, associated with soils and environmental characteristics of each region.
The overall wood density characteristics of T. occidentalis stems, regarding their low variation between sites, among trees, and within the tree (between latewood and earlywood), are advantageous factors for the utilization of this wood for quality wood products that require wood uniformity, such as veneer peeling and slicing are an excellent end-use where a high degree of uniformity is very beneficial.
The regional and site variations of growth and wood density parameters of T. occidentalis suggested a complex response to ecological factors. Additional research is needed considering tree age, diameter at breast height, stand density, tree origin, anatomical characteristics, the topology of the xylem network, and soil hydrology to understand the variation of T. occidentalis wood quality as related to site and region characteristics. Additional research is also needed on the cedar’s genetic variation and adaptation to site characteristics.
Author Contributions
Conceptualization, A.K., B.B. and Y.B.; methodology, B.B.; software, B.B.; validation, B.B. and A.K.; formal analysis, B.B.; investigation, B.B.; resources, B.B. and A.K.; data curation, B.B.; writing—original draft preparation, B.B.; writing—review and editing, B.B., A.K. and Y.B.; visualization, B.B.; supervision, A.K. and Y.B.; project administration, A.K.; funding acquisition, A.K. and Y.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Canada Research Chair Program, grant number 512364…, the Ministère des Ressources Naturelles et de la Faune, Quebec (MRNF), and the NSERC-UQAT-UQAM Industrial chair on sustainable forest management.
Data Availability Statement
The data generated within the framework of this work are available upon request from the first author (B.B.).
Acknowledgments
The authors thank Veronique Paul for her insights on cedar ecology and her valuable help in the field sites. Melanie Desrochers for her help in localizing the studied sites and Igor Drobyshev for the climate data. The authors also thank Gilles Villeneuve for technical assistance.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the manuscript, or in the decision to publish the results.
References
- Koubaa, A.; Zhang, S.Y. White Cedar, Thuja occidentalis L. In Softwoods of Eastern Canada. Their Sylvics, Characteristics, Manufacturing and End-Use; Koubaa, A., Zhang, S.Y., Eds.; Special Publication, SP-526E, Chapter 11; FPInnovations: Québec, QC, Canada, 2008; 18p. [Google Scholar]
- Taylor, A.M.; Gartner, B.L.; Morrell, J.J. Heartwood formation and natural durability—A review. Wood Fiber. Sci. 2002, 34, 587–611. [Google Scholar]
- Johnston, W.F. Thuja occidentalis L. Northern white-cedar. In Silvics of North America; Forest Service, Agricultural Handbook; Volume 1: Conifers; Burns, R.M., Honkala, B.H., Eds.; United States Department of Agriculture: Washington, DC, USA, 1990; pp. 580–589. [Google Scholar]
- Behr, E.A. Special physical and chemical properties of northern white cedar. In Proceedings of the National Northern White Cedar Conference; No. 3-76; Michigan State University Publication: East Lansing, MI, USA, 1976; pp. 11–15. [Google Scholar]
- Wan, H.; Wang, X.M.; Yang, D.Q. Utilizing eastern white cedar to improve the resistance of strand boards to mold and decay fungi. For. Prod. J. 2007, 57, 54–59. [Google Scholar]
- Behr, E.A. Decay and termite resistance of medium-density fiberboards made from wood residue. For. Prod. J. 1972, 22, 48–51. [Google Scholar]
- Haataja, B.A.; Laks, P.E. Properties of flakeboard made from northern white cedar. For. Prod. J. 1995, 45, 68–70. [Google Scholar]
- Panshin, A.J.; De Zeeuw, C. Textbook of Wood Technology; McGraw-Hill Book Co.: New York, NY, USA, 1980; p. 772. [Google Scholar]
