
Assessing Forest Structural and Topographic Effects on Habitat Productivity for the Endangered Apennine Brown Bear


Abstract
:1. Introduction
2. Materials and Methods
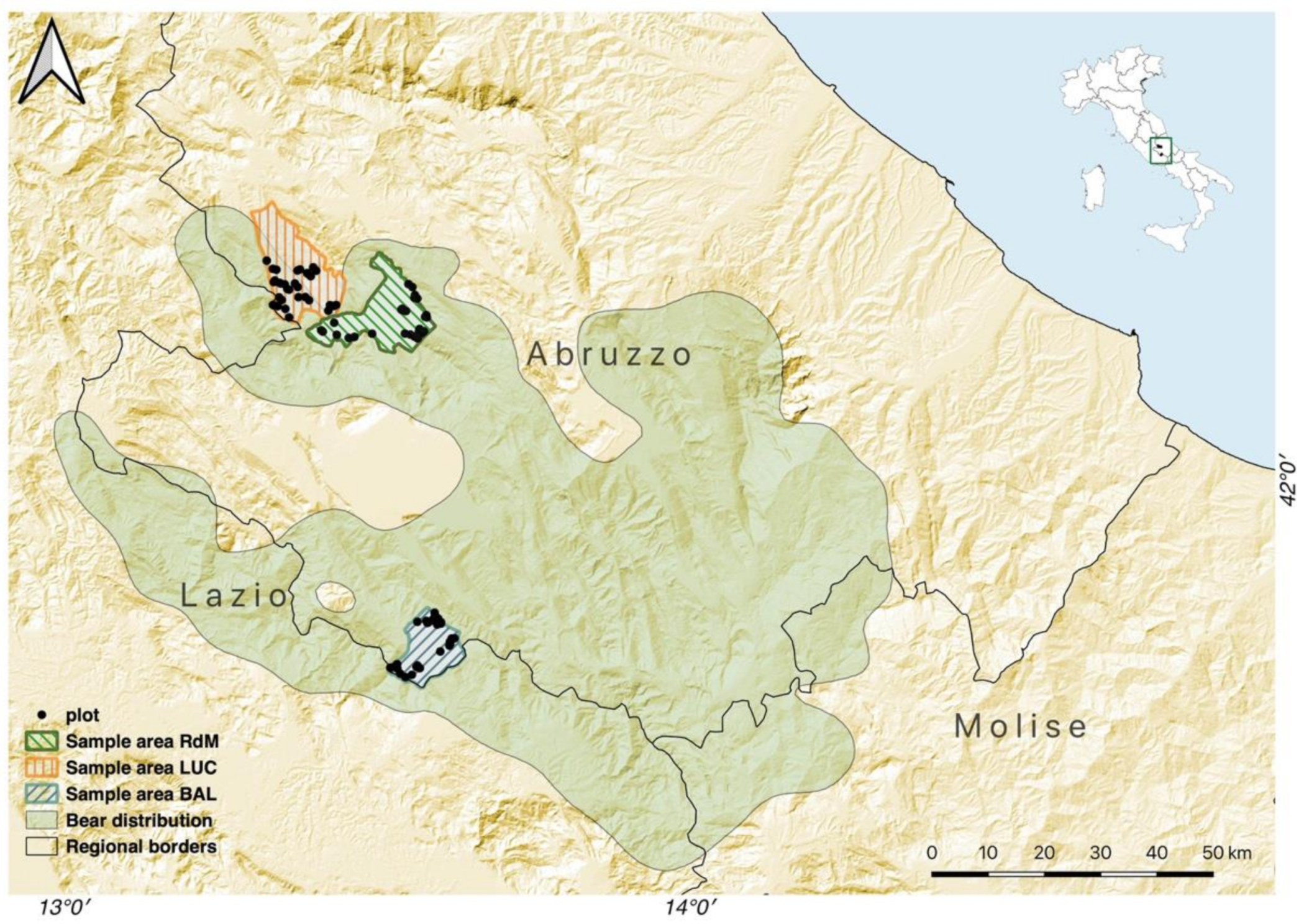
2.1. Study Area
2.2. Study Design
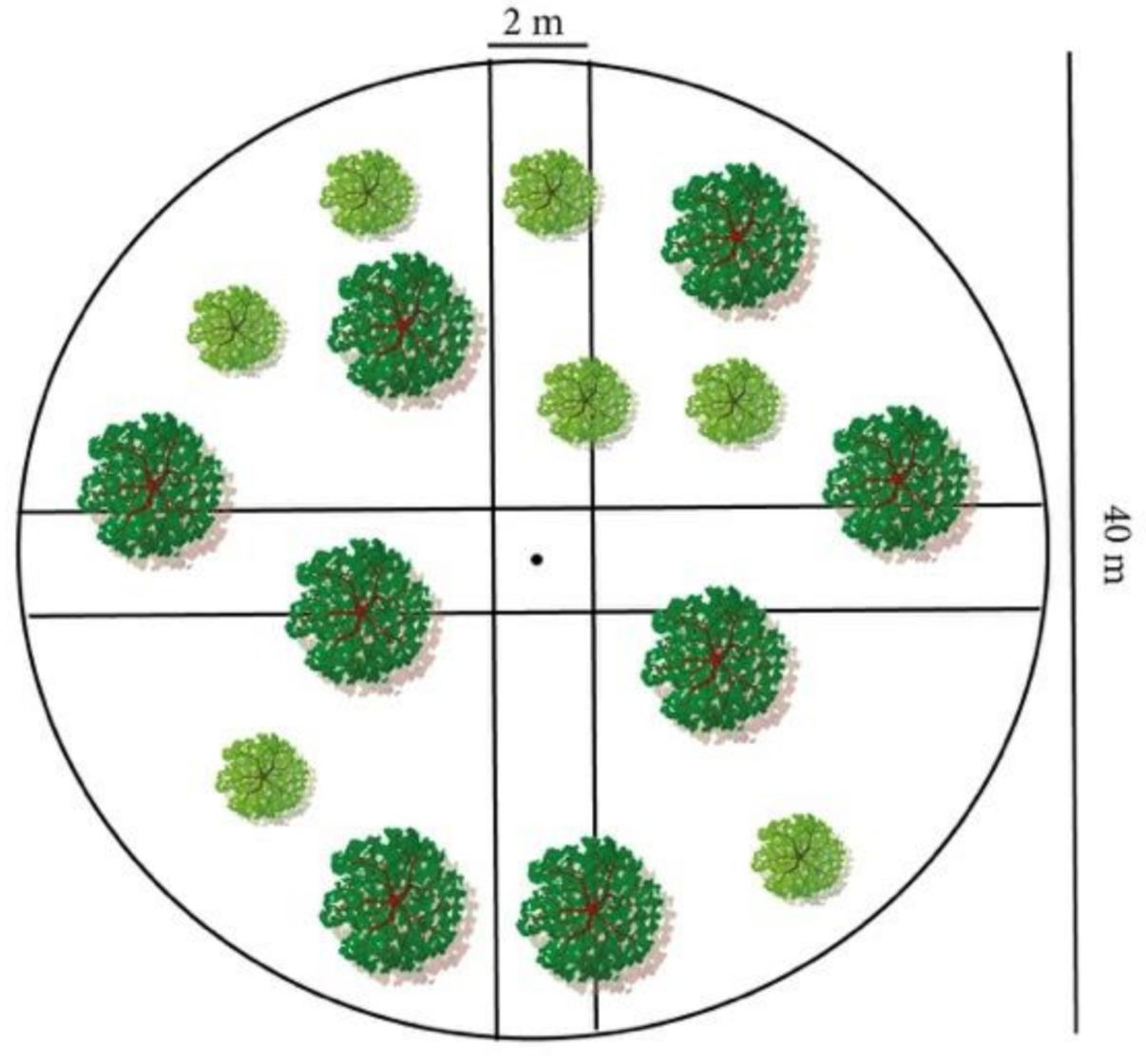
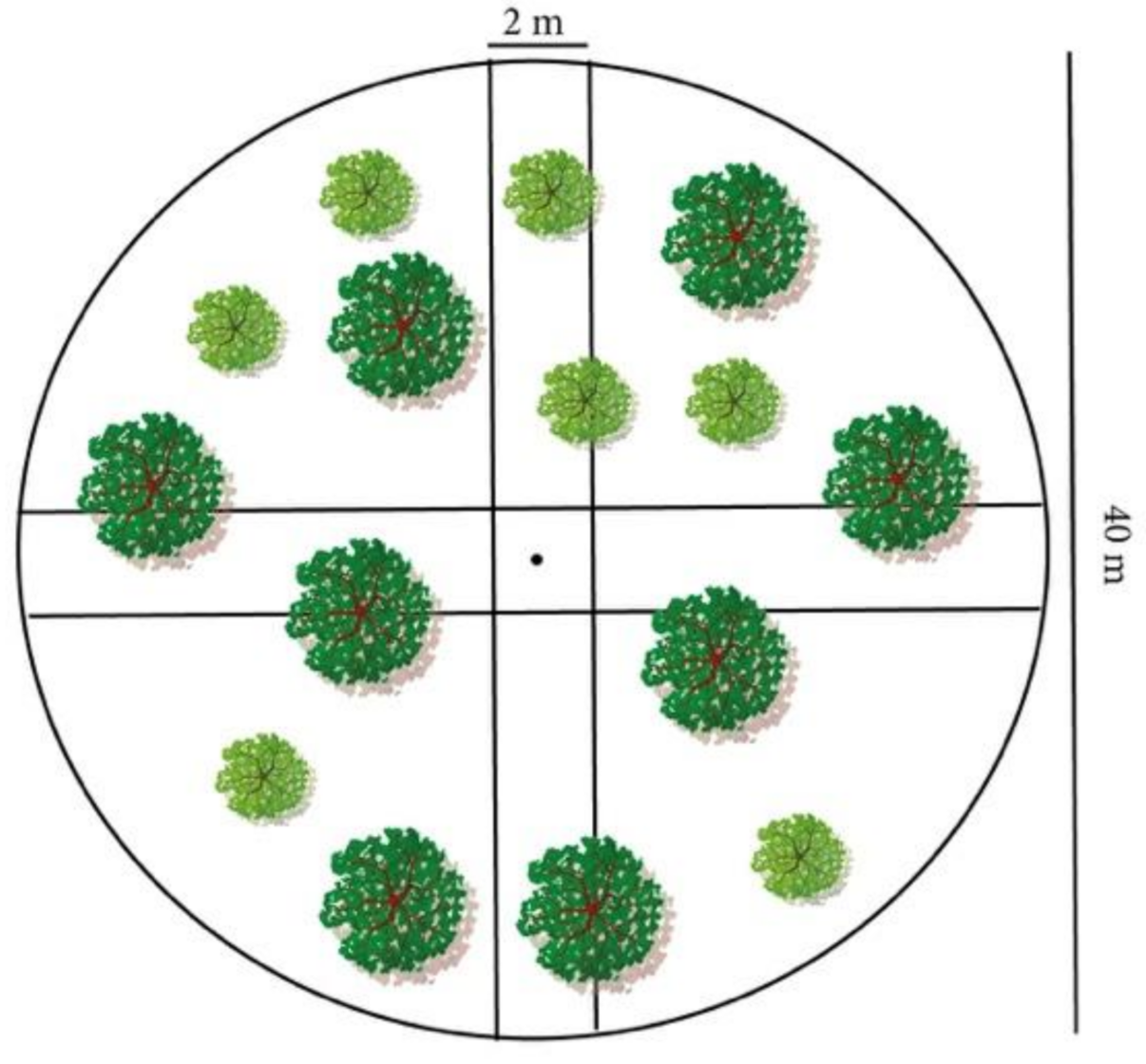
2.3. Data Collection
2.4. Model Development
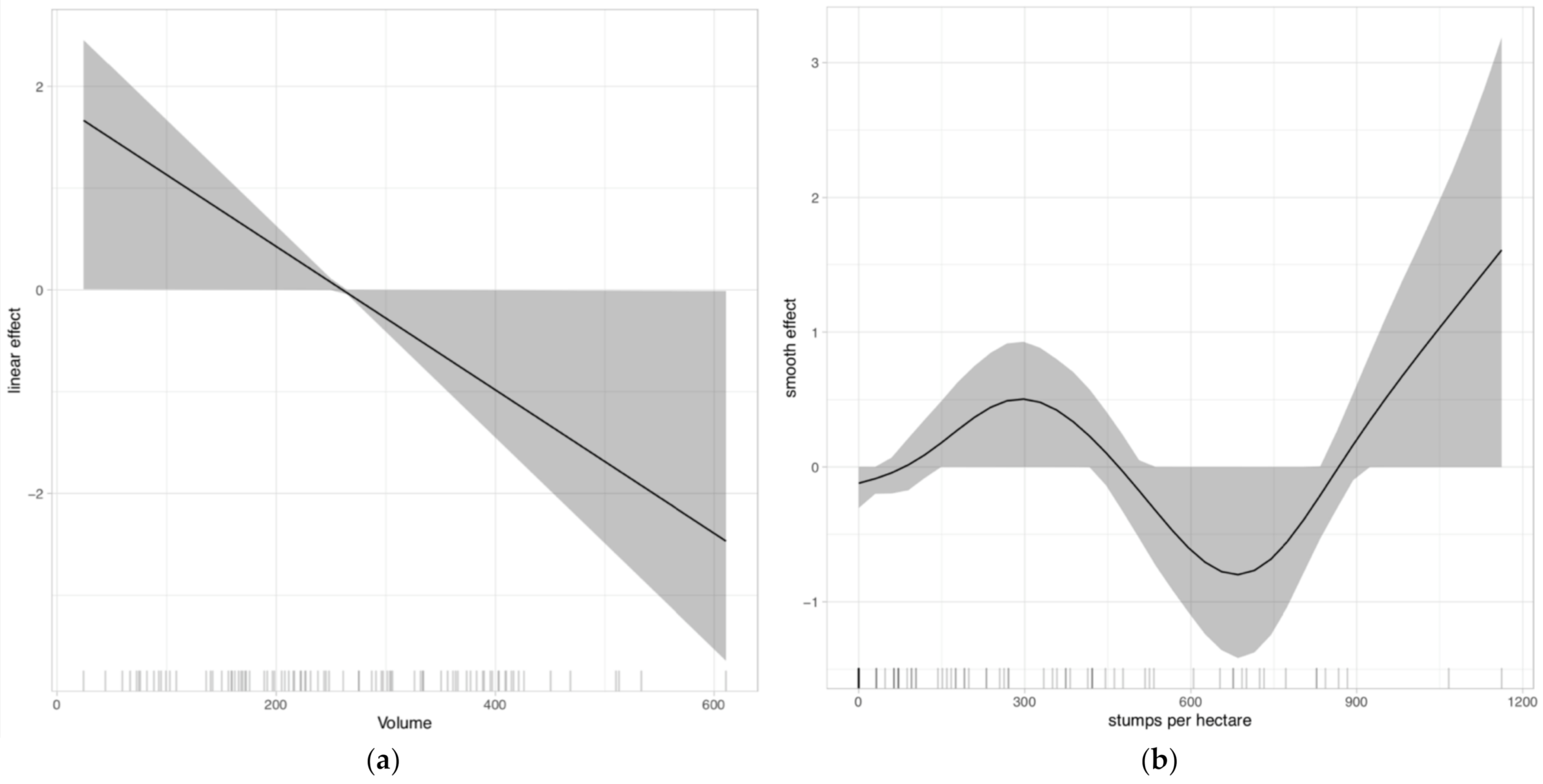
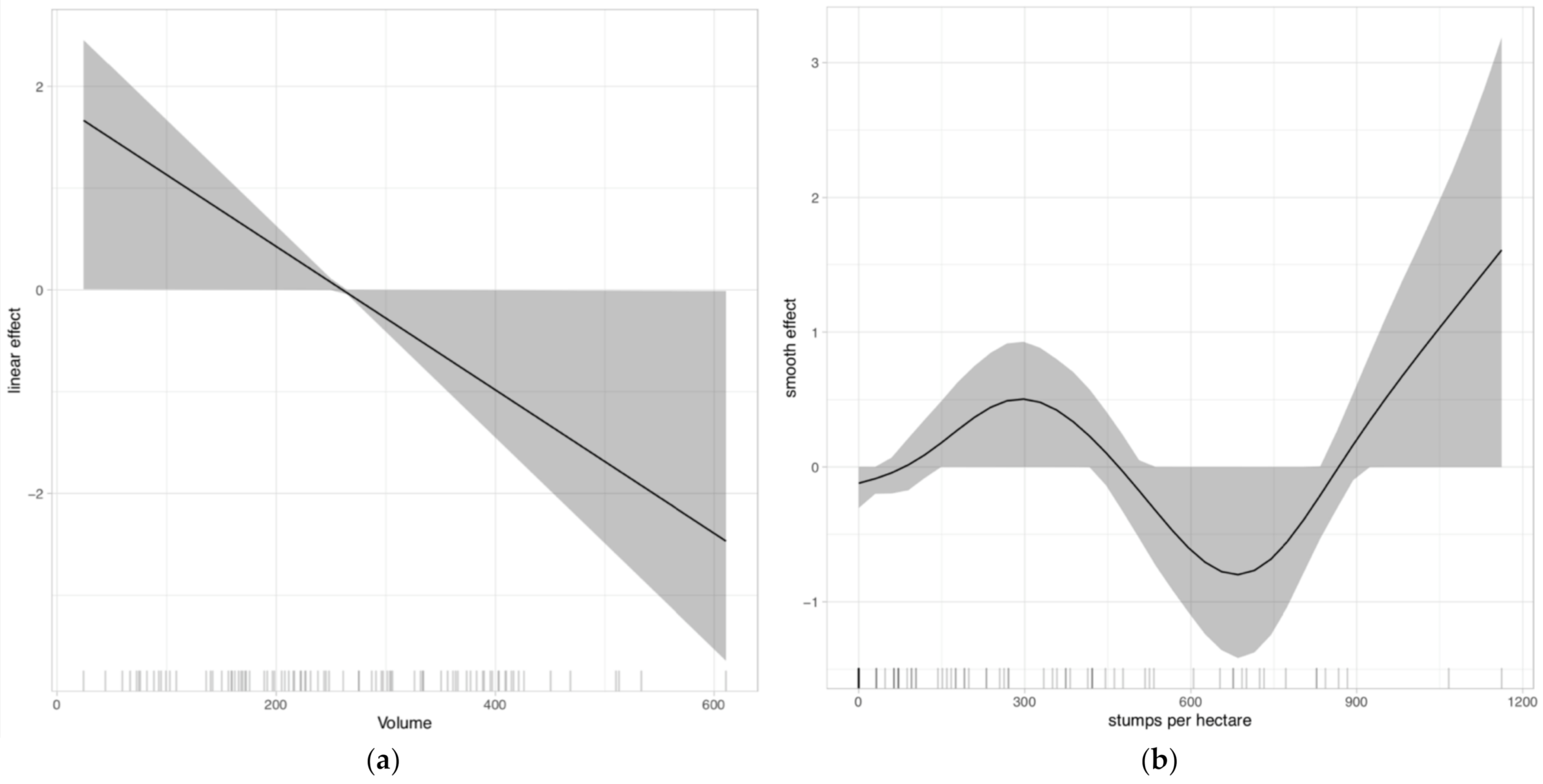
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; MacE, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Hall, L.S.; Krausman, P.R.; Morrison, M.L. The habitat concept and a plea for standard terminology. Wildl. Soc. Bull. 1997, 25, 173–182. [Google Scholar]
- Grodsky, S.; Moorman, C.; Russell, K. Forest Wildlife Management. In Ecological Forest Management Handbook; Larocque, G.R., Ed.; CRC Press Taylor & Francis Group: New York, NY, USA, 2015; pp. 47–85. [Google Scholar]
- Chaudhary, A.; Burivalova, Z.; Koh, L.P.; Hellweg, S. Impact of Forest Management on Species Richness: Global Meta-Analysis and Economic Trade-Offs. Sci. Rep. 2016, 6, 23954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holbrook, J.D.; Squires, J.R.; Bollenbacher, B.; Graham, R.; Olson, L.E.; Hanvey, G.; Jackson, S.; Lawrence, R.L. Spatio-temporal responses of Canada lynx (Lynx canadensis) to silvicultural treatments in the Northern Rockies, US. For. Ecol. Manag. 2018, 422, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Askins, R.A. Sustaining biological diversity in early successional communities: The challenge of managing unpopular habitats. Wildl. Soc. Bull. 2001, 29, 407–412. [Google Scholar]
- Paillet, Y.; Bergès, L.; Hjältén, J.; Odor, P.; Avon, C.; Bernhardt-Römermann, M.; Bijlsma, R.-J.; De Bruyn, L.; Fuhr, M.; Grandin, U.; et al. Biodiversity differences between managed and unmanaged forests: Meta-analysis of species richness in Europe. Conserv. Biol. 2010, 24, 101–112. [Google Scholar] [CrossRef]
- Riffell, S.; Verschuyl, J.; Miller, D.; Wigley, T.B. Biofuel harvests, coarse woody debris, and biodiversity—A meta-analysis. For. Ecol. Manag. 2011, 261, 878–887. [Google Scholar] [CrossRef]
- Fedrowitz, K.; Koricheva, J.; Baker, S.C.; Lindenmayer, D.B.; Palik, B.; Rosenvald, R.; Beese, W.; Franklin, J.F.; Kouki, J.; Macdonald, E.; et al. Can retention forestry help conserve biodiversity? A meta-analysis. J. Appl. Ecol. 2014, 51, 1669–1679. [Google Scholar] [CrossRef] [Green Version]
- Verschuyl, J.; Riffell, S.; Miller, D.; Wigley, T.B. Biodiversity response to intensive biomass production from forest thinning in North American forests—A meta-analysis. For. Ecol. Manag. 2011, 261, 221–232. [Google Scholar] [CrossRef]
- Franklin, C.M.A.; Macdonald, S.E.; Nielsen, S.E. Can retention harvests help conserve wildlife? Evidence for vertebrates in the boreal forest. Ecosphere 2019, 10, e02632. [Google Scholar] [CrossRef]
