
Fire Severity Controls Successional Pathways in a Fire-Affected Spruce Forest in Eastern Fennoscandia

 , ,
, ,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
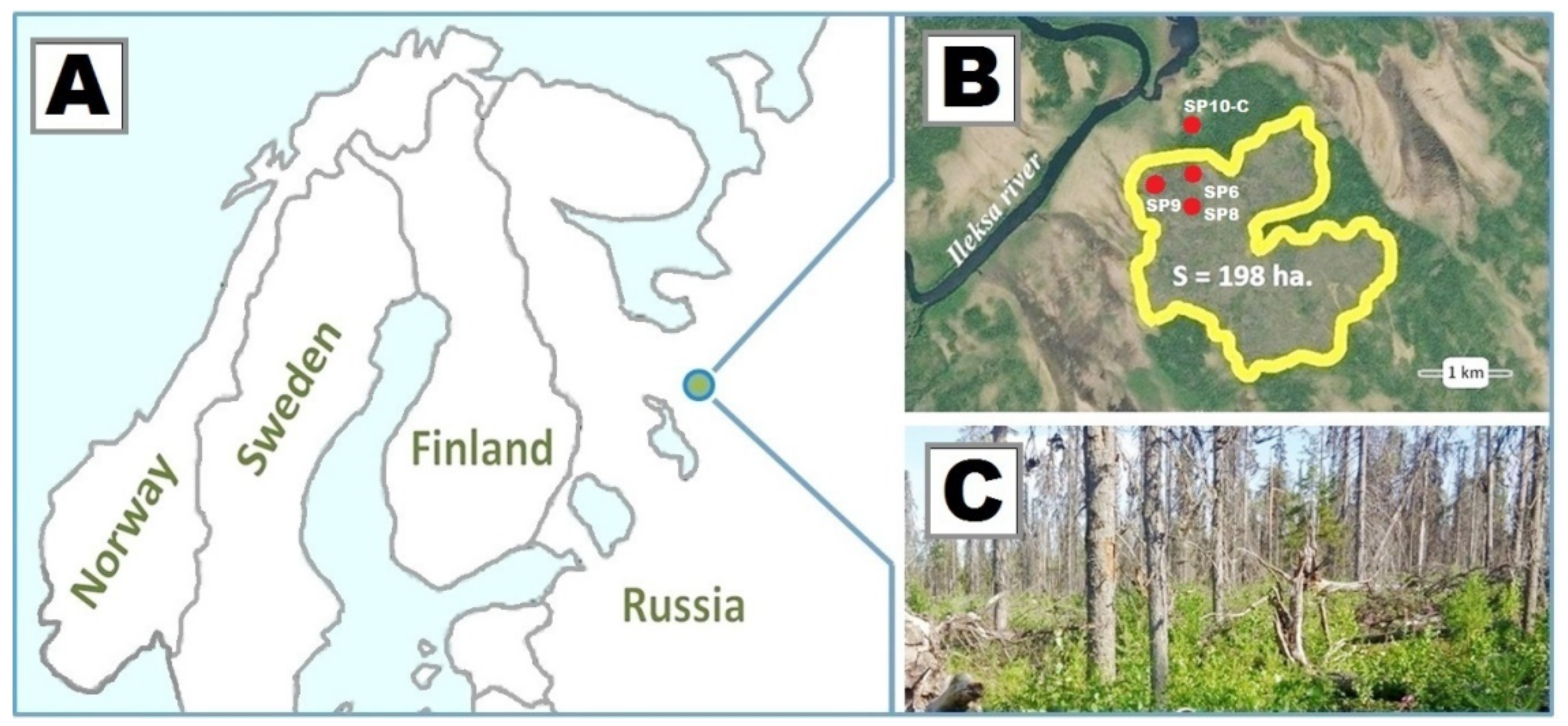
2.1. Study Area
2.2. Methods
3. Results
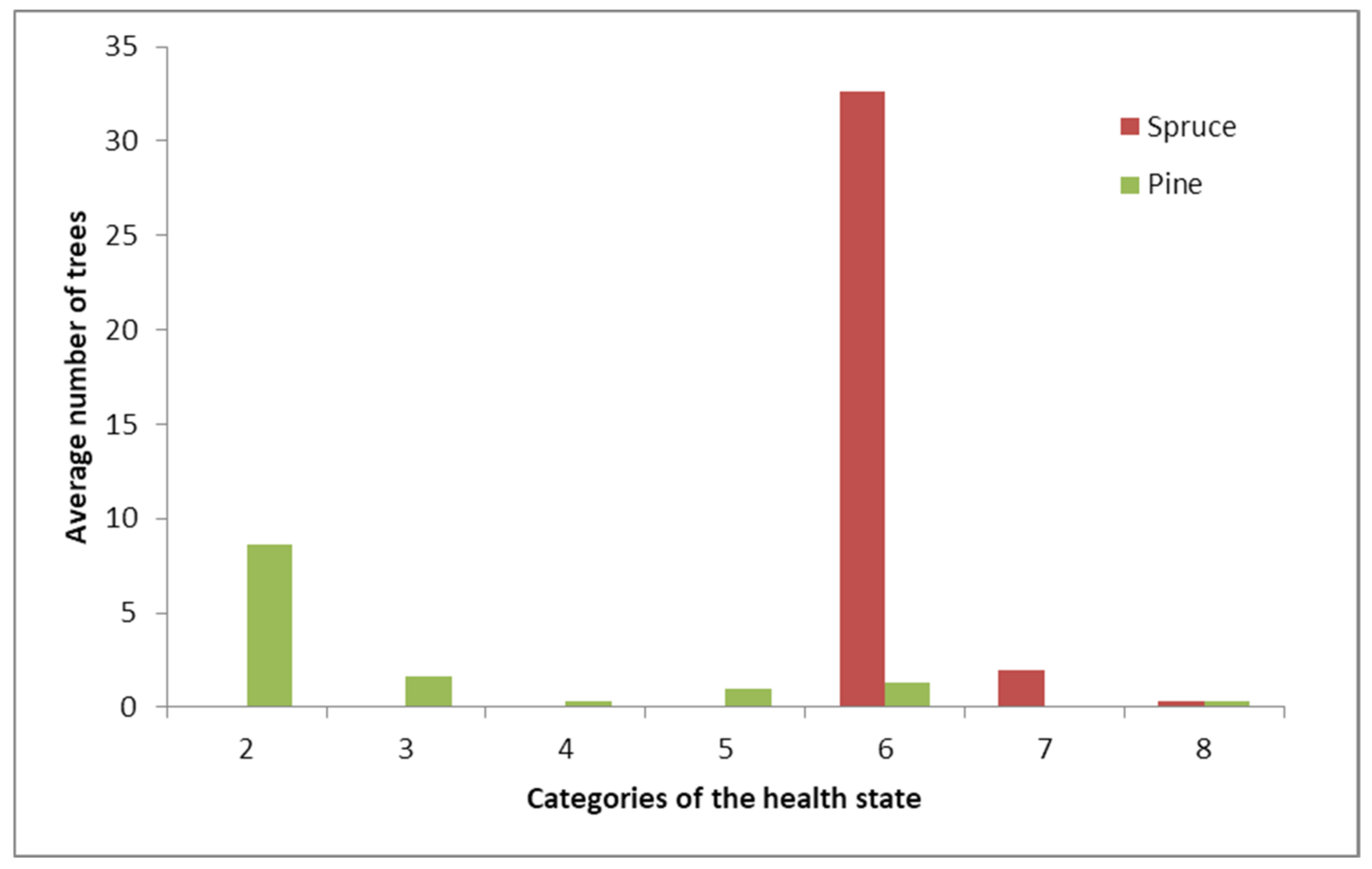
3.1. State of the Tree Layer after Surface Fire
3.2. Biotic Factors Influencing Tree Stand Die-Back after Surface Fires
3.3. Regeneration of the Tree-Shrub and Herb-Subshrub Layers
3.4. Diversity of Aphyllophoroid Fungi

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Log Species Identity | 2014 | Diameter, cm | Decay Class | 2020 | SP 10-C | ||
|---|---|---|---|---|---|---|---|---|
| SP 6 | SP 8 | SP 9 | ||||||
| Amphinema byssoides (Pers.) J. Erikss. | S | – | 8 | 4 | 1 | – | – | – |
| Amylocorticium suaveolens Parmasto | S | – | – | – | – | – | – | + |
| ** Amylocystis lapponica (Romell) Bondartsev et Singer | S | + | – | – | – | – | – | + |
| Antrodia sinuosa (Fr.) P. Karst. | S | – | 8 | 4 | 1 | 1 | – | – |
| * Asterodon ferruginosus Pat. | S | – | 18–20 | 3–4 | – | – | – | + |
| ** Byssomerulius albostramineus(Torrend) Hjortstam | P | – | – | – | – | – | – | + |
| Cerrena unicolor (Bull.) Murrill | B | – | 15 | 3 | 1 | – | – | + |
| ** Chaetodermella luna (Romell ex D.P. Rogers et H.S. Jacks.) Rauschert | P | – | – | – | – | – | – | + |
| Coltricia perennis (L.) Murrill | litter | – | – | – | – | – | – | + |
| Coniophora arida (Fr.) P. Karst. | S | – | – | – | – | – | – | + |
| C. olivacea (Fr) P. Karst. | S | – | 18 | 2 | 1 | – | – | + |
| ** Crustoderma dryinum (Berk. et M.A. Curtis) Parmasto | S | – | 24 | 2 | – | – | 1 | + |
| Cyanosporus caesius (Schrad.) McGinty | S | – | 8–17 | 2–4 | – | 1 | 1 | – |
| Daedalea xantha (Fr.) A. Roy et A.B. De | S | – | 10–70 | 2–4 | 1 | 2 | 1 | + |
| Daldinia concentrica (Bolton) Ces. et De Not. | B | + | – | – | – | – | – | – |
| Dichomitus squalens (P. Karst.) D.A. Reid | S | – | 20 | 1 | – | – | 1 | – |
| Diplomitoporus flavescens (Bres.) Domański | P | – | – | – | – | – | – | + |
| Fibrodontia brevidens (Pat.) Hjortstam et Ryvarden | S | – | 20 | 2 | – | – | – | + |
| Fomes fomentarius (L.) Fr. | B | + | 12–22 | 1–3 | 3 | 1 | 7 | + |
| Fomitiporia punctata (P. Karst.) Murrill | B | – | 12 | 2 | – | – | 1 | – |
| Fomitopsis betulina (Bull.) B.K. Cui, M.L. Hanet Y.C. Dai | B | + | 14–16 | 1–2 | – | 1 | 3 | + |
| F. pinicola (Sw.) P. Karst. | B, S | + | 11–26 | 1–3 | 11 | 6 | 7 | + |
| Gloeophyllum odoratum (Wulfen) Imazeki | S | – | 28 | 4 | – | 1 | – | – |
| G. sepiarium (Wulfen) P. Karst. | S | + | 10–26 | 1–4 | 4 | 12 | 15 | + |
| * Hericium coralloides (Scop.) Pers. | A, B | – | 18 | 2 | 1 | – | – | + |
| Hydnomerulius pinastri (Fr.) Jarosch et Besl | S | – | – | – | – | – | – | + |
| Hyphoderma setigerum (Fr.) Donk | S | – | 5 | 1 | 1 | – | – | – |
| Incrustoporia chrysella (Niemelä) Zmitr. | S | – | – | – | – | – | – | + |
| ** Inonotus leporinus (Fr.) Gilb. et Ryvarden | S | – | – | – | – | – | – | + |
| I. obliquus (Fr.) Pilát | B | – | – | – | – | – | – | + |
| ** Junghuhnia collabens (Fr.) Ryvarden | S | – | – | – | – | – | – | + |
| Mycoacia livida (Pers.) Zmitr. | S | – | – | – | – | – | – | + |
| Neoantrodia serialis (Fr.) Audet | S | – | – | – | – | – | – | + |
| Peniophora cinerea (Pers.) Cooke | B | – | – | – | – | – | – | + |
| P. pithya (Pers.) J. Erikss. | S | – | 12 | 2–3 | – | – | 1 | – |
| P. septentrionalis Laurila | S | – | 8–18 | 2–4 | 1 | 1 | 4 | – |
| Phanerochaete sordida (P. Karst.) J. Erikss. et Ryvarden | S | – | – | – | – | – | – | + |
| * Phellinidium ferrugineofuscum (P. Karst.) Fiasson et Niemelä | S | + | 4–26 | 2–4 | 4 | 5 | 1 | + |
| * Phellinus chrysoloma (Fr.) Donk | S | + | 12–18 | 2–3 | – | – | 4 | + |
| P. igniarius (L.) Quél. | W | – | – | – | – | – | – | + |
| P. laevigatus (P. Karst.) Bourdot et Galzin | B | – | 10 | 2 | – | – | 1 | – |
| P. lundellii Niemelä | B | – | – | – | – | – | – | + |
| P. nigricans (Fr.) P. Karst. | B | – | 12–24 | 1–2 | 1 | 1 | 4 | – |