- Zobel, B.J.; Van Buijtenen, J.P. Wood Variation: Its Causes and Control; Springer: Berlin, Germany, 1989; p. 363. [Google Scholar]
- Zhang, S.Y.; Nepveu, G.; Owoundi, R.E. Intratree and intertree variation in selected wood quality characteristics of European oak (Quercus petraea and Quercus robur). Can. J. For. Res. 1994, 24, 1818–1823. [Google Scholar] [CrossRef]
- Koga, S.; Zhang, S.Y. Inter-tree and intra-tree variations in ring width and wood density components in balsam fir [Abies balsamea]. Wood Sci. Technol. 2004, 38, 149–162. [Google Scholar] [CrossRef]
- Guller, B.; Isik, K.; Cetinay, S. Variations in the radial growth and wood density components in relation to cambial age in 30-year-old Pinus brutia Ten. at two test sites. Trees 2012, 26, 975–986. [Google Scholar] [CrossRef]
- Barbour, J. Wood formation and properties. In Wood Quality, in Encyclopedia of Forest Sciences; Jeffery, B., Ed.; Elsevier: Oxford, UK, 2004; pp. 1840–1846. [Google Scholar]
- Bao, F.C.; Jiang, Z.H.; Jiang, X.M.; Lu, X.X.; Luo, X.Q.; Zhang, S.Y. Differences in wood properties between juvenile wood and mature wood in 10 species grown in China. Wood Sci. Technol. 2001, 35, 363–375. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Morgenstern, E.K. Genetic variation and inheritance of wood density in black spruce (Picea mariana) and its relationship with growth: Implications for tree breeding. Wood Sci. Technol. 1996, 30, 63–75. [Google Scholar] [CrossRef]
- Chave, J.; Muller-Landau, H.C.; Baker, T.R.; Easdale, T.A.; Steege, H.; Webb, C.O. Regional and phylogenetic variation of wood density across 2456 neotropical tree species. Ecol. Appl. 2006, 16, 2356–2367. [Google Scholar] [CrossRef] [PubMed]
- Savva, Y.; Koubaa, A.; Tremblay, F.; Bergeron, Y. Effects of radial growth, tree age, climate, and seed origin on wood density of diverse jack pine populations. Trees-Struct. Funct. 2010, 24, 53–65. [Google Scholar] [CrossRef]
- Bergès, L.; Nepveu, G.; Franc, A. Effects of ecological factors on radial growth and wood density components of sessile oak (Quercus petraea Liebl.) in Northern France. For. Ecol. Manag. 2008, 255, 567–579. [Google Scholar] [CrossRef]
- Oliva, A.G.; Merino, V.B.; Seco, J.I.F.G.; Garcia, M.C.; Prieto, E.H. Effect of growth conditions on wood density of Spanish Pinus nigra. Wood Sci. Technol. 2006, 40, 190–204. [Google Scholar] [CrossRef]
- DeRose, R.J.; Seymour, R.S. The effect of site quality on growth efficiency of upper crown class Picea rubens and Abies balsamea in Maine, USA. Can. J. For. Res. 2009, 39, 777–784. [Google Scholar] [CrossRef]
- Muller-Landau, H.C. Interspecific and inter-site variation in wood specific gravity of tropical trees. Biotropica 2004, 36, 20–32. [Google Scholar] [CrossRef]
- Ackermann, F. Relationship between forest site and intra-ring wood density components for the pedunculate oak (Quercus robur L.) of southwestern France. Ann. For. Sci. 1995, 52, 635–652. [Google Scholar] [CrossRef]
- Guilley, E.; Hervé, J.C.; Nepveu, G. The influence of site quality, silviculture and region on wood density mixed model in Quercus petraea Liebl. For. Ecol. Manag. 2004, 189, 111–121. [Google Scholar] [CrossRef]
- Knapic, S.; Louzada, J.L.; Pereira, H. Variation in wood density components within and between Quercus faginea trees. Can. J. For. Res. 2011, 41, 1212–1219. [Google Scholar] [CrossRef]
- Tardif, J.; Stevenson, D. Radial growth-climate association of Thuja occidentalis L. at the northwestern limit of its distribution, Manitoba, Canada. Dendrochronologia 2001, 19, 179–187. [Google Scholar]
- Tardif, J.; Bergeron, Y. Comparative dendroclimatological analysis of two black ash and two white cedar populations from contrasting sites in the Lake Duparquet region, northwestern Quebec. Can. J. For. Res. 1997, 27, 108–116. [Google Scholar] [CrossRef]