- Fisher, J.; Wilkinson, L. The response of mammals to forest fire and timber harvest in the North American boreal forest. Mamm. Rev. 2005, 35, 51–81. [Google Scholar] [CrossRef]
- Kearney, S.P.; Coops, N.C.; Stenhouse, G.B.; Nielsen, S.E.; Hermosilla, T.; White, J.C.; Wulder, M.A. Grizzly bear selection of recently harvested forests is dependent on forest recovery rate and landscape composition. For. Ecol. Manag. 2019, 449, 117459. [Google Scholar] [CrossRef]
- Schwenk, W.S.; Donovan, T.; Keeton, W.S.; Nunery, J.S. Carbon storage, timber production, and biodiversity: Comparing ecosystem services with multi-criteria decision analysis. Ecol. Appl. 2012, 22, 1612–1627. [Google Scholar] [CrossRef]
- Holbrook, J.D.; Squires, J.R.; Bollenbacher, B.; Graham, R.; Olson, L.E.; Hanvey, G.; Jackson, S.; Lawrence, R.L.; Savage, S.L. Management of forests and forest carnivores: Relating landscape mosaics to habitat quality of Canada lynx at their range periphery. For. Ecol. Manag. 2019, 437, 411–425. [Google Scholar] [CrossRef]
- Ciucci, P.; Gervasi, V.; Boitani, L.; Boulanger, J.; Paetkau, D.; Prive, R.; Tosoni, E. Estimating abundance of the remnant Apennine brown bear population using multiple noninvasive genetic data sources. J. Mammal. 2015, 96, 206–220. [Google Scholar] [CrossRef] [Green Version]
- Benazzo, A.; Trucchi, E.; Cahill, J.A.; Delser, P.M.; Mona, S.; Fumagalli, M.; Bunnefeld, L.; Cornetti, L.; Ghirotto, S.; Girardi, M.; et al. Survival and divergence in a small group: The extraordinary genomic history of the endangered Apennine brown bear stragglers. Proc. Natl. Acad. Sci. USA 2017, 114, E9589–E9597. [Google Scholar] [CrossRef] [Green Version]
- Gervasi, V.; Ciucci, P. Demographic projections of the Apennine brown bear population Ursus arctos marsicanus (Mammalia: Ursidae) under alternative management scenarios. Eur. Zool. J. 2018, 85, 242–252. [Google Scholar] [CrossRef] [Green Version]
- Ciucci, P.; Altea, T.; Antonucci, A.; Chiaverini, L.; Di Croce, A.; Fabrizio, M.; Forconi, P.; Latini, R.; Maiorano, L.; Monaco, A.; et al. Distribution of the brown bear (Ursus arctos marsicanus) in the central apennines, Italy, 2005–2014. Hystrix 2017, 28. [Google Scholar] [CrossRef]
- Maiorano, L.; Chiaverini, L.; Falco, M.; Ciucci, P. Combining multi-state species distribution models, mortality estimates, and landscape connectivity to model potential species distribution for endangered species in human dominated landscapes. Biol. Conserv. 2019, 237, 19–27. [Google Scholar] [CrossRef]
- Anonymous. Piano d ’Azione Nazionale per la Tutela Dell’orso Bruno Marsicano-PATOM; Ministero dell’ Ambiente-ISPRA: Roma, Italy, 2011; Volume 37. [Google Scholar]
- Falcucci, A.; Ciucci, P.; Maiorano, L.; Gentile, L.; Boitani, L. Assessing habitat quality for conservation using an integrated occurrence-mortality model. J. Appl. Ecol. 2009, 46, 600–609. [Google Scholar] [CrossRef]
- Posillico, M.; Meriggi, A.; Pagnin, E.; Lovari, S.; Russo, L. A habitat model for brown bear conservation and land use planning in the central Apennines. Biol. Conserv. 2004, 118, 141–150. [Google Scholar] [CrossRef]