| P. tremulae (Bondartsev) Bondartsev et P.N. Borisov | A | – | – | – | – | – | – | + |
| * P. viticola (Schwein.) Donk | S | + | 4–70 | 2 | 2 | – | – | + |
| ** Phellopilus nigrolimitatus (Romell) Niemelä, T. Wagner et M. Fisch. | S | – | – | – | – | – | 4 | + |
| Piloderma bicolor (Peck) Jülich | S | – | 6–18 | 2–4 | 1 | 1 | – | – |
| Porodaedalea pini (Brot.) Murrill | P | – | 32 | 1 | – | 1 | – | – |
| * Pycnoporellus fulgens (Fr.) Donk | S | – | 19–20 | 2 | 2 | – | – | + |
| Pycnoporus cinnabarinus (Jacq.) P. Karst. | B | – | 7–24 | 1–2 | 1 | 1 | 3 | – |
| * Rhodofomes roseus (Alb. et Schwein.) Kotl. et Pouzar | S | + | 5–22 | 2–4 | 4 | 3 | 1 | + |
| Scytinostroma odoratum (Fr.) Donk | S | – | 38 | 1 | 1 | – | – | – |
| Stereum sanguinolentum (Alb. et Schwein.) Fr. | S | – | – | – | – | – | – | + |
| Thelephora terrestris Ehrh. ex Fr. | S, litter | + | 6 | 2 | 1 | – | – | – |
| Thelephora wakefieldiae Zmitr., Shchepin, Volobuev et Myasnikov [= Tomentella sublilacina (Elliset Holw.) Wakef.] | S | – | 6 | 1 | 1 | – | – | – |
| Tomentella coerulea Höhn. et Litsch. | S | – | 12 | 4 | 1 | – | 1 | – |
| T. lapida (Pers.) Stalpers | S | – | – | – | 1 | – | – | – |
| Trametes ochracea (Pers.) Gilb. et Ryvarden | B | – | 16 | 2 | – | – | 1 | + |
| Trechispora sp. | S | – | – | – | – | – | – | + |
| Trichaptum abietinum (Pers. ex J.F. Gmel.) Ryvarden | S | + | 5–26 | 1–4 | 5 | 2 | 2 | + |
| T. biforme (Fr.) Ryvarden | B | – | 16–30 | 2–3 | 2 | – | 2 | – |
| T. fuscoviolaceum (Ehrenb.) Ryvarden | S | + | 8–32 | 2–3 | 4 | 5 | 2 | + |
| T. laricinum (P. Karst.) Ryvarden | P, S | – | 18–20 | 1–2 | – | 1 | 2 | + |
| Tubulicrinis borealis J. Erikss. | S | – | 12 | 4 | – | – | 1 | – |
| T. calothrix (Pat.) Donk | S | – | 12 | 4 | – | – | – | + |
| ** Tyromyces odorus (Sacc.) Zmitr. | S | – | – | – | – | – | – | + |
| Veluticeps abietina (Pers.) Hjortstam et Tellería | S | – | 70 | 2 | 1 | – | – | – |
| Xylodon asper (Fr.) Hjortstam et Ryvarden | S | – | 12 | 4 | – | – | – | + |
3.5. Soils 10 Years after Surface Fire
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dyrenkov, S.A. Structure and Dynamics of Taiga Spruce Forests; Nauka: Leningrad, Russia, 1984; p. 174. (In Russian) [Google Scholar]
- Bakhtin, A.A. Formation of deciduous-spruce forest stands on the burned areas of the Arkhangelsk region. In Flora of the North and Plant Resources of the European Part of the USSR; Arkhangelsk, Russia, 1997; pp. 81–83. (In Russian) [Google Scholar]
- Ryan, K.C. Dynamic interactions between forest structure and fire behavior in boreal ecosystems. Silva Fenn. 2002, 36, 13–39. [Google Scholar] [CrossRef]
- Lundqvist, L.; Nilson, K. Regeneration dynamics in an uneven-aged virgin Norway Spruce forest in northern Sweden. Scand. J. For. Res. 2007, 22, 304–309. [Google Scholar] [CrossRef]
- Cai, W.H.; Yang, J. High-severity fire reduces early successional boreal larch forest aboveground productivity by shifting stand density in north-eastern China. Int. J. Wildland Fire 2016, 25, 861–875. [Google Scholar] [CrossRef]
- Chambers, M.E.; Fornwalt, P.J.; Malone, S.L.; Battaglia, M.A. Patterns of conifer regeneration following high severity wildfire in ponderosa pine—dominated forests of the Colorado Front Range. For. Ecol. Manag. 2016, 378, 57–67. [Google Scholar] [CrossRef]
- Stavrova, N.I.; Gorshkov, V.V.; Katyutin, P.N.; Bakkal, I.J. The Structure of Northern Siberian Spruce–Scots Pine Forests at Different Stages of Post-Fire Succession. Forests 2020, 11, 558. [Google Scholar] [CrossRef]
- Foster, D.R. Vegetation development following fire in Picea mariana (black spruce) Pleurozium forests of south-eastern Labrador, Canada. J. Ecol. 1985, 73, 517–534. [Google Scholar] [CrossRef]
- Stickney, P.F. First Decade Plant Succession Following the Sundance Fire, Northern Idaho; General Technical Report INT-197; US Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1986; p. 26.
- Schimmel, J.; Granstrom, A. Fire severity and vegetation response in the boreal Swedish forest. Ecology 1996, 77, 1436–1450. [Google Scholar] [CrossRef]