- Housset, J.M.; Girardin, M.P.; Baconnet, M.; Carcaillet, C.; Bergeron, Y. Unexpected warming-induced growth decline in Thuja occidentalis at its northern limits in North America. J. Biogeogr. 2015, 42, 1233–1245. [Google Scholar] [CrossRef]
- Archambault, S.; Bergeron, Y. An 802-year tree-ring chronology from the Quebec boreal forest. Can. J. For. Res. 1992, 22, 674–682. [Google Scholar] [CrossRef]
- Maeglin, R.R. Wisconsin Wood Density Suarvey; Forest Products Lab: Madison, WI, USA, 1973. [Google Scholar]
- Bouslimi, B.; Koubaa, A.; Bergeron, Y. Anatomical properties in Thuja occidentalis: Variation and relationship to biological processes. IAWA 2014, 35, 363–384. [Google Scholar] [CrossRef]
- Xu, H.; Tremblay, F.; Bergeron, Y.; Paul, V.; Chen, C. Genetic consequences of fragmentation in arbor vitae, eastern white cedar (Thuja occidentalis L.), toward the northern limit of its distribution range. Ecol. Evol. 2012, 2, 2506–2520. [Google Scholar] [CrossRef]
- Paul, V. Les Facteurs Ecologiques Limitant la Répartition Nordique du Thuja de L’est (Thuja occidentalis L.); Université du Québec en Abitibi-Témiscamingue: Rouyn-Noranda, QC, Canada, 2011; p. 84. [Google Scholar]
- Régnière, J.; St-Amant, R. Stochastic simulation of daily air temperature and precipitation from monthly normals in North America north of Mexico. Int. J. Biometeorol. 2007. 51, 415–430. [CrossRef]
- Bergeron, Y.; Gauthier, S.; Flannigan, M.; Kafka, V. Fire regimes at the transition between mixedwood and coniferous boreal forest in northwestern Quebec. Ecology 2004, 85, 1916–1932. [Google Scholar] [CrossRef]
- Bouslimi, B.; Koubaa, A.; Bergeron, Y. Variation of brown rot decay in eastern white cedar (Thuja occidentalis L.). BioResources 2013, 8, 4735–4755. [Google Scholar] [CrossRef]
- Koubaa, A.; Zhang, S.Y.; Makni, S. Defining the transition from earlywood to latewood in black spruce based on intra-ring wood density profiles from X-ray densitometry. Ann. For. Sci. 2002, 59, 511–518. [Google Scholar] [CrossRef]
- Singleton, R.; DeBell, D.S.; Gartner, B.L. Effect of extraction on wood density of western hemlock (Tsuga heterophylla (Raf.) Sarg.). Wood Fiber Sci. 2003, 35, 363–369. [Google Scholar]
- Grabner, M.; Wimmer, R.; Gierlinger, N.; Evans, R.; Downes, G. Heartwood extractives in larch and effects on X-ray densitometry. Can. J. For. Res. 2005, 35, 2781–2786. [Google Scholar] [CrossRef]
- Bouslimi, B.; Koubaa, A.; Bergeron, Y. Intra-Ring Variations and Interrelationships for Selected Wood Anatomical and Physical Properties of Thuja occidentalis L. Forests 2019, 10, 339. [Google Scholar] [CrossRef]
- Smith, D. Maximum Moisture Content Method for Determining Specific Gravity of Small Wood Samples; Forest Products Laboratory Report 2014; Forest Products Laboratory: Madison, WI, USA, 1954. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–75. [Google Scholar]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Schabenberber, O. SAS for Mixed Models, 2nd ed.; SAS Institute, Inc.: Cary, NC, USA, 2006; p. 814. [Google Scholar]
- SAS Institute Inc. Statistical Analysis Software. Users’ Guide Statistics Version 9.4; SAS Institute Inc.: Cary, NC, USA, 2008. [Google Scholar]
- Park, Y.I.; Koubaa, A.; Brais, S.; Mazerolle, M.J. Effects of cambial age and stem height on wood density and growth of jack pine grown in boreal stands. Wood Fiber Sci. 2009, 41, 346–358. [Google Scholar]
- Koubaa, A.; Zhang, S.Y.; Isabel, N.; Beaulieu, J.; Bousquet, J. Phenotypic correlations between juvenile-mature wood density and growth in black spruce. Wood Fiber Sci. 2000, 32, 61–71. [Google Scholar]
- Jozsa, L.A.; Middleton, G.R. A Discussion of Wood Quality Attributes and Their Practical Implications; No. SP-34; Forintek Canada Corp.: Ottawa, ON, Canada, 1994; p. 42. [Google Scholar]