- Tosoni, E.; Boitani, L.; Gentile, L.; Gervasi, V.; Latini, R.; Ciucci, P. Assessment of key reproductive traits in the Apennine brown bear population. Ursus 2017, 28, 105. [Google Scholar] [CrossRef]
- Gervasi, V.; Boitani, L.; Paetkau, D.; Posillico, M.; Randi, E.; Ciucci, P. Estimating survival in the Apennine brown bear accounting for uncertainty in age classification. Popul. Ecol. 2017, 59, 119–130. [Google Scholar] [CrossRef]
- Ciucci, P.; Tosoni, E.; Di Domenico, G.; Quattrociocchi, F.; Boitani, L. Seasonal and annual variation in the food habits of Apennine brown bears, central Italy. J. Mammal. 2014, 95, 572–586. [Google Scholar] [CrossRef] [Green Version]
- Larsen, T.A.; Nielsen, S.E.; Cranston, J.; Stenhouse, G.B. Do remnant retention patches and forest edges increase grizzly bear food supply? For. Ecol. Manag. 2019, 433, 741–761. [Google Scholar] [CrossRef]
- Nielsen, S.E.; McDermid, G.; Stenhouse, G.B.; Boyce, M.S. Dynamic wildlife habitat models: Seasonal foods and mortality risk predict occupancy-abundance and habitat selection in grizzly bears. Biol. Conserv. 2010, 143, 1623–1634. [Google Scholar] [CrossRef]
- McLellan, B.N. Some mechanisms underlying variation in vital rates of grizzly bears on a multiple use landscape. J. Wildl. Manag. 2015, 79, 749–765. [Google Scholar] [CrossRef]
- Bojarska, K.; Selva, N. Spatial patterns in brown bear Ursus arctos diet: The role of geographical and environmental factors. Mamm. Rev. 2012, 42, 120–143. [Google Scholar] [CrossRef]
- Nielsen, S.E.; Boyce, M.S.; Stenhouse, G.B. Grizzly bears and forestry I. Selection of clearcuts by grizzly bears in west-central Alberta, Canada Scott. For. Ecol. Manag. 2004, 199, 51–65. [Google Scholar] [CrossRef]
- Clevenger, A.P.; Purroy, F.J.; Pelton, M.R. Food Habits of Brown Bears (Ursus arctos) in the Cantabrian Mountains, Spain. J. Mammal. 1992, 73, 415–421. [Google Scholar] [CrossRef]
- Costello, C.M.C.; Jones, D.E.; Inman, R.R.M.; Inman, K.H.; Bruce, C.; Quigley, H.B. Relationship of variable mast production to American black bear reproductive parameters in New Mexico. Ursus 2003, 14, 1–16. [Google Scholar]
- Reynolds-Hogland, M.J.; Pacifici, L.B.; Mitchell, M.S. Linking resources with demography to understand resource limitation for bears. J. Appl. Ecol. 2007, 44, 1166–1175. [Google Scholar] [CrossRef]
- Hashimoto, Y.; Kaji, M.; Sawada, H.; Takatsuki, S. Five-year study on the autumn food habits of the Asiatic black bear in relation to nut production. Ecol. Res. 2003, 18, 485–492. [Google Scholar] [CrossRef]
- Atkinson, S.N.; Ramsay, M.A. The Effects of Prolonged Fasting of the Body Composition and Reproductive Success of Female Polar Bears (Ursus maritimus). Funct. Ecol. 1995, 9, 559. [Google Scholar] [CrossRef]
- Tosoni, E.; Boitani, L.; Mastrantonio, G.; Latini, R.; Ciucci, P. Counts of unique females with cubs in the Apennine brown bear population, 2006–2014. Ursus 2017, 28, 1–14. [Google Scholar] [CrossRef]
