
Effects of Planting Density on Morphological and Photosynthetic Characteristics of Leaves in Different Positions on Cunninghamia lanceolata Saplings

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Experiment Design
2.2. Measurements of Leaf Area and Dry Mass
2.3. Measurement of Leaf Gas Exchange Attributes
2.4. Measurement of Photosynthetic Pigment Contents
2.5. Statistical Analysis
3. Results
3.1. Interactions between Planting Density and Leaf Position
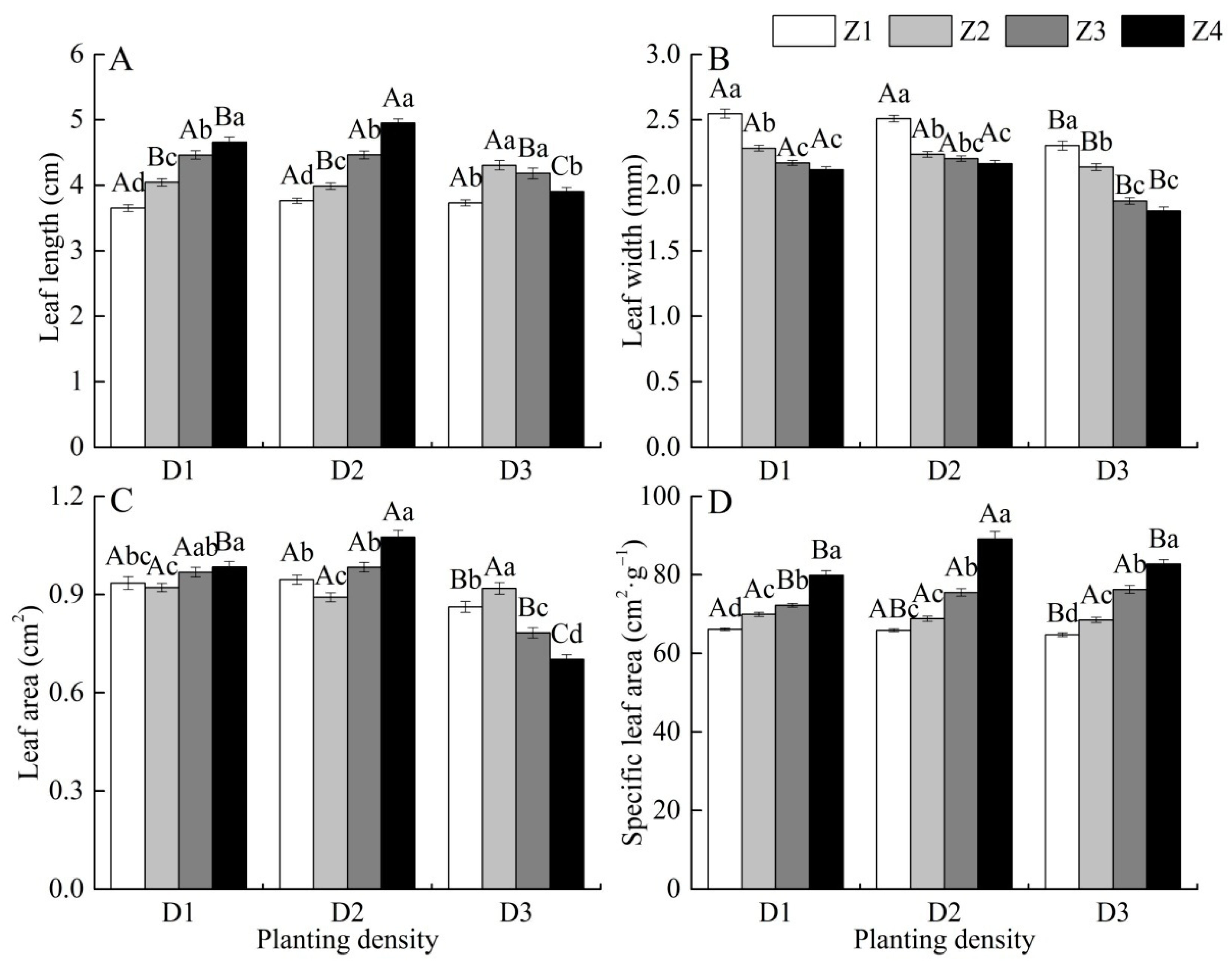
3.2. Effects of Planting Density on Leaf Morphology
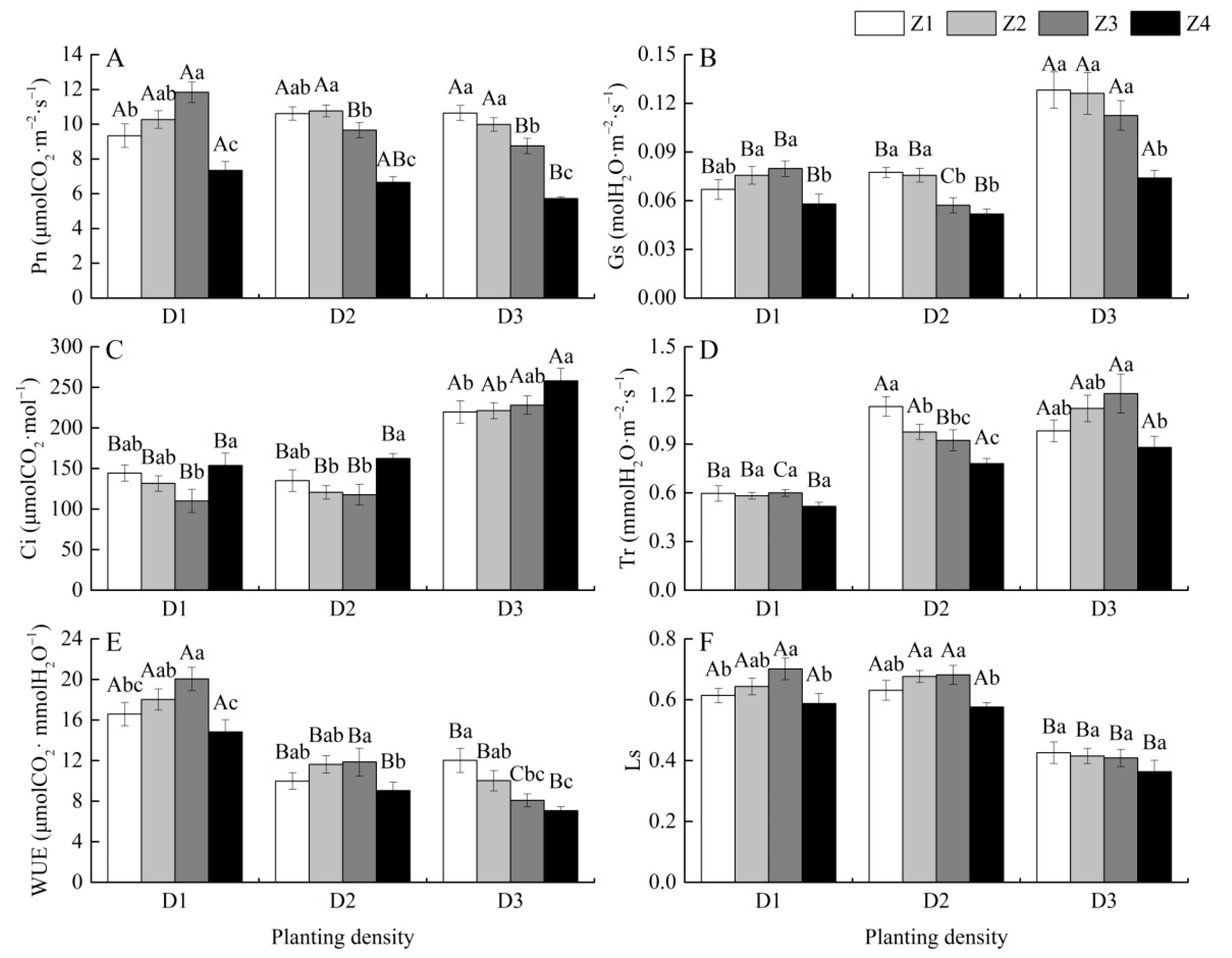
3.3. Effects of Planting Density on Leaf Gas Exchange Parameters
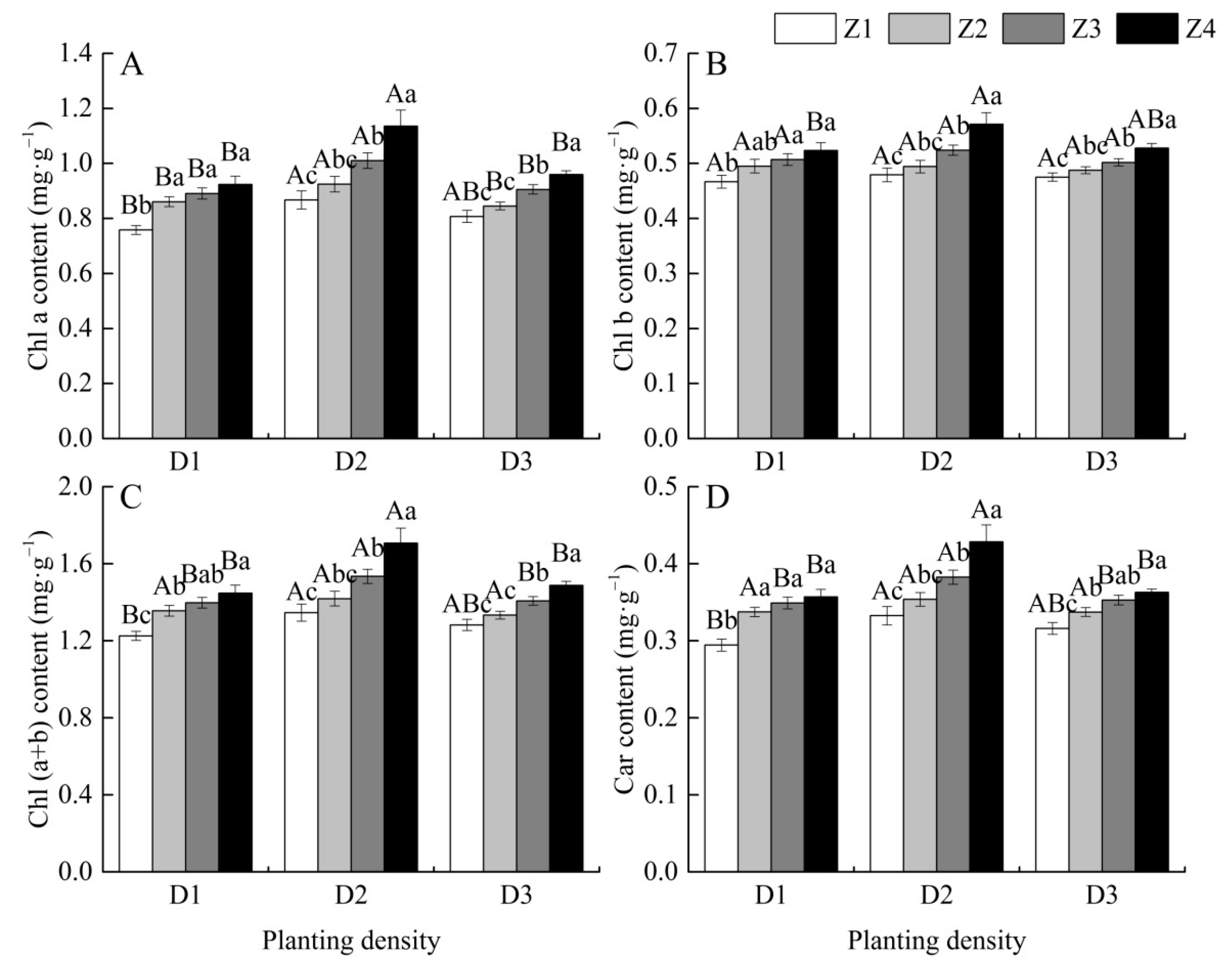
3.4. Effects of Planting Density on Photosynthetic Pigment Contents
3.5. Phenotypic Plasticity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jafari, T.; Rahikainen, M.; Puljula, E.; Sinkkonen, J.; Kangasjärvi, S. The impact of light intensity on metabolomic profile of Arabidopsis thaliana wild type and reticulata mutant by NMR spectroscopy. Phytochem. Lett. 2018, 26, 170–178. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef]
- Zhang, S.R.; Ma, K.P.; Chen, L.Z. Response of photosynthetic plasticity of Paeoniasuffruticosa to changed light environments. Environ. Exp. Bot. 2003, 49, 121–133. [Google Scholar] [CrossRef]
- Harrington, T.B.; Harrington, C.A.; DeBell, D.S. Effects of planting spacing and site quality on 25-year growth and mortality relationships of Douglas-fir (Pseudotsugamenziesii var. menziesii). For. Ecol. Manag. 2009, 258, 18–25. [Google Scholar] [CrossRef]
- Rodney, W.; Thomas, H.; Thomas, L.; Randal, H.; Robert, H. Effects of planting density and seed source on loblolly pine stands in southeastern Oklahoma. For. Sci. 2010, 56, 437–443. [Google Scholar]