- Turner, M.G.; Romme, W.H.; Gardner, R.H.; Hargrove, W.W. Effects of fire size and pattern on early succession in Yellowstone National Park. Ecol. Monogr. 1997, 67, 411–433. [Google Scholar] [CrossRef]
- Hart, B.T.; Burgman, M.; Grace, M.; Pollino, C.; Thomas, C.; Webb, J.A.; Allison, G.A.; Chapman, M.; Duivenvoorden, L.; Feehan, P.; et al. Ecological Risk Management Framework for the Irrigation Industry; Technical Report; Land and Water Australia: Canberra, Australia, 2005. [Google Scholar]
- Ruokolainen, L.; Salo, K. The effect of the fire intensity on vegetation succession on a sub-xeric heath during ten years after wildfire. Ann. Bot. Fenn. 2009, 46, 30–42. [Google Scholar] [CrossRef]
- Kovaleva, N.M.; Ivanova, G.A. Recovery of living ground vegetation at the initial stage of pyrogenic succession. Contemp. Probl. Ecol. 2013, 2, 203–213. (In Russian) [Google Scholar]
- Kovaleva, N.M. Effect of surface fires on ground layer in larch stands of the lower Angara region. Bot. Zhurn. 2014, 11, 1269–1277. (In Russian) [Google Scholar]
- Khapugin, A.A.; Vargot, E.V.; Chugunov, G.G. Vegetation recovery in fire-damaged forests: A case study at the southern boundary of the taiga zone. For. Stud. Metsanduslikud Uurim. 2016, 64, 39–50. [Google Scholar] [CrossRef]
- Ivanova, G.A.; Ivanov, V.A.; Kovaleva, N.M.; Konard, S.G.; Zhila, S.V.; Tarasov, P.A. Succession of vegetation after the high intensity fire in a pine forest with lichens. Contemp. Probl. Ecol. 2017, 17, 61–71. (In Russian) [Google Scholar] [CrossRef]
- Kuleshova, L.V.; Korotkov, V.N.; Potapova, N.A.; Evstigneev, O.I.; Kozlenko, A.B.; Rusanova, O.M. Comprehensive analysis of post-fire successions in the forests of the Kostomuksha Reserve. Bull. Mosc. Soc. Naturalists. Biol. Ser. 1996, 101, 3–15. (In Russian) [Google Scholar]
- Zyryanova, O.A.; Bugaenko, T.N. Species diversity of larch associations in the permafrost zone of Central Siberia and its post-fire transformation. Structural and functional organization and dynamics of forests. In Proceedings of the All-Russian Conference, Krasnoyarsk, Russia, 1–3 September 2004; pp. 301–303. (In Russian). [Google Scholar]
- Ivanova, G.A.; Perevoznikova, V.D.; Ivanov, V.A. Transformation of the lower tiers of forest vegetation after ground fires. Russ. J. For. Sci. 2002, 2, 30–35. (In Russian) [Google Scholar]
- Zyryanova, O.A.; Bugaenko, T.N.; Abaimov, A.P.; Bugaenko, N.N. Pyrogenic transformation of species diversity in the larch forests of the cryolithozone. In Forest Ecosystems of the Yenisey Meridian; Publishing House SO RAS: Novosibirsk, Russia, 2002; pp. 135–146. (In Russian) [Google Scholar]
- Isaeva, L.G.; Himich, Y.R.; Kostina, V.A. Diversity of spruce forests and aphyllophoroid fungi in the Murmansk region. In Coniferous Forests of Northern Latitudes—From Research to Environmentally Responsible Forestry; Kauhanen, H., Neshataev, V., Huhta, E., Vuopio, M., Eds.; Kopijyvä: Jyväskylä, Finland, 2009; pp. 49–60. (In Russian) [Google Scholar]
- Himich, Y.R. Polypore Fungi in the Process of Succession of Spruce Forests of the Murmansk Region. Ph.D. Thesis, Syktyvkar, Russia, 2011; p. 20. (In Russian). [Google Scholar]
- Suominen, M.; Junninen, K.; Heikkala, O.; Kouki, J. Combined effects of retention forestry and prescribed burning on polypore fungi. J. Appl. Ecol. 2015, 52, 1001–1008. [Google Scholar] [CrossRef]
- Komarova, T.A. Post-Fire Successions in the Forests of Southern Sikhote-Alin; Publishing House of the Far Eastern Scientific Center: Vladivostok, Russia, 1992; p. 222. (In Russian) [Google Scholar]
- Abaimov, A.P.; Prokushkin, S.G.; Zyryanova, O.A. Ecological and phytocenotic assessment of the impact of fires on the forests of the permafrost zone of Central Siberia. Contemp. Probl. Ecol. 1996, 1, 51–601. (In Russian) [Google Scholar]
- Kalinin, K.K. Vegetation cover successions on large burnt areas of the Middle Trans-Volga. Vestn. Mari State Univ. Ser. For. Ecology. Nat. Manag. 2008, 1, 19–28. (In Russian) [Google Scholar]