- Mitchell, M.S.; Powell, R.A. Response of Black Bears to Forest Management in the Southern Appalachian Mountains. J. Wildl. Manag. 2003, 67, 692. [Google Scholar] [CrossRef]
- Brodeur, V.; Ouellet, J.-P.; Courtois, R.; Fortin, D. Habitat selection by black bears in an intensively logged boreal forest. Can. J. Zool. 2008, 86, 1307–1316. [Google Scholar] [CrossRef]
- Davis, H.; Hamilton, A.N.; Harestad, A.S.; Weir, R.D. Longevity and reuse of black bear dens in managed forests of coastal British Columbia. J. Wildl. Manag. 2012, 76, 523–527. [Google Scholar] [CrossRef]
- Zager, P.; Jonkel, C.; Habeck, J. Logging and Wildfire Influence on Grizzly Bear Habitat in Northwestern Montana. Int. Conf. Bear Res. Manag. 1980, 5, 124–132. [Google Scholar] [CrossRef]
- Wielgus, R.B.; Vernier, P.R. Grizzly bear selection of managed and unmanaged forests in the Selkirk Mountains. Can. J. For. Res 2003, 33, 822–829. [Google Scholar] [CrossRef]
- Nielsen, S.E.; Munro, R.H.M.; Bainbridge, E.L.; Stenhouse, G.B.; Boyce, M.S. Grizzly bears and forestry II. Distribution of grizzly bear foods in clearcuts of west-central Alberta, Canada. For. Ecol. Manag. 2004, 199, 67–82. [Google Scholar] [CrossRef]
- Braid, A.C.R.; Manzer, D.; Nielsen, S.E. Wildlife habitat enhancements for grizzly bears: Survival rates of planted fruiting shrubs in forest harvests. For. Ecol. Manag. 2016, 369, 144–154. [Google Scholar] [CrossRef]
- Souliere, C.M.; Coogan, S.C.P.; Stenhouse, G.B.; Nielsen, S.E. Harvested forests as a surrogate to wildfires in relation to grizzly bear food- supply in west-central Alberta. For. Ecol. Manag. 2020, 456, 117685. [Google Scholar] [CrossRef]
- Ciucci, P.; Boitani, L. The Apennine brown bear: A critical review of its status and conservation problems. Ursus 2008, 19, 130–145. [Google Scholar] [CrossRef]
- Kane, V.R.; Lutz, J.A.; Alina Cansler, C.; Povak, N.A.; Churchill, D.J.; Smith, D.F.; Kane, J.T.; North, M.P. Water balance and topography predict fire and forest structure patterns. For. Ecol. Manag. 2015, 338, 1–13. [Google Scholar] [CrossRef]
- Tateno, R.; Takeda, H. Forest structure and tree species distribution in relation to topography-mediated heterogeneity of soil nitrogen and light at the forest floor. Ecol. Res. 2003, 18, 559–571. [Google Scholar] [CrossRef]
- Underwood, E.C.; Viers, J.H.; Quinn, J.F.; North, M. Using topography to meet wildlife and fuels treatment objectives in fire-suppressed landscapes. Environ. Manag. 2010, 46, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Morini, P.; Pinchera, F.P.; Nucci, L.M.; Ferlini, F.; Cecala, S.; Di Nino, O.; Penteriani, V. Brown bears in Central Italy: A 15-year study on bear occurrence. Eur. Zool. J. 2017, 84, 26–33. [Google Scholar] [CrossRef]
- Ministero dell’Ambiente e della Tutela del Territorio e del Mare Carta Fitoclimatica d’Italia. Available online: http://www.pcn.minambiente.it/viewer/index.php?services=Fitoclima (accessed on 7 April 2020).