- Moreno, G.; Cubera, E. Impact of stand density on water status and leaf gas exchange in Quercus ilex. For. Ecol. Manag. 2008, 254, 74–84. [Google Scholar] [CrossRef]
- Dai, Y.J.; Shen, Z.G.; Liu, Y.; Wang, L.L.; Hannaway, D.; Lu, H.F. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigmahemsleyanum Diels et Gilg. Environ. Exp. Bot. 2009, 65, 177–182. [Google Scholar] [CrossRef]
- Tang, H.; Hu, Y.Y.; Yu, W.W.; Song, L.L.; Wu, J.S. Growth, photosynthetic and physiological responses of Torreya grandis seedlings to varied light environments. Trees. 2015, 29, 1011–1022. [Google Scholar] [CrossRef]
- Pearcy, R.W. Responses of plants to heterogeneous light environments. In Handbook of Functional Plant Ecology; Dekker, M., Ed.; CRC Press: New York, NY, USA, 2007; pp. 213–258. [Google Scholar]
- Gandolfi, S.; Joly, C.A.; Leitão Filho, H.F. “Gaps of deciduousness”: Cyclical gaps in tropical forests. Sci. Agric. 2009, 66, 280–284. [Google Scholar] [CrossRef]
- Vemmos, S.N.; Petri, E.; Stournaras, V. Seasonal changes in photosynthetic activity and carbohydrate content in leaves and fruit of three fig cultivars (Ficuscarica L.). Sci. Hortic. Amst. 2013, 160, 198–207. [Google Scholar] [CrossRef]
- Chen, F.S.; Zeng, D.H.; Fahey, T.J.; Yao, C.Y.; Yu, Z.Y. Response of leaf anatomy of Chenopodium acuminatum to soil resource availability in a semi-arid grassland. Plant Ecol. 2010, 209, 375–382. [Google Scholar] [CrossRef]
- Xia, J.B.; Lang, Y.; Zhao, Q.K.; Liu, P.; Su, L. Photosynthetic characteristics of Tamarixchinensis under different groundwater depths in freshwater habitats. Sci. Total Environ. 2020, 761, 143221. [Google Scholar] [CrossRef]