- Engelmark, O.; Hytteborn, H. Coniferous forests. Acta Phytogeogr. Suec. 1999, 84, 55–74. [Google Scholar]
- Oikonomakis, N.; Ganatsas, P. Land cover changes and forest succession trends in a site of Natura 2000 network (Elatia forest), in northern Greece. For. Ecol. Manag. 2012, 285, 153–163. [Google Scholar] [CrossRef]
- Christopoulou, A.; Mallinis, G.; Vassilakis, E.; Farangitakis, G.P.; Fyllas, N.M.; Kokkoris, G.D.; Arianoutsou, M. Assessing the impact of different landscape features on post-fire forest recovery with multitemporal remote sensing data: The case of Mount Taygetos (southern Greece). Int. J. Wildland Fire 2019, 28, 521–532. [Google Scholar] [CrossRef]
- Hjältén, J.; Dynesius, M.; Hekkala, A.-M.; Karlsson-Tiselius, A.; Löfroth, T.; Mugerwa-Pettersson, R. Saproxylic Insects and Fire. In Saproxylic Insects; Diversity, Ecology and Conservation, Zoological Monographs; Ulyshen, M.D., Ed.; Springer: Cham, Switzerland, 2018; pp. 669–691. [Google Scholar] [CrossRef]
- Kamdem, D.P.; Pizzi, A.; Jermannaud, A. Durability of heat-treated wood. Holz Roh Werkst 2002, 60, 1–6. [Google Scholar] [CrossRef]
- Weiland, J.J.; Guyonnet, R. Study of chemical modifications and fungi degradation of thermally modified wood using DRIFT spectroscopy. Holz Roh Werkst 2003, 61, 216–220. [Google Scholar] [CrossRef]
- Hakkou, M.; Pétrissans, M.; Gérardin, P.; Zoulalian, A. Investigation of the reasons for fungal durability of heat-treated beech wood. Polym. Degrad. Stabil. 2006, 91, 393–397. [Google Scholar] [CrossRef]
- Shorokhova, E.; Kapitsa, E.; Vanha-Majamaa, I. Decomposition of stumps in a chronosequence after clear-felling vs. clear-felling with prescribed burning in a southern boreal forest in Finland. For. Ecol. Manag. 2008, 255, 3606–3612. [Google Scholar] [CrossRef]
- Edman, M.; Eriksson, A.-M. Competitive outcomes between wood decaying fungi are altered in burnt wood. FEMS Microbiol. Ecol. 2016, 92, fiw068. [Google Scholar] [CrossRef]
- Eriksson, A.M.; Olsson, J.; Jonsson, B.G.; Toivonen, S.; Edman, M. Effects of restoration fire on dead wood heterogeneity and availability in three Pinus sylvestris forests in Sweden. Silva Fenn. 2013, 47, 954. [Google Scholar] [CrossRef]
- Van der Wal, A.; Ottosson, E.; de Boer, W. Neglected role of fungal community composition in explaining variation in wood decay rates. Ecology 2015, 96, 124–133. [Google Scholar] [CrossRef]
- Carlsson, F.; Edman, M.; Jonsson, B.G. Increased CO2 evolution caused by heat treatment in wood-decaying fungi. Mycol. Prog. 2017, 16, 513–519. [Google Scholar] [CrossRef]
- Godfree, R.C.; Knerr, N.; Encinas-Viso, F.; Albrecht, D.; Bush, D.; Christine Cargill, D.; Broadhurst, L.M. Implications of the 2019–2020 megafires for the biogeography and conservation of Australian vegetation. Nat. Commun. 2021, 12, 1023. [Google Scholar] [CrossRef]
- Stefanidis, S.; Alexandridis, V.; Spalevic, V.; Mincato, R.L. Wildfire Effects on Soil Erosion Dynamics: The Case of 2021 Megafires in Greece. Agric. For. 2022, 68, 49–63. [Google Scholar] [CrossRef]
- Bogorodskaya, A.V.; Kukavskaya, E.A. State of microbial communities in soils of deciduous and light coniferous forests of Central Siberia after logging and fires. Russ. J. For. Sci. 2016, 5, 383–396. (In Russian) [Google Scholar]
- Narita, K.; Harada, K.; Saito, K.; Sawada, Y.; Fukuda, M.; Tsuyuzaki, S. Vegetation and Permafrost Thaw Depth 10 Years after a Tundra Fire in 2002, Seward Peninsula, Alaska. Arctic Antarct. Alp. Res. 2015, 47, 547–559. [Google Scholar] [CrossRef]
- Vázquez, F.J.; Acea, M.J.; Carballas, T. Soil Microbial Populations after Wildfire. FEMS Microbiol. Ecol. 1993, 13, 93–103. [Google Scholar] [CrossRef]
- Bezkorovaynaya, I.N.; Tarasov, P.A.; Krasnoshchekova, E.N. Ecological state of soils after a fire in the pine forests of the middle taiga of the Krasnoyarsk territory. Bull. KrasGAU 2006, 13, 178–183. (In Russian) [Google Scholar]
- Bogdanov, V.V.; Prokushkin, A.S.; Prokushkin, S.G. The influence of grass-roots fires on the mobility of soil organic matter in the larch forests of the cryolithozone of Central Siberia. Bull. KrasGAU 2009, 2, 88–93. (In Russian) [Google Scholar]