- Collalti, D.; D’Alessandro, L.; Marchetti, M.; Sebastiani, A. La Carta Tipologico-Forestale Della Regione Abruzzo. Volume Generale; Regione Abruzzo: L’Aquila, Italy, 2009; p. 340. [Google Scholar]
- Vacchiano, G.; Garbarino, M.; Lingua, E.; Motta, R. Forest dynamics and disturbance regimes in the Italian Apennines. For. Ecol. Manag. 2017, 388, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Nocentini, S. Structure and management of beech (Fagus sylvatica L.) forests in Italy. iForest-Biogeosci. For. 2009, 2, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Costantini, E.A.C.; Dazzi, C. The Soils of Italy; Springer: Dordrecht, The Netherlands, 2013; ISBN 9400756429. [Google Scholar]
- McElhinny, C.; Gibbons, P.; Brack, C.; Bauhus, J. Forest and woodland stand structural complexity: Its definition and measurement. For. Ecol. Manag. 2005, 218, 1–24. [Google Scholar] [CrossRef]
- Pretzsch, H. Structural Diversity As a Result of silviculural operations. Lesnictvi-Forestry 1998, 44, 429–439. [Google Scholar]
- Latham, P.A.; Zuuring, H.R.; Coble, D.W. A method for quantifying vertical forest structure. For. Ecol. Manag. 1998, 104, 157–170. [Google Scholar] [CrossRef]
- Kutsch, W.L.; Wirth, C.; Kattge, J.; Nöllert, S.; Herbst, M.; Kappen, L. Old-Growth Forests: Function, Fate and Value; Wirth, C., Gleixner, G., Heimann, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 57–79. ISBN 978-3-540-92706-8. [Google Scholar]
- Shannon, C. A mathematical theory of communication. Bell Syst. Technol. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biolagical collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Tabacchi, G.; Di Cosmo, L.; Gasparini, P.; Morelli, S. Stima del Volume e Della Fitomassa Delle Principali Specie Forestali Italiane, Equazioni di Previsione, Tavole del Volume e Tavole Della Fitomassa Arborea Epigea; Consiglio per la Ricerca e la Sperimentazione in Agricoltura Unità di Ricerca per il Monitoraggio e la Pianificazione Forestale: Trento, Italy, 2011; ISBN 9788897081111.
- West, P.W. Stem Volume. In Tree and Forest Measurement; Springer: Berlin/Heidelberg, Germany, 2009; pp. 23–32. ISBN 978-3-540-95966-3. [Google Scholar]
- Hunter, M.L.J. Wildlife, Forests, and Forestry: Principles of Managing Forests for Biological Diversity; Prentice-Hall: Hoboken, NJ, USA, 1990. [Google Scholar]
- Cindolo, C.; Petriccione, B. Progetto Biosoil—Biodiversity. Valutazione Della Biodiversità Forestale Sulla Rete Sistematica di Livello I. Manuale Nazionale, Italia; Corpo Forestale dello Stato: Roma, Italy, 2006. [Google Scholar]
- Stokland, J.N. The coarse woody debris profile: An archive of recent history and an important biodiversity. Ecol. Bull. 2001, 49, 71–83. [Google Scholar] [CrossRef]
- Rouvinen, S.; Rautiainen, A.; Kouki, J. A relation between historical forest use and current dead woody material in a boreal protected old-growth forest in Finland. Silva Fenn. 2005, 39, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, C.H. Individual variation in acorn production by five species of southern Appalachian oaks. For. Ecol. Manag. 2000, 132, 199–210. [Google Scholar] [CrossRef]
- Molina, M.; Pardo-de-santayana, M.; Aceituno, L.; Morales, R.; Tardío, J. Fruit production of strawberry tree (Arbutus unedo L.) in two Spanish forests. Forestry 2011, 84, 419–429. [Google Scholar] [CrossRef]
- Potena, G.; Sammarone, L.; Posillico, M.; Romano, M.; Consalvo, M. Fruttificazione del Faggio (Fagus sylvatica) e delle Querce (Quercus cerris, Q. pubescens) nel Parco Nazionale d’Abruzzo, Lazio e Molise e Zona di Protezione Esterna Nel 2007; Ufficio Foreste Demaniali di Castel di Sangro: L’Aquila, Italy, 2008. [Google Scholar]
- Canfield, R.H. Application of the line interception method in sampling range vegetation. J. For. 1941, 39, 34–40. [Google Scholar]
- Bonham, C.D. Measurements for Terrestrial Vegetation, 2nd ed.; John Wiley and Sons: New York, NY, USA, 2013; ISBN 9780470972588. [Google Scholar]
- Alvarado, M. Habitat correlates of ant assemblages in different forests of the South Pannonian Plain. Tiscia 2000, 32, 35–42. [Google Scholar]
- Frank, S.C.; Steyaert, S.M.J.G.; Swenson, J.E.; Storch, I.; Kindberg, J.; Barck, H.; Zedrosser, A. A “clearcut” case? Brown bear selection of coarse woody debris and carpenter ants on clearcuts. For. Ecol. Manag. 2015, 348, 164–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, R.J.; Lindgren, B.S. The Fine Scale Physical Attributes of Coarse Woody Debris and Effects of Surrounding Stand Structure on Its Utilization by Ants (Hymenoptera: Formicidae) in British Columbia, Canada; General Technical Report SRS-93; Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2006; pp. 67–74.