- Guo, Q.X.; Song, H.F.; Kang, J.Y.; Korpelainen, H.; Li, C.Y. Different responses in leaf-level physiology to competition and facilitation under different soil types and N fertilization. Environ. Exp. Bot. 2018, 150, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.D.; Wang, X.; Gao, H.Y.; Shi, L.; Chow, W.S. Systemic regulation of leaf anatomical structure, photosynthetic performance, and high-light tolerance in sorghum. Plant Physiol. 2011, 155, 1416–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhusal, N.; Bhusal, S.J.; Yoon, T.M. Comparisons of physiological and anatomical characteristics between two cultivars in bi-leader apple trees (Malus x domestica Borkh.). Sci. Hortic. Amst. 2018, 231, 73–81. [Google Scholar] [CrossRef]
- Santos, V.; Ferreira, M. Are photosynthetic leaf traits related to the first-year growth of tropical tree seedlings? A light-induced plasticity test in a secondary forest enrichment planting. For. Ecol. Manag. 2020, 460, 117900. [Google Scholar] [CrossRef]
- Carlson, J.E.; Adams, C.A.; Holsinger, K.E. Intraspecific variation in stomatal traits, leaf traits and physiology reflects adaptation along aridity gradients in a South African shrub. Ann. Bot. 2016, 117, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.Q.; Huang, Z.J.; Wang, Z.N.; Chen, Y.F.; Wen, Z.M.; Liu, B.; Tigabu, M. Responses of leaf morphology, NSCs contents and C:N:P stoichiometry of Cunninghamia lanceolata and Schima superba to shading. BMC Plant Biol. 2020, 20, 386–403. [Google Scholar]
- SFA. Reports of Chinese Forestry Resource: The 8th National Forest Resource Inventory; Chinese Forestry Publishing House: Beijing, China, 2013; p. 16. [Google Scholar]
- Duan, A.G.; Lei, J.; Hu, X.Y.; Zhang, J.G.; Du, H.L.; Zhang, X.Q.; Guo, W.F.; Sun, J.J. Effects of planting density on soil bulk density, pH and nutrients of unthinned Chinese fir mature stands in south subtropical region of China. Forests. 2019, 10, 351. [Google Scholar] [CrossRef] [Green Version]
- Xiang, C.W.; Zhang, J.G.; Duan, A.G.; Zhang, X.Q. Effects of planting density and site quality on stand volume of Chinese fir plantation. For. Res. 2014, 27, 801–808. [Google Scholar]