- Dymov, A.A.; Dubrovsky, Y.A.; Gabov, D.N.; Zhangurov, E.V.; Nizovtsev, N.A. Influence of a fire in a northern taiga spruce forest on soil organic matter. Russ. J. For. Sci. 2015, 1, 52–62. (In Russian) [Google Scholar]
- Michaelides, R.; Schaefer, K.; Zebker, H.; Parsekian, A.; Liu, L.; Chen, J.; Natali, S.M.; Ludwig, S.; Schaefer, S. Inference of the impact of wildfire on permafrost and active layer thickness in a discontinuous permafrost region using the remotely sensed active layer thickness (ReSALT) algorithm. Environ. Res. Lett. 2018, 14, 035007. [Google Scholar] [CrossRef]
- Heim, R.J.; Bucharova, A.; Rieker, D.; Yurtaev, A.; Kamp, J.; Hölzel, N. Long-term effects of fire on Arctic tundra vegetation in Western Siberia. Sci. Total Environ. 2019, 760, 143425. [Google Scholar] [CrossRef]
- Ananyev, V.A. National Park “Vodlozerskii”. In Karelia: Encyclopedia; PetroPress Publishing House: Petrozavodsk, Russia, 2009; Volume 2, pp. 270–272. (In Russian) [Google Scholar]
- Nazarova, L.E. Climate. In Biodiversity of Karelia: Formation Conditions, Communities, Species; Gromtsev, A.N., Kitaev, S.P., Krutov, V.I., Eds.; Karelian Scientific Center of the Russian Academy of Sciences: Petrozavodsk, Russia, 2003; pp. 6–8. [Google Scholar]
- Filatov, N.N.; Derusova, O.V.; Zhukov, A.I. Atlas of the Republic of Karelia; Verso: Petrozavodsk, Russia, 2021; p. 48. (In Russian) [Google Scholar]
- Kipruhin, I.; Valdaev, V.; Volkova, A. Protected Natural Territories of the Republic of Karelia; Petrozavodsk, Russia, 2017; p. 432. (In Russian) [Google Scholar]
- Gromtsev, A.N. Primeval Forests in Karelia: Present State and Protection Prospects. Primeval Forests in the European Taiga Zone: The Present State and Conservation Problems; Review of the Proceedings of the International Scientific and Practical Conference; KarNC: Petrozavodsk, Russia, 1999; pp. 18–22. [Google Scholar]
- Pellikka, J.; Kurhinen, J.; Danilov, P.; Lindén, H.; Ovaskainen, O.; Gromtsev, A. Dimensions of the wildlife richness in Eastern Fennoscandia. Vestn. Ohotovedeniya 2014, 11, 266–269. [Google Scholar]
- Ananyev, V.A.; Raevsky, B.V.; Grabovik, S.I. Monitoring of biodiversity in the primeval spruce forests of Vodlozero national park. In Biodiversity and Conservation of Boreal Nature, Proceedings of the 10th Year Anniversary of the National Reserve Friendship, Vantaa, Finland, 2 October 2003; Heikkilä, R., Lindholm, T., Eds.; 485, Dark Oy; Kainuu Regional Environment Centre: Kajaani, Finland, 2003; pp. 113–116. [Google Scholar]
- Gromtsev, A.N. Fundamentals of Landscape Ecology of European Taiga Forests in Russia; KarRC of RAS: Petrozavodsk, Russia, 2008; p. 250. (In Russian) [Google Scholar]
- Zalesov, S.V. Forest Pyrology: Textbook; UGLTU: Ekaterinburg, Russia, 1998; p. 296. (In Russian) [Google Scholar]
- Zalesov, S.V. Forest Pyrology: Textbook; UGLTU: Ekaterinburg, Russia, 2021; p. 396. (In Russian) [Google Scholar]
- Andreeva, E.; Bakkal, I.; Gorshkov, V.; Lianguzova, I.; Maznaia, E.; Neshataev, V.; Neshataeva, V.; Stavrova, N.; Iarmishko, V.; Iarmishko, M. Methods for Studying Forest Communities; NIIKhimii: Saint Petersburg State University: Saint Petersburg, Russia, 2002; p. 240. (In Russian) [Google Scholar]
- Vorobyeva, L.A. Theory and Methods of Chemical Analysis of Soils; Publishing House of Moscow State University: Moscow, Russia, 1995; p. 136. [Google Scholar]
- Rules for Reforestation; Approved by order of the Ministry of Natural Resources of the Russian Federation, N 118, dated 25 March 2019; Ministry of Natural Resources of the Russian Federation: Moscow, Russia, 2020; p. 138.
- International Plant Names Index. The Royal Botanic Gardens, Kew, Harvard University Herbaria & Libraries and Australian National Botanic Gardens. 2021. Available online: http://www.ipni.org (accessed on 1 October 2021).
- Index Fungorum. CABI Bioscience. 2022. Available online: http://www.indexfungorum.org (accessed on 23 March 2022).