- Zhao, Y.; Staudenmayer, J.; Coull, B.A.; Wand, M.P. General Design Bayesian Generalized Linear Mixed Models. Stat. Sci. 2006, 21, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Spiegelhalter, D.J.; Best, N.G.; Carlin, B.P.; van der Linde, A. Bayesian Measures of Model Complexity and fit. J. R. Stat. Soc. Ser. B Stat. Methodol. 2002, 64, 583–639. [Google Scholar] [CrossRef] [Green Version]
- Wakefield, J. Bayesian and Frequentist Regression Methods; Springer: New York, NY, USA, 2013; ISBN 1441909257. [Google Scholar]
- Wood, S.N. Generalized additive models: An introduction with R. Chapman and Hall/CRC. Texts Stat. Sci. 2006, 67, 391. [Google Scholar]
- Scheipl, F. spikeSlabGAM: Bayesian Variable Selection, Model Choice and Regularization for Generalized Additive Mixed Models in R. J. Stat. Softw. 2011, 43, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Ishwaran, H.; Rao, J.S. Spike and Slab Variable Selection: Frequentist and Bayesian Strategies. Ann. Stat. 2005, 33, 730–773. [Google Scholar] [CrossRef] [Green Version]
- Plummer, M. JAGS: A Program for Analysis of Bayesian Graphical Models Using Gibbs Sampling. In Proceedings of the 3rd International Workshop on Distributed Statistical Computing, Vienna, Austria, 20–22 March 2003. [Google Scholar]
- Meyer, P.; Schmidt, M. Accumulation of dead wood in abandoned beech (Fagus sylvatica L.) forests in northwestern Germany. For. Ecol. Manag. 2011, 261, 342–352. [Google Scholar] [CrossRef]
- Castagneri, D.; Garbarino, M.; Berretti, R.; Motta, R. Site and stand effects on coarse woody debris in montane mixed forests of Eastern Italian Alps. For. Ecol. Manag. 2010, 260, 1592–1598. [Google Scholar] [CrossRef] [Green Version]
- Scolastri, A.; Cancellieri, L.; Iocchi, M.; Cutini, M. Old coppice versus high forest: The impact of beech forest management on plant species diversity in central Apennines (Italy). J. Plant Ecol. 2017, 10, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Juchheim, J.; Annighöfer, P.; Ammer, C.; Calders, K.; Raumonen, P.; Seidel, D. How management intensity and neighborhood composition affect the structure of beech (Fagus sylvatica L.) trees. Trees Struct. Funct. 2017, 31, 1723–1735. [Google Scholar] [CrossRef]
- Abrahamson, W.G.; Layne, J.N. Long-term patterns of acorn production for five oak species in xeric Florida uplands. Ecology 2003, 84, 2476–2492. [Google Scholar] [CrossRef]
- Kamler, J.; Dobrovolný, L.; Drimaj, J.; Kadavý, J.; Kneifl, M.; Adamec, Z.; Knott, R.; Martiník, A.; Plhal, R.; Zeman, J.; et al. The impact of seed predation and browsing on natural sessile oak regeneration under different light conditions in an over-aged coppice stand. iForest-Biogeosci. For. 2016, 9, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Cutini, A.; Chianucci, F.; Giannini, T. Effetti del trattamento selvicolturale su caratteristiche della copertura, produzione di lettiera e di seme in cedui di faggio in conversione. Ann. CRA 2009, 36, 109–124. [Google Scholar]
- Pignatti, S. La faggeta. In I Boschi d’Italia. Sinecologia e Biodiversità; UTET: Torino, Italy, 1998; p. 673. [Google Scholar]
- Hein, S.; Dhôte, J.-F. Effect of species composition, stand density and site index on the basal area increment of oak trees (Quercus sp.) in mixed stands with beech (Fagus sylvatica L.) in northern France. Ann. For. Sci. 2006, 63, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Pignatti, G.; Terzuolo, P.G.; Varese, P.; Semerari, P.; Lombardi, V. Criteri per la definizione di tipi forestali nei boschi dell’ Appennino meridionale. Forest 2004, 1, 112–127. [Google Scholar] [CrossRef]
- Müllerová, J.; Hédl, R.; Szabó, P. Coppice abandonment and its implications for species diversity in forest vegetation. For. Ecol. Manag. 2015, 343, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Mangipane, L.S.; Belant, J.L.; Hiller, T.L.; Colvin, M.E.; Gustine, D.D.; Mangipane, B.A.; Hilderbrand, G.V. Influences of landscape heterogeneity on home-range sizes of brown bears. Mamm. Biol. 2018, 88, 1–7. [Google Scholar] [CrossRef]
- Mitchell, M.S.; Powell, R.A. Optimal use of resources structures home ranges and spatial distribution of black bears. Anim. Behav. 2007, 74, 219–230. [Google Scholar] [CrossRef]
- Mitchell, M.S.; Powell, R.A. A mechanistic home range model for optimal use of spatially distributed resources. Ecol. Modell. 2004, 177, 209–232. [Google Scholar] [CrossRef]
- Koenig, W.D.; Knops, J.M.H. Patterns of Annual Seed Production by Northern Hemisphere Trees: A Global Perspective. Am. Nat. 2000, 155, 59–69. [Google Scholar] [CrossRef]
- Bogdziewicz, M.; Steele, M.A.; Marino, S.; Crone, E.E. Correlated seed failure as an environmental veto to synchronize reproduction of masting plants. New Phytol. 2018, 219, 98–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garshelis, D.L.; Noyce, K.V. Seeing the World through the Nose of a Bear—Diversity of Foods Fosters Behavioral and Demographic Stability. In Wildlife Science Linking Ecological Theory and Management Applications; CRC Press Taylor & Francis Group: New York, NY, USA, 2007; pp. 139–164. [Google Scholar] [CrossRef]
- Olson, M.G.; Wolf, A.J.; Jensen, R.G. Influence of Forest Management on Acorn Production in the Southeastern Missouri Ozarks: Early Results of a Long-Term Ecosystem Experiment. Open J. For. 2015, 5, 568–583. [Google Scholar] [CrossRef] [Green Version]
- Reynolds-Hogland, M.J.; Mitchell, M.S.; Powell, R.A. Spatio-temporal availability of soft mast in clearcuts in the Southern Appalachians. For. Ecol. Manag. 2006, 237, 103–114. [Google Scholar] [CrossRef]
- Fredericksen, T.S.; Ross, B.D.; Hoffman, W.; Morrison, M.L.; Beyea, J.; Johnson, B.N.; Lester, M.B.; Ross, E. Short-term understory plant community responses to timber-harvesting intensity on non-industrial private forestlands in Pennsylvania. For. Ecol. Manag. 1999, 116, 129–139. [Google Scholar] [CrossRef]
- Saremi, H.; Kumar, L.; Turner, R.; Stone, C.; Melville, G. Impact of local slope and aspect assessed from LiDAR records on tree diameter in radiata pine (Pinus radiata D. Don) plantations. Ann. For. Sci. 2014, 71, 771–780. [Google Scholar] [CrossRef]
- Melini, D. A spatial model for sporadic tree species distribution in support of tree oriented silviculture. Ann. Silvic. Res. 2013, 37, 64–68. [Google Scholar]
- Raspe, O.; Findlay, C.; Jacquemart, L. Sorbus aucuparia L. J. Ecol. 2000, 910–930. [Google Scholar] [CrossRef]
- Pyttel, P.; Kunz, J.; Bauhus, J. Growth, regeneration and shade tolerance of the Wild Service Tree (Sorbus torminalis (L.) Crantz) in aged oak coppice forests. Trees Struct. Funct. 2013, 27, 1609–1619. [Google Scholar] [CrossRef]
- Paganová, V. Ecological requirements of wild service tree (Sorbus torminalis [L.] CRANTZ.) and service tree (Sorbus domestica L.) in relation with their utilizatiion in forestry and landscape. J. For. Sci. 2008, 54, 216–226. [Google Scholar] [CrossRef] [Green Version]
- Bernetti, G. Note sul trattamento delle fustaie di Faggio. Trattamento delle faggete in Italia: Dal metodo scientifico all’empirismo dei nostri giorni. I Georg. Quad. 2012, 2012-III, 7–11. [Google Scholar]
- Hertel, A.G.; Steyaert, S.M.J.G.; Zedrosser, A.; Mysterud, A.; Lodberg-Holm, H.K.; Gelink, H.W.; Kindberg, J.; Swenson, J.E. Bears and berries: Species-specific selective foraging on a patchily distributed food resource in a human-altered landscape. Behav. Ecol. Sociobiol. 2016, 70, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, K.K.; Kollmann, J. Defining the habitat niche of Sorbus torminalis from phytosociological relevés along a latitudinal gradient. Phytocoenologia 2004, 34, 639–662. [Google Scholar] [CrossRef]
- Piussi, P. Selvicoltura Generale; UTET: Torino, Italy, 1994; ISBN 880204869X.