- Wang, C.Q.; Xue, L.; Dong, Y.H.; Hou, L.Y.; Wei, Y.H.; Jiao, R.Z. Responses of soil microbial community structure to stand densities of Chinese fir plantations. J. For. Res. 2019, 24, 162–167. [Google Scholar] [CrossRef]
- Wang, C.Q.; Xue, L.; Jiao, R.Z. Soil phosphorus fractions, phosphatase activity, and the abundance of pho C and pho D genes vary with planting density in subtropical Chinese fir plantations. Soil Till. Res. 2021, 209, 104946. [Google Scholar] [CrossRef]
- Modrzyński, J.; Chmura, D.J.; Tjoelker, M.G.; Thomas, S. Seedling growth and biomass allocation in relation to leaf habit and shade tolerance among 10 temperate tree species. Tree Physiol. 2015, 35, 879–893. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.M.; Wang, L.; Wei, Y.; Zhang, Z.H.; Zhu, H.; Kong, L.G.; Meng, S.Y.; Song, C.D.; Wang, H.T.; Ma, F.Y. Irrigation with magnetically treated saline water influences the growth and photosynthetic capability of Vitis vinifera L. seedlings. Sci. Hortic. Amst. 2020, 262, 109056. [Google Scholar] [CrossRef]
- Li, H.S. Principles and Techniques of Plant Physiological and Biochemical Experimental; Higher Education Press: Beijing, China, 2000; pp. 134–137. [Google Scholar]
- Kramer-Walter, K.R.; Laughlin, D.C. Root nutrient concentration and biomass allocation are more plastic than morphological traits in response to nutrient limitation. Plant Soil. 2017, 416, 539–550. [Google Scholar] [CrossRef]
- Ku, L.X.; Ren, Z.Z.; Chen, X.; Shi, Y.; Qi, J.S.; Su, H.H.; Wang, Z.Y.; Li, G.H.; Wang, X.B.; Zhu, Y.G.; et al. Genetic analysis of leaf morphology underlying the plant density response by QTL mapping in maize (Zea mays L.). Mol. Breed. 2016, 36, 63. [Google Scholar] [CrossRef]