- Larson, P.R.; Kretschmann, D.E.; Clark, A., III; Isebrands, J.G. Formation and Properties of Juvenile Wood in Southern Pines: A Synopsis; USDA Forest Service, Forest Products Laboratory: Madison, WI, USA, 2001; p. 42. [Google Scholar]
- Alteyrac, J.; Cloutier, A.; Zhang, S.Y. Characterization of juvenile wood to mature wood transition age in black spruce (Picea mariana (Mill.) B.S.P.) at different stand densities and sampling heights. Wood Sci. Technol. 2006, 40, 124–138. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Simpson, D.; Morgenstern, E.K. Variation in the relationship of wood density with growth in 40 black spruce (Picea mariana) families grown in New Brunswick. Wood Fiber Sci. 1996, 28, 91–99. [Google Scholar]
- Koga, S.; Zhang, S.Y. Relationships between wood density and annual growth rate components in balsam fir (Abies balsamea). Wood Fiber Sci. 2002, 34, 146–157. [Google Scholar]
- Bergeron, Y. Species and stand dynamics in the mixed woods of Quebec’s southern boreal forest. Ecology 2000, 81, 1500–1516. [Google Scholar] [CrossRef]
- Larson, D.W.; Doubt, J.; Matthes-Sears, U. Radially Sectored Hydraulic Pathways in the Xylem of Thuja occidentalis as Revealed by the Use of Dyes. Int. J. Plant Sci. 1994, 155, 569–582. [Google Scholar] [CrossRef]
- Matthes-Sears, U.; Nash, C.H.; Larson, D.W. Constrained growth of trees in a hostile environment: The role of water and nutrient availability for Thuja occidentalis on cliff faces. Int. J. Plant Sci. 1995, 156, 311–319. [Google Scholar] [CrossRef]
- Matthes-Sears, U.; Larson, D.W. Growth and physiology of Thuja occidentalis L. from cliffs and swamps: Is variation habitat or site-specific? Bot. Gaz. 1991, 152, 500–508. [Google Scholar] [CrossRef]
- Koubaa, A.; Isabel, N.; Zhang, S.Y.; Beaulieu, J.; Bousquet, J. Transition from juvenile to mature wood in black spruce (Picea mariana (Mill.) B.S.P.). Wood Fiber Sci. 2005, 37, 445–455. [Google Scholar]
- Paul, V.; Bergeron, Y.; Tremblay, F. Does climate control the northern range limit of eastern white cedar (Thuja occidentalis L.)? Plant Ecol. 2014, 215, 181–194. [Google Scholar] [CrossRef]
- Bergeron, Y. Les conséquences des changements climatiques sur la fréquence des feux et la composition forestière au sud-ouest de la forêt boréale québécoise. Géogr. Phys. Quat. 1998, 52, 167–174. [Google Scholar] [CrossRef][Green Version]
- Schulz, K. Northern White-Cedar Stand Structure, Composition, and Ecophysiology: Implications for Management and Climate Change Resilience. Electronic Theses and Dissertations. 3694. p. 90. Available online: https://digitalcommons.library.umaine.edu/etd/3694 (accessed on 4 October 2022).
- Fromm, J. Cellular Aspects of Wood Formation; Springer: Berlin/Heidelberg, Germany, 2013; p. 264. [Google Scholar]
- Larocque, G.R. Importance of different climatic parameters on growth and wood formation of red pine (Pinus resinosa Ait) in Ontario (Canada). Ann. Des. Sci. For. 1997, 54, 51–63. [Google Scholar] [CrossRef]
- Fei, B.H.; Ruan, X.G. Effects of temperature and precipitation on tree-ring and wood density of Ginkgo in Beijing. For. Res. 2001, 14, 176–180. [Google Scholar]
- Gindl, W.; Grabner, M.; Wimmer, R. The influence of temperature on latewood lignin content in treeline Norway spruce compared with maximum density and ring width. Trees 2000, 14, 409–414. [Google Scholar] [CrossRef]
- Zhang, S.Y. Variations and correlations of various ring width and ring density features in European oak: Implications in dendroclimatology. Wood Sci. Technol. 1997, 31, 63–72. [Google Scholar] [CrossRef]
- Plomion, C.; Leprovost, G.; Stokes, A. Wood formation in trees. Plant Physiol. 2001, 127, 1513–1523. [Google Scholar] [CrossRef]
- Hofmeyer, P.V.; Kenefic, L.S.; Seymour, R.S. Northern white-cedar ecology and silviculture in the northeastern United States and southeastern Canada: A synthesis of knowledge. North. J. Appl. For. 2009, 26, 21–27. [Google Scholar] [CrossRef]