- Hodgetts, N.G.; Söderström, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schröck, C.; Sim-Sim, M.; et al. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- Lindblad, I. Wood-inhabiting fungi on fallen logs of Norway spruce: Relations to forest management and substrate quality. Nord. J. Bot. 1998, 18, 243–255. [Google Scholar] [CrossRef]
- Junninen, K.; Kouki, J. Are woodland key-habitats in Finland hotspots for polypores (Basidiomycota)? Scand. J. For. Res. 2006, 21, 32–40. [Google Scholar] [CrossRef]
- Shorokhova, E.V.; Shorokhov, A.A. Coarse woody debris dynamics and stores in a boreal virgin spruce forest. Ecol. Bull. 2001, 49, 129–137. [Google Scholar]
- Tuzov, V.K. (Ed.) Methods of Monitoring of Forest Pests and Diseases. Reference Book; VNIILM: Moscow, Russia, 2004; Volume III, p. 200. (In Russian) [Google Scholar]
- Vorontsov, A.I.; Mozolevskaya, E.G.; Sokolova, E.S. Technology of Forest Protection; Ecologia: Moscow, Russia, 1991; p. 304. [Google Scholar]
- Hill, M.O. DECORANA—A FORTRAN Program for Detrended Correspondence Analysis and Reciprocal Averaging. Ecology and Systematics; Cornell University: Ithaca, NY, USA, 1979; p. 52. [Google Scholar]
- Hill, M.O.; Gauch, H.G. Detrended correspondence analysis: An improved ordination technique. Vegetatio 1980, 42, 47–58. [Google Scholar] [CrossRef]
- Vasilevich, V.I. Statistical Methods in Geobotany; Nauka: Leningrad, Russia, 1969; p. 232. (In Russian) [Google Scholar]
- Anufriev, V.; Bespalaia, I.; Bolotov, I.; Ezhov, O.; Mamontov, V.; Mizin, I.; Novoselov, A.; Potapov, G.; Puchnina, L.; Pystina, T.; et al. (Eds.) Red Data Book of the Arkhangelsk Region; Northern (Arctic) Federal University: Arkhangelsk, Russia, 2020; p. 490. (In Russian) [Google Scholar]
- Kubartova, A.; Ottosson, E.; Dahlberg, A.; Stenlid, J. Patterns of fungal communities among and within decaying logs, revealed by 454 sequencing. Mol. Ecol. 2012, 21, 4514–4532. [Google Scholar] [CrossRef]
- Juutilainen, K.; Mönkkönen, M.; Kotiranta, H.; Halme, P. Resource use of wood-inhabiting fungi in different boreal forest types. Fungal Ecol. 2017, 27, 96–106. [Google Scholar] [CrossRef]
- Ruokolainen, A.; Shorokhova, E.; Penttilä, R.; Kotkova, V.; Kushnevskaya, E. A continuum of dead wood with various habitat elements maintains the diversity of wood-inhabiting fungi in an old-growth boreal forest. Eur. J. For. Res. 2018, 137, 707–718. [Google Scholar] [CrossRef]
- Yang, S.; Limpens, J.; Sterck Frank, J.; Sass-Klaassen, U.; Cornelissen, J.H.C.; Hefting, M.; Richard van Logtestijn, S.P.; Goudzwaard, L.; Dam, N.; Dam, M.; et al. Dead wood diversity promotes fungal diversity. Oikos 2021, 130, 2202–2216. [Google Scholar] [CrossRef]
- Kuznetsov, O.L. (Ed.) The Red Data Book of the Republic of Karelia; KONSTANTA: Belgorod, Russia, 2020; p. 448. (In Russian) [Google Scholar]
- Andersson, L.; Mikhailova, N.; Kuznetsova, E. (Eds.) Survey of Biologically Valuable Forests in North-Western European Russia; Identification Manual of Species to be Used During Survey at Stand Level; Pobeda: Saint-Petersburg, Russia, 2009; Volume 2, p. 258. (In Russian) [Google Scholar]
- Dymov, A.A.; Abakumov, E.V.; Bezkorovaynaya, I.N.; Prokushkin, A.S.; Kuzyakov, Y.V.; Milanovskyy, E.Y. Impact of forest fire on soil properties (review). Theor. Ecol. 2018, 4, 13–23. [Google Scholar] [CrossRef]
- Fernandes, P.M.; Vega, J.A.; Jiménez, E.; Rigolot, E. Fire resistance of European pines. For. Ecol. Manag. 2008, 256, 246–255. [Google Scholar] [CrossRef]
- Majder-Łopatka, M.; Szulc, W.; Rutkowska, B.; Ptasiński, D.; Kazberuk, W. Influence of fire on selected physico-chemical properties of forest soil. Soil Sci. Annu. 2019, 70, 39–43. [Google Scholar] [CrossRef]
- Tzvetkova, N.; Hadjiivanova, C. Chemical composition and biochemical changes in needles of Scots pine (Pinus sylvestris L.) stands at different stages of decline in Bulgaria. Trees Struct. Funct. 2006, 20, 405–409. [Google Scholar] [CrossRef]
- Barid, M.; Everett, R.; Zabowski, D. Wildfire effects on carbon and nitrogen in inland coniferous forests. Plant Soil 1999, 209, 233–243. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Kovaleva, N.M.; Ivanova, G.A.; Kukavskaya, E.A. Restoration of ground cover after ground fires in middle taiga pine forests. Russ. J. For. Sci. 2011, 5, 30–35. (In Russian) [Google Scholar]
- Zhila, S.V.; Ivanova, G.A.; Ivanov, V.A.; Tsvetkov, P.A. Reforestation after fires of different intensity in pine forests of Central Siberia. Sib. Lesn. Zurnal 2019, 6, 53–62. (In Russian) [Google Scholar]
- Degteva, S.V.; Dubrovsky, Y.A. Vegetation cover dynamics during regenerative successions on burnt areas of dark coniferous forests of the Pechoro-Ilychsky Reserve. Proc. Pechoro-Ilychsky Reserve 2010, 16, 35–41. (In Russian) [Google Scholar]
- Kalinin, K.K. Natural reforestation and formation of young stands in spruce and birch plantations on large burnt areas of the Middle Trans-Volga region. Bull. Mari State Tech. Univ. Ser. Forest. Ecology. Nat. Manag. 2010, 1, 5–15. (In Russian) [Google Scholar]
- Gavrilova, O.I.; Pak, K.A. Natural forest restoration after fires in the Republic of Karelia. Adv. Curr. Nat. Sci. 2017, 12, 38–44. (In Russian) [Google Scholar]