- Bhusal, N.; Kim, H.S.; Han, S.G.; Yoon, T.M. Photosynthetic traits and plant–water relations of two apple cultivars grown as bi-leader trees under long-term waterlogging conditions. Environ. Exp. Bot. 2020, 176, 1041. [Google Scholar] [CrossRef]
- Granata, M.U.; Bracco, F.; Nola, P.; Catoni, R. Photosynthetic characteristic and leaf traits variations along a natural light gradient in Acer campestre and Crataegus monogyna. Flora 2020, 268, 151626. [Google Scholar] [CrossRef]
- Khan, M.N.I.; Khatun, S.; Azad, M.S.; Mollick, A.S. Leaf morphological and anatomical plasticity in Sundri (Heritiera fomes Buch.-Ham.) along different canopy light and salinity zones in the Sundarbans mangrove forest, Bangladesh. Glob. Ecol. Conserv. 2020, 23, 1–15. [Google Scholar] [CrossRef]
- Khan, M.N.I.; Suwa, R.; Hagihara, A.; Ogawa, K. Interception of photosynthetic photon flux density in a mangrove stand of Kandeliacandel (L.) Druce. J. For. Res. 2004, 9, 205–210. [Google Scholar] [CrossRef]
- Klich, M.G. Leaf variations in Elaeagnus angustifolia related to environmental heterogeneity. Environ. Exp. Bot. 2000, 44, 171–183. [Google Scholar] [CrossRef]
- Nascimento, K.C.; Pastorini, L.H.; Romagnolo, M.B.; de Souza, L.A. Do Eugenia hiemalis seedling leaves under different light conditions develop phenotypic plasticity? Plant Ecol. 2015, 216, 1571–1581. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Q.Q.; Daryanto, S.; Guo, S.; Huang, Z.J.; Wang, Z.N.; Wang, L.X.; Ma, X.Q. Responses of Chinese fir and Schimasuperba seedlings to light gradients: Implications for the restoration of mixed broadleaf-conifer forests from Chinese fir monocultures. For. Ecol. Manag. 2018, 419–420, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.Q. Photosynthetic Efficiency; Shanghai science and Technology Press: Shanghai, China, 2002. [Google Scholar]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Gutiérrez, E.; Pérez, P.; Morcuende, R.; Verdejo, A.L.; Martinez-Carrasco, R. Acclimation to future atmospheric CO2 levels increases photochemical efficiency and mitigates photochemistry inhibition by warm temperatures in wheat under field chambers. Physiol. Plant. 