- Perry, R.W.; Thill, R.E.; Peitz, D.G.; Tappe, P.A. Effect of different silvicultural systems on initial soft mast production. Wildl. Soc. Bull. 1999, 27, 915–923. [Google Scholar]
- Greenberg, C.H.; States, U.; Service, F.; Creek, B.; Forest, E.; Road, B. Fruit Production in Mature and Recently Regenerated Forests of the Appalachians. J. Wildl. Manag. 2007, 71, 321–335. [Google Scholar] [CrossRef]
- Greenberg, C.H.; Levey, D.J.; Kwit, C.; Mccarty, J.P.; Pearson, S.F.; Sargent, S.; Kilgo, J. Long-term patterns of fruit production in five forest types of the south carolina upper coastal plain. J. Wildl. Manag. 2012, 76, 1036–1046. [Google Scholar] [CrossRef]
- Spiecker, H.; Sebastian, H.; Makkonnen-Spiecker, K.; Thies, M. Valuable Broadleaved Forests in Europe; EFI Research Report 22; Brill: Leiden, The Netherlands; Boston, MA, USA; Köln, Germany, 2009; ISBN 978-90-04-16795-7. [Google Scholar]
- Manetti, M.C.; Becagli, C.; Sansone, D.; Pelleri, F. Tree-oriented silviculture: A new approach for coppice stands. iForest-Biogeosci. For. 2016, 9, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Pyttel, P.; Kunz, J.; Großmann, J. Growth of Sorbus torminalis after release from prolonged suppression. Trees Struct. Funct. 2019, 33, 1549–1557. [Google Scholar] [CrossRef]
- Warren, R.J. Mechanisms driving understory evergreen herb distributions across slope aspects: As derived from landscape position. Plant Ecol. 2008, 198, 297–308. [Google Scholar] [CrossRef]
- Gracia, M.; Montané, F.; Piqué, J.; Retana, J. Overstory structure and topographic gradients determining diversity and abundance of understory shrub species in temperate forests in central Pyrenees (NE Spain). For. Ecol. Manag. 2007, 242, 391–397. [Google Scholar] [CrossRef]
- Small, C.J.; McCarthy, B.C. Spatial and temporal variability of herbaceous vegetation in an eastern deciduous forest. Plant Ecol. 2002, 164, 37–48. [Google Scholar] [CrossRef]
- Horvat, V.; Biurrun, I.; García-Mijangos, I. Herb layer in silver fir—Beech forests in the western Pyrenees: Does management affect species diversity? For. Ecol. Manag. 2017, 385, 87–96. [Google Scholar] [CrossRef]
- Pinder, J.E.; Kroh, G.C.; White, J.D.; Basham May, A.M. The relationships between vegetation type and topography in Lassen Volcanic National Park. Plant Ecol. 1997, 131, 17–29. [Google Scholar] [CrossRef]
- Small, C.J.; McCarthy, B.C. Relationship of understory diversity to soil nitrogen, topographic variation, and stand age in an eastern oak forest, USA. For. Ecol. Manag. 2005, 217, 229–243. [Google Scholar] [CrossRef]
- Leuschner, C.; Lendzion, J. Air humidity, soil moisture and soil chemistry as determinants of the herb layer composition in European beech forests. J. Veg. Sci. 2009, 20, 288–298. [Google Scholar] [CrossRef]
- Hokkanen, P.J. Environmental patterns and gradients in the vascular plants and bryophytes of eastern Fennoscandian herb-rich forests. For. Ecol. Manag. 2006, 229, 73–87. [Google Scholar] [CrossRef]
- North, M.; Oakley, B.; Fiegener, R.; Gray, A.; Barbour, M. Influence of light and soil moisture on Sierran mixed-conifer understory communities. Plant Ecol. 2005, 177, 13–24. [Google Scholar] [CrossRef]
- Gálhidy, L.; Mihók, B.; Hagyó, A.; Rajkai, K.; Standovár, T. Effects of gap size and associated changes in light and soil moisture on the understorey vegetation of a Hungarian beech forest. Plant Ecol. 2006, 183, 133–145. [Google Scholar] [CrossRef]
- Gabriela Sonohat; Philippe Balandier; Felix Ruchaud Predicting solar radiation transmittance in the understory of even-aged coniferous stands in temperate forests. Ann. For. Sci. 2004, 61, 629–641. [CrossRef] [Green Version]
- Hale, S.E. The effect of thinning intensity on the below-canopy light environment in a Sitka spruce plantation. For. Ecol. Manag. 2003, 179, 341–349. [Google Scholar] [CrossRef]
- Page, L.M.; Cameron, A.D.; Clarke, G.C. Influence of overstorey basal area on density and growth of advance regeneration of Sitka spruce in variably thinned stands. For. Ecol. Manag. 2001, 151, 25–35. [Google Scholar] [CrossRef]
- Campione, M.A.; Nagel, L.M.; Webster, C.R. Herbaceous-Layer Community Dynamics along a Harvest-Intensity Gradient after 50 Years of Consistent Management. Open J. For. 2012, 02, 97–106. [Google Scholar] [CrossRef]
- Hofmeister, J.; Hosek, J.; Brabec, M.; Hedl, R.; Modry, M. Strong influence of long-distance edge effect on herb-layer vegetation in forest fragments in an agricultural landscape. Perspect. Plant Ecol. Evol. Syst. 2013, 15, 293–303. [Google Scholar] [CrossRef]
- Coll, L.; González-Olabarria, J.R.; Mola-Yudego, B.; Pukkala, T.; Messier, C. Predicting understory maximum shrubs cover using altitude and overstory basal area in different Mediterranean forests. Eur. J. For. Res. 2011, 130, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Vockenhuber, E.A.; Scherber, C.; Langenbruch, C.; Meiner, M.; Seidel, D.; Tscharntke, T. Tree diversity and environmental context predict herb species richness and cover in Germany’s largest connected deciduous forest. Perspect. Plant Ecol. Evol. Syst. 2011, 13, 111–119. [Google Scholar] [CrossRef]
- Decocq, G.; Aubert, M.; Dupont, F.; Alard, D.; Saguez, R.; Wattez-franger, A.; Foucault, B.D.E.; Delelis-dusollier, A. Plant diversity in a managed temperate deciduous forest: Understorey response to two silvicultural systems. J. Appl. Ecol. 2004, 41, 1065–1079. [Google Scholar] [CrossRef]
- Arnan, X.; Gracia, M.; Lluis, C.; Retana, J. Forest management conditioning ground ant community structure and composition in temperate conifer forests in the Pyrenees Mountains. For. Ecol. Manag. 2009, 258, 51–59. [Google Scholar] [CrossRef]
- Grevé, M.E.; Hager, J.; Weisser, W.W.; Schall, P.; Gossner, M.M.; Feldhaar, H. Effect of forest management on temperate ant communities. Ecosphere 2018, 9, e02303. [Google Scholar] [CrossRef]
- Noyce, K.V.; Kannowski, P.B.; Riggs, M.R. Black bears as ant-eaters: Seasonal associations between bear myrmecophagy and ant ecology in north-central Minnesota. Can. J. Zool. 1997, 75, 1671–1686. [Google Scholar] [CrossRef]
- Warren, R.J.; Bradford, M.A. Ant colonization and coarse woody debris decomposition in temperate forests. Insectes Soc. 2011, 59, 215–221. [Google Scholar] [CrossRef]