- Rooney, T.P.; Solheim, S.L.; Waller, D.M. Factors affecting the regeneration of northern white cedar in lowland forests of the Upper Great Lakes region, USA. For. Ecol. Manag. 2002, 163, 119–130. [Google Scholar] [CrossRef]
- Rosner, S. Hydraulic and biomechanical optimization in Norway spruce trunkwood—A review. IAWA J. 2013, 34, 365–390. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Pockman, W.T.; Davis, S.D.; McCulloh, K.A. Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 2001, 126, 457–461. [Google Scholar] [CrossRef]
- Tyree, M.T.; Ewers, F.W. The hydraulic architecture of trees and other woody plants. New Phytol. 1991, 119, 345–360. [Google Scholar] [CrossRef]
- Larson, D.W.; Matthes-Sears, U.; Kelly, P.E. Cambial Dieback and Partial Shoot Mortality in Cliff-Face Thuja occidentalis: Evidence for Sectored Radial Architecture. Int. J. Plant Sci. 1993, 154, 496–505. [Google Scholar] [CrossRef]
- Fujiwara, S.; Yang, K. The relationship between cell length and ring width and circumferential growth rate in five Canadian species. IAWA J. 2000, 21, 335–346. [Google Scholar] [CrossRef]
- Hofmeyer, P.V.; Kenefic, L.S.; Seymour, R.S. Historical stem development of northern White-cedar (Thuja occidentalis L.) in Maine. North. J. Appl. For. 2010, 27, 92–96. [Google Scholar] [CrossRef]
- Mansfield, S.D.; Parish, R.; Goudie, J.W.; Kang, K.Y.; Ott, P. The effects of crown ratio on the transition from juvenile to mature wood production in lodgepole pine in western Canada. Can. J. For. Res. 2007, 37, 1450–1459. [Google Scholar] [CrossRef]
- Kuprevicius, A.P. Quantifying the Influence of Crown Size on Mechanical Wood Properties in White Spruce (Picea glauca); University of Toronto: Toronto, ON, Canada, 2011; p. 72. [Google Scholar]
- Briand, C.H.; Posluszny, U.; Larson, D.W. Influence of age and growth rate on radial anatomy of annual rings of Thuja occidentalis L. (eastern white cedar). Int. J. Plant Sci. 1993, 154, 406–411. [Google Scholar] [CrossRef]
- Koubaa, A.; Zhang, S.Y. Jack pine (Pinus banksiana Lamb). In Softwoods of Eastern Canada. Their Sylvics, Characteristics, Manufacturing and End-Uses; Koubaa, A., Zhang, S.Y., Eds.; Special Publication, SP-526F, Chapter 4; FPInnovations: Quebec, QC, Canada, 2008; 35p. [Google Scholar]
- Koubaa, A.; Zhang, S.Y. White spruce (Picea glauca (Moench) Voss). In Softwoods of Eastern Canada. Their Sylvics, Characteristics, Manufacturing and End-Uses; Zhang, S.Y., Koubaa, A., Eds.; Special Publication, SP-526F, Chapter 2; FPInnovations: Quebec, QC, Canada, 2008; 33p. [Google Scholar]
- Gryc, V.; Vavrčík, H.; Horn, K. Density of juvenile and mature wood of selected coniferous species. J. For. Sci. 2011, 57, 123–130. [Google Scholar] [CrossRef]
- Knapic, S.; Louzada, J.L.; Leal, S.; Pereira, H. Within-tree and between-tree variation of wood density components in cork oak trees in two sites in Portugal. Forestry 2008, 81, 465–473. [Google Scholar] [CrossRef]
- Bouriaud, O.; Leban, J.-M.; Bert, D.; Deleuze, C. Intra-annual variations in climate influence growth and wood density of Norway spruce. Tree Physiol. 2005, 25, 651–660. [Google Scholar] [CrossRef]
- Spicer, R.; Gartner, B.L. The effects of cambial age and position within the stem on specific conductivity in Douglas-fir (Pseudotsuga menziesii) sapwood. Trees-Struct. Funct. 2001, 15, 222–229. [Google Scholar] [CrossRef]
- Hofmeyer, P.V.; Kenefic, L.S.; Seymour, R.S.; Brissette, J.C. Growth Comparison of Northern White-Cedar to Balsam Fir and Red Spruce by Site Class; Forest Science; University of Maine: Orono, ME, USA, 2006; p. 04469. [Google Scholar]
- Gartner, B.L.; Robbins, J.M.; Newton, M. Effects of pruning on wood density and tracheid length in young Douglas-fir. Wood Fiber Sci. 2005, 37, 304–313. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).