2009, 137, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.H.; Song, L.L.; Yu, W.W.; Hu, Y.Y.; Liu, Y.; Wu, J.S.; Ying, Y.Q. Growth, physiological, and biochemical responses of Camptothecaacuminata seedlings to different light environments. Front. Plant Sci. 2015, 6, 321. [Google Scholar] [CrossRef] [Green Version]
- Hallik, L.; Niinemets, U.; Kull, O. Photosynthetic acclimation to light in woody and herbaceous species: A comparison of leaf structure, pigment content and chlorophyll fluorescence characteristics measured in the field. Plant Biol. 2012, 14, 88–99. [Google Scholar] [CrossRef]
- Shao, M.J.; Liu, W.K.; Zha, L.Y.; Zhou, C.B.; Zhang, Y.B.; Li, B.S. Altering light-dark cycle at pre-harvest stage regulated growth, nutritional quality, and photosynthetic pigment content of hydroponic lettuce. Acta Physiol. Plant. 2021, 43, 9. [Google Scholar] [CrossRef]
- Shao, Q.S.; Wang, H.Z.; Guo, H.P.; Zhou, A.C.; Huang, Y.Q.; Sun, Y.L.; Li, M.Y. Effects of shade treatments on photosynthetic characteristics, chloroplast ultrastructure, and physiology of Anoectochilus roxburghii. PLoS ONE. 2014, 9, e85996. [Google Scholar] [CrossRef]
- Griffin, J.J.; Ranney, T.G.; Pharr, D.M. Photosynthesis, chlorophyll fluorescence, and carbohydrate content of illicium taxa grown under varied irradiance. J. Am. Soc. Hort. Sci. 2004, 129, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Fontana, M.; Collin, A.; Courchesne, F.; Labrecque, M.; Bélanger, N. Root system architecture of Salix miyabeana “SX67” and relationships with aboveground biomass yields. Bio. Energy Res. 2020, 13, 183–196. [Google Scholar] [CrossRef]