- Man, R.; Lieffers, V.J. Are mixtures of aspen and white spruce more productive than single species stands? For. Chron. 1999, 75, 505–513. [Google Scholar] [CrossRef]
- Gryazkin, A.V. Renewal Potential of Taiga Forests (On the Example of Spruce Forests in the North-West of Russia); SPbGLTA: Saint-Petersburg, Russia, 2001; p. 188. (In Russian) [Google Scholar]
- Ranade, S.S.; García-Gil, M.R. Molecular signatures of local adaptation to light in Norway Spruce. Planta 2021, 253, 53. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, Y. Species and stand dynamics in the mixed woods of Quebec’s southern boreal forest. Ecology 2000, 81, 1500–1516. [Google Scholar] [CrossRef]
- Farber, S.K. The impact of fires on the forests of Eastern Siberia. For. Inventory For. Plan. 2012, 1, 131–141. (In Russian) [Google Scholar]
- Parro, K.; Metslaid, M.; Renel, G.; Sims, A.; Stanturf, J.A.; Jõgiste, K.; Köster, K. Impact of postfire management on forest regeneration in a managed hemiboreal forest, Estonia. Can. J. For. Res. 2015, 45, 1192–1197. [Google Scholar] [CrossRef]
- Green, D.G. Simulated effects of fire, dispersal and spatial pattern on competition within forest mosaics. Vegetatio 1989, 82, 139–153. [Google Scholar] [CrossRef]
- Rajala, T.; Tuomivirta, T.; Pennanen, T.; Mäkipää, R. Habitat models of wood-inhabiting fungi along a decay gradient of Norway spruce logs. Fungal Ecol. 2015, 18, 48–55. [Google Scholar] [CrossRef]
- Müller, J.; Ulyshen, M.; Seibold, S.; Cadotte, M.; Chao, A.; Bässler, C.; Vogel, S.; Hagge, J.; Weiß, I.; Baldrian, P.; et al. Primary determinants of communities in deadwood vary among taxa but are regionally consistent. Oikos 2020, 129, 1579–1588. [Google Scholar] [CrossRef]
- Oberle, B.; Lee, M.R.; Myers, J.A.; Osazuwa-Peters, O.L.; Spasojevic, M.J.; Walton, M.L.; Young, D.F.; Zanne, A.E. Accurate forest projections require long-term wood decay experiments because plant trait effects change through time. Glob. Chang. Biol. 2020, 26, 864–875. [Google Scholar] [CrossRef]
- Jonsell, M.; Weslien, J.; Ehnström, B. Substrate requirements of red-listed saproxylic invertebrates in Sweden. Biodivers. Conserv. 1998, 7, 749–764. [Google Scholar] [CrossRef]
- Økland, B.; Bakke, A.; Hågvar, S.; Kvamme, T. What factors influence the diversity of saproxylic beetles? A multiscaled study from a spruce forest in southern Norway. Biodivers. Conserv. 1996, 5, 75–100. [Google Scholar] [CrossRef]
- Renvall, P. Community structure and dynamics of wood-rotting Basidiomycetes on decomposing conifer trunks in northern Finland. Karstenia 1995, 35, 1–51. [Google Scholar] [CrossRef]
- Vanha-Majamaa, I.; Lilja, S.; Ryömä, R.; Kotiaho, J.S.; Laaka-Lindberg, S.; Lindberg, H.; Puttonen, P.; Tamminen, P.; Toivanen, T.; Kuuluvainen, T. Rehabilitating boreal forest structure and species composition in Finland through logging, dead wood creation and fire: The EVO experiment. For. Ecol. Manag. 2007, 250, 77–88. [Google Scholar] [CrossRef]
- Dynesius, M.; Gibb, H.; Hjältén, J. Surface Covering of Downed Logs: Drivers of a Neglected Process in Dead Wood Ecology. PLoS ONE 2010, 5, e13237. [Google Scholar] [CrossRef] [PubMed]








 —SP 6 in 2014,
—SP 6 in 2014,  —SP 6 in 2020,
—SP 6 in 2020,  —SP 8 in 2014,
—SP 8 in 2014,  —SP 8 in 2020,
—SP 8 in 2020,  —SP 9 in 2014,
—SP 9 in 2014,  —SP 9 in 2020. A–C—see text for explanations. Axis loadings: axis 1—0.33, axis 2—0.09.
—SP 6 in 2014, —SP 6 in 2020, —SP 8 in 2014, —SP 8 in 2020, —SP 9 in 2014, —SP 9 in 2020. A–C—see text for explanations. Axis loadings: axis 1—0.33, axis 2—0.09.
—SP 9 in 2020. A–C—see text for explanations. Axis loadings: axis 1—0.33, axis 2—0.09.
—SP 6 in 2014, —SP 6 in 2020, —SP 8 in 2014, —SP 8 in 2020, —SP 9 in 2014, —SP 9 in 2020. A–C—see text for explanations. Axis loadings: axis 1—0.33, axis 2—0.09.
| Tree and Shrub Species | Number of Plants, 100 pcs./ha | |||||
|---|---|---|---|---|---|---|
| 2014 | 2020 | |||||
| SP 6 | SP 8 | SP 9 | SP 6 | SP 8 | SP 9 | |
| Coniferous species | ||||||
| Picea abies (L.) H. Karst. | 21.6 | 8.6 | 14.0 | 20.3 | 11.8 | 18.5 |
| Pinus sylvestris L. | 19.8 | 10.3 | 7.2 | 14.4 | 12.5 | 12.0 |
| Coniferous species total: | 41.4 | 18.9 | 21.2 | 34.7 | 24.3 | 30.5 |
| Deciduous species | ||||||
| Alnus incana (L.) Moench | – | – | – | 0.1 | – | – |
| Betula pendula Roth | – | – | – | – | – | 0.1 |
| Betula pubescens Ehrh. | 222.4 | 30.9 | 318.5 | 110.1 | 71.4 | 290.0 |
| Populus tremula L. | 11.2 | 15.1 | 40.7 | 9.9 | 12.6 | 24.5 |
| Salix aurita L. | – | – | – | – | 0.1 | – |
| Salix caprea L. | 12.4 | 16.9 | 40.6 | 9.1 | 16.5 | 27.2 |
| Salix myrsinifolia Salisb. | – | – | – | – | – | 0.1 |
| Salix pentandra L. | – | – | – | – | 0.2 | – |
| Salix phylicifolia L. | – | – | – | – | 0.1 | – |
| Sorbus aucuparia L. | – | – | – | – | 1.8 | – |
| Deciduous species total: | 246.0 | 62.9 | 399.8 | 129.2 | 102.7 | 341.9 |
| Total amount of regeneration, 1000 pcs./ha | 287.4 | 81.8 | 421.0 | 163.9 | 127.0 | 372.4 |
| Species | Species Occurrence/Average Percentage Cover | ||||||
|---|---|---|---|---|---|---|---|
| 2014 | 2020 | ||||||
| SP 6 | SP 8 | SP 9 | SP 6 | SP 8 | SP 9 | SP10-C * | |
| Herb-subshrub layer | |||||||