- King, J.R.; Warren, R.J.; Maynard, D.S.; Bradford, M.A. Ants: Ecology and Impacts in Dead Wood. In Saproxylic Insects: Diversity, Ecology and Conservation; Ulyshen, M.D., Ed.; Springer: Cham, Switzerland, 2018; pp. 237–262. ISBN 978-3-319-75937-1. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Forest Parameters | Rocca Di Mezzo | Lucoli | Balsorano | |||
|---|---|---|---|---|---|---|
| SD | SD | SD | ||||
| Basal area of trees (diameter ≥ 55 cm) (m2) | 1.8 | 3.1 | 1.4 | 4.7 | 3.1 | 4.9 |
| Basal Area (m2) | 33.2 | 7.6 | 33.5 | 8.9 | 30.9 | 9.2 |
| Basal area of trees (15 ≤ diameter ≤ 45 cm) (m2) | 19.4 | 9.4 | 21.3 | 10.3 | 19.2 | 11.2 |
| Tree volume (m3) | 258 | 125.2 | 249.1 | 120.7 | 272.6 | 145.7 |
| dq (cm) | 17.3 | 7.1 | 17.4 | 7.4 | 21.0 | 9.7 |
| Number of vertical strata (Latham index) | 3.4 | 1.4 | 2.9 | 0.8 | 2.8 | 0.9 |
| Number of stems | 1922 | 1353 | 2203 | 1649 | 1421 | 1094 |
| Number of stumps | 285 | 255.7 | 325.3 | 317.1 | 207.5 | 339.2 |
| Total volume of dead wood (m3) | 4.1 | 4.7 | 0.9 | 1.1 | 4.0 | 8.5 |
| Species richness (n) | 3.7 | 3.1 | 1.8 | 1.2 | 2.7 | 2.2 |
| Trees Shannon index | 0.5 | 0.6 | 0.1 | 0.3 | 0.4 | 0.5 |
| Code | Description |
|---|---|
| Food abundance | |
| BAhm | Basal area of hard-mast-producing species |
| BAsm | Basal area of soft-mast-producing species |
| COVherb | Cover of herbaceous vegetation |
| COVhm | Cover of hard-mast-producing species |
| COVshr | Shrub cover |
| COVsm | Cover of soft-mast-producing species |
| DEAD | Total volume of dead wood |
| VOLhm | Tree volume of hard-mast-producing species |
| VOLsm | Tree volume of soft-mast-producing species |
| Forest structure | |
| BA | Basal area |
| BA45 | Basal area of trees with a diameter of between 15 and 45 cm |
| BA55 | Basal area of trees with a diameter greater than 55 cm |
| COV | Canopy cover |
| COVgap | Gap (% of cover) |
| DEAD | Total volume of dead wood |
| DECn | Volume of decay class n |
| DECd | Number of decay classes |
| dq | Quadratic mean diameter |
| LCWPd | Volume of lying dead wood pieces |
| LOGd | Volume of logs |
| R | Tree species richness |
| SDdbh | Standard deviation of dbh |
| SH | Tree Shannon index |
| SNAGd | Volume of snags |
| STAd | Volume of standing dead trees |
| STE | Number of stems |
| STO | Number of stumps |
| STUd | Volume of dead stumps |
| VERLat | Number of vertical strata (Latham index) |
| VERPre | Index of vertical profile (Pretzsch) |
| VOL | Tree volume |
| Topographic variables | |
| ALT | Altitude |
| SLO | Slope |
| X; Y | Coordinates of the center of the plot (longitude; latitude) |
| Code | Description |
|---|---|
| Proxies of bear key foods | |
| ANTS | Presence/absence of ants |
| WIL | Presence/absence of wild ungulates |
| Forest structure | |
| SYS | Forest system |
| SYS0 | High forest |
| SYS1 | Coppice |
| TYP | Forest type |
| TYP0 | Oak and hornbeam forest |
| TYP1 | Beech forest |
| Topographic variables | |
| EXP | Exposure |
| SOI | Type of soil |
| SOI1 | Limestone substrates and moraine deposits |
| SOI2 | Mainly karst faces. Limestone and residual substrates |
| SOI3 | Faces with incision valleys on calcareous substrate |
| SOI4 | Summit faces with glacial cirques. Limestone substrates |
| X; Y | Coordinates of the center of the plot (latitude; longitude) |
| Code | Description |
|---|---|
| Y | Latitude |
| EXPN | Northern exposure |
| EXPS | Southern exposure |
| EXPW | Western exposure |
| SLO | Slope (degree) |
| ALT | Altitude (m) |
| BA | Basal area (m2/ha) |
| TYP0 | Forest type |
| VERLat | Number of vertical strata (Latham index) |
| STU | Number of stumps |
| SYS | Forest system management |
| DECd | Number of decay classes |
| dq | Quadratic mean diameter (cm) |
| Parameter | Posterior Mean | Lower 95% CI | Lower 95% CI | |
|---|---|---|---|---|
| Hard mast | Y | 1.26 | 0.07 | 2.45 |
| SYS1 | −3.05 | −5.37 | −0.73 | |
| TYP | −8.59 | −11.67 | −5.53 | |
| STU | −0.006 | −0.01 | −0.001 | |
| DECd | −0.71 | −1.35 | −0.08 | |
| Soft mast | SLO | −0.01 | −0.02 | −0.001 |
| Green vegetation | Y | −4.90 | −7.37 | −2.43 |
| EXPS | 11.37 | 4.07 | 18.69 | |
| EXPN | 8.61 | 1.52 | 15.69 | |
| SLO | −0.34 | −0.58 | −0.11 | |
| BA | −0.58 | −0.84 | −0.32 | |
| Deadwood | DECd | 1.68 | 0.89 | 2.47 |
| BAL | LUC | RdM | |
|---|---|---|---|
| Acer campestre | 4 | 4 | 3 |
| Acer opalus | 52 | 5 | 91 |
| Acer pseudoplatanus | 12 | 1 | 0 |
| Carpinus betulus | 0 | 0 | 46 |
| Carpinus orientalis | 358 | 0 | 0 |
| Corylus avellana * | 0 | 0 | 1 |
| Crataegus monogyna | 0 | 0 | 1 |
| Fagus sylvestris * | 824 | 2073 | 1056 |
| Fraxinus ornus | 146 | 52 | 191 |
| Ilex aquifolium | 0 | 0 | 1 |
| Juniperus | 4 | 0 | 0 |
| Laburnum anagyroides | 1 | 2 | 4 |
| Malus spp. * | 0 | 0 | 0 |
| Ostrya carpinifolia | 216 | 45 | 396 |
| Populus tremula | 0 | 0 | 4 |
| Pyrus spp. * | 0 | 0 | 0 |
| Quercus cerris * | 15 | 12 | 161 |
| Quercus ilex * | 65 | 0 | 0 |
| Quercus pubescens * | 72 | 3 | 10 |
| Salix caprea | 0 | 0 | 1 |
| Sorbus aria * | 0 | 8 | 1 |
| Sorbus aucuparia * | 0 | 3 | 1 |
| Sorbus torminalis * | 0 | 0 | 7 |
| Tilia spp. | 0 | 0 | 15 |
| Ulmus glabra | 0 | 0 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rositi, A.A.; Jona Lasinio, G.; Ciucci, P. Assessing Forest Structural and Topographic Effects on Habitat Productivity for the Endangered Apennine Brown Bear. Forests 2021, 12, 916. https://doi.org/10.3390/f12070916
Rositi AA, Jona Lasinio G, Ciucci P. Assessing Forest Structural and Topographic Effects on Habitat Productivity for the Endangered Apennine Brown Bear. Forests. 2021; 12(7):916. https://doi.org/10.3390/f12070916
Chicago/Turabian StyleRositi, Angela Anna, Giovanna Jona Lasinio, and Paolo Ciucci. 2021. "Assessing Forest Structural and Topographic Effects on Habitat Productivity for the Endangered Apennine Brown Bear" Forests 12, no. 7: 916. https://doi.org/10.3390/f12070916
APA StyleRositi, A. A., Jona Lasinio, G., & Ciucci, P. (2021). Assessing Forest Structural and Topographic Effects on Habitat Productivity for the Endangered Apennine Brown Bear. Forests, 12(7), 916. https://doi.org/10.3390/f12070916