| Avenella flexuosa (L.) Drejer | 23/2 | 60/4 | 10/<1 | 36/3 | 83/3 | 48/2 | – |
| Chamaenerion angustifolium (L.) Scop. | 73/10 | 88/10 | 100/20 | 55/1 | 82/2 | 87/2 | – |
| Dactylorhiza maculata (L.) Soó | – | – | – | 1/<1 | – | – | – |
| Goodyera repens (L.) R. Br. | – | – | – | – | – | – | <1 |
| Linnaea borealis L. | – | – | – | – | 2/<1 | – | <1 |
| Luzula pilosa (L.) Willd. | – | 20/1 | 2/<1 | – | 73/2 | 9/<1 | – |
| Lycopodium clavatum L. | – | – | – | – | – | 1/<1 | – |
| Maianthemum bifolium (L.) F.W. Schmidt | – | – | – | – | 3/<1 | – | <1 |
| Melampyrum pratense L. | – | – | – | – | 47/1 | – | <1 |
| Vaccinium myrtillus L. | 42/9 | 66/10 | 4/<1 | 42/3 | 56/5 | 8/1 | 45 |
| Vaccinium uliginosum L. | – | 6/<1 | – | – | 12/<1 | – | – |
| Vaccinium vitis-idaea L. | 25/1 | 52/3 | 3/<1 | 59/7 | 80/10 | 20/<1 | 35 |
| Moss-lichen layer | |||||||
| Ceratodon purpureus (Hedw.) Brid. | – | – | – | 1/<1 | 1/<1 | – | – |
| Dicranum majus Sm. | – | – | – | 1/<1 | 1/<1 | – | – |
| Dicranum polysetum Sw. ex anon. | – | – | – | – | 1/<1 | – | – |
| Dicranum scoparium Hedw. | 18/1 | – | – | 29/7 | 17/<1 | – | <1 |
| Hylocomium splendens (Hedw.) Bruch et al. | 1/<1 | 1/<1 | – | 1/<1 | 4/<1 | – | 8 |
| Marchantia polymorpha L. | 38/3 | 35/3 | 92/20 | – | – | – | – |
| Pleurozium schreberi (Brid.) Mitt. | 6/<1 | 3/<1 | – | 42/4 | 20/3 | 15/1 | 75 |
| Pohlia nutans (Hedw.) Lindb. | – | – | – | 15/<1 | 11/1 | 14/1 | – |
| Polytrichum commune Hedw. | – | – | – | – | 1/<1 | – | – |
| Polytrichum juniperinum Hedw. | 41/2 | 75/10 | 55/2 | 60/10 | 90/25 | 98/15 | – |
| Ptilium crista-castrensis (Hedw.) De Not. | – | – | – | – | 1/<1 | – | – |
| Species in total | 9 | 10 | 7 | 10 | 15 | 9 | 9 |
| Total cover of the herb-subshrub layer, % | 20 | 25 | 20 | 10 | 20 | 10 | 75 |
| Total cover of the moss-lichen layer, % | 7 | 10 | 20 | 20 | 30 | 10 | 80 |
| Total ground vegetation cover, % | 25 | 40 | 40 | 30 | 50 | 20 | 95 |
| Tree Species | Size Distribution of Regenerating Trees, 1000 Plants/ha | |||||||
|---|---|---|---|---|---|---|---|---|
| SP 6 | SP 8 | SP 9 | ||||||
| Small | Medium | Small | Medium | Large | Small | Medium | Large | |
| Betula pubescens | 55.0 | 55.1 | 19.4 | 51.9 | 0.1 | 10.4 | 279.6 | 0 |
| Populus tremula | 4.3 | 5.6 | 5.7 | 6.9 | 0 | 1.6 | 22.9 | 0 |
| Salix caprea | 2.1 | 7 | 8.2 | 8.3 | 0 | 1.7 | 25.5 | 0 |
| Deciduous in total, 1000 plants/ha | 61.4 | 67.7 | 33.3 | 67.1 | 0.1 | 13.7 | 328 | 0 |
| Picea abies | 20.2 | 0.1 | 10.8 | 1.0 | 0 | 18.4 | 0.1 | 0 |
| Pinus sylvestris | 9.8 | 4.6 | 4.0 | 8.5 | 0 | 1.0 | 10.9 | 0.1 |
| Coniferous in total, 1000 plants/ha | 30 | 4.7 | 14.8 | 9.5 | 0 | 19.4 | 11 | 0.1 |
| Total, 1000 plants/ha | 91.4 | 72.4 | 48.1 | 76.6 | 0.1 | 33.1 | 339.0 | 0.1 |
| Parameters | Sample Plot Number | Soil Horizons | ||
|---|---|---|---|---|
| Organic | Mineral | |||
| O | E | BF | ||
| N, % | 6 | 1.20 | 0.08 | 0.03 |
| 8 | 0.90 | 0.02 | 0.01 | |
| 9 | 0.71 | 0.01 | 0.04 | |
| 10-C | 1.24 | 0.01 | 0.09 | |
| P, mg/100 g | 6 | 23.10 | 0.13 | 4.17 |
| 8 | 11.06 | 0.82 | 2.30 | |
| 9 | 24.96 | 0.60 | 2.30 | |
| 10-C | 63.09 | 0.25 | 6.40 | |
| K, mg/kg | 6 | 579.52 | 4.73 | 9.87 |
| 8 | 431.18 | 9.79 | 11.89 | |
| 9 | 587.75 | 4.97 | 17.55 | |
| 10-C | 2190.09 | 9.67 | 13.28 | |
| C, % | 6 | 44.64 | 0.46 | 0.88 |
| 8 | 46.93 | 1.02 | 2.29 | |
| 9 | 31.49 | 0.22 | 1.44 | |
| 10-C | 46.29 | 0.28 | 2.32 | |
| Ash content/loss on ignition | 6 | 18.3/- | -/0.5 | -/1.6 |
| 8 | 14.78/- | -/2.0 | -/4.8 | |
| 9 | 25.68/- | -/0.52 | -/3.06 | |
| 10-C | 12.21/- | -/0.3 | -/3.1 | |
| V, % | 6 | 19.85 | 24.62 | 14.04 |
| 8 | 3.16 | 0.00 | 12.17 | |
| 9 | 8.83 | 0.00 | 0.00 | |
| 10-C | 12.70 | 10.26 | 11.73 | |
| pHH2O | 6 | 3.4 | 3.38 | 4.13 |
| 8 | 2.75 | 3.24 | 4.03 | |
| 9 | 3.01 | 3.29 | 4.5 | |
| 10-C | 3.25 | 3.19 | 3.87 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ananyev, V.A.; Timofeeva, V.V.; Kryshen’, A.M.; Pekkoev, A.N.; Kostina, E.E.; Ruokolainen, A.V.; Moshnikov, S.A.; Medvedeva, M.V.; Polevoi, A.V.; Humala, A.E. Fire Severity Controls Successional Pathways in a Fire-Affected Spruce Forest in Eastern Fennoscandia. Forests 2022, 13, 1775. https://doi.org/10.3390/f13111775
Ananyev VA, Timofeeva VV, Kryshen’ AM, Pekkoev AN, Kostina EE, Ruokolainen AV, Moshnikov SA, Medvedeva MV, Polevoi AV, Humala AE. Fire Severity Controls Successional Pathways in a Fire-Affected Spruce Forest in Eastern Fennoscandia. Forests. 2022; 13(11):1775. https://doi.org/10.3390/f13111775
Chicago/Turabian StyleAnanyev, Vladimir A., Vera V. Timofeeva, Alexandr M. Kryshen’, Alexey N. Pekkoev, Ekaterina E. Kostina, Anna V. Ruokolainen, Sergei A. Moshnikov, Maria V. Medvedeva, Alexei V. Polevoi, and Andrey E. Humala. 2022. "Fire Severity Controls Successional Pathways in a Fire-Affected Spruce Forest in Eastern Fennoscandia" Forests 13, no. 11: 1775. https://doi.org/10.3390/f13111775
APA StyleAnanyev, V. A., Timofeeva, V. V., Kryshen’, A. M., Pekkoev, A. N., Kostina, E. E., Ruokolainen, A. V., Moshnikov, S. A., Medvedeva, M. V., Polevoi, A. V., & Humala, A. E. (2022). Fire Severity Controls Successional Pathways in a Fire-Affected Spruce Forest in Eastern Fennoscandia. Forests, 13(11), 1775. https://doi.org/10.3390/f13111775