- Fagundes, M.D.; Dalmolin, A.C.; Lobo, L.S.; Schilling, A.C.; dos Santos, M.S.; Mielke, M.S. Growth and phenotypic plasticity of two tropical tree species under low light availability. J. Plant Ecol. 2021, 14, 270–279. [Google Scholar] [CrossRef]
- Limón, F.; González-Astorga, J.; Nicolalde-Morejón, F.; Guevara, R. Phenotypic variation of Zamia loddigesii Miq. and Z. prasina W. Bull. (Zamiaceae, Cycadales): The effect of environmental heterogeneity. Plant Syst. Evol. 2016, 302, 1395–1404. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group | Trait | F-Value | Model R2 | ||
|---|---|---|---|---|---|
| Planting Density | Leaf Position | Planting Density × Leaf Position | |||
| Morphology | Leaf length | 18.776 *** | 96.263 *** | 24.772 *** | 0.308 |
| Leaf width | 121.998 *** | 158.859 *** | 5.542 *** | 0.414 | |
| Leaf area | 111.154 *** | 0.221 ns | 30.034 *** | 0.274 | |
| Specific leaf area | 9.284 *** | 225.683 *** | 7.945 *** | 0.410 | |
| Gas exchange parameters | Net photosynthetic rate | 4.450 ** | 49.233 *** | 4.962 *** | 0.477 |
| Stomatal conductance | 50.285 *** | 12.840 *** | 2.835 ** | 0.433 | |
| Intercellular CO2 concentration | 88.031 *** | 6.326 *** | 0.666ns | 0.494 | |
| Transpiration rate | 66.897 *** | 6.286 *** | 2.729 ** | 0.453 | |
| Water use efficiency | 74.294 *** | 6.091 *** | 2.676 ** | 0.473 | |
| Limiting value of stomata | 86.676 *** | 5.064 ** | 0.706 ns | 0.486 | |
| Photosynthetic pigment content | Chl a | 23.674 *** | 27.126 *** | 1.564 ns | 0.590 |
| Chl b | 3.575 * | 18.307 *** | 0.992 ns | 0.415 | |
| Car | 18.072 *** | 25.563 *** | 1.972 ns | 0.565 | |
| Chl (a + b) | 17.422 *** | 26.706 *** | 1.512 ns | 0.564 | |
| Group | Trait | Z1 | Z2 | Z3 | Z4 | Average CV |
|---|---|---|---|---|---|---|
| Gas exchange parameters | Net photosynthetic rate | 40.56 | 40.36 | 51.01 | 48.07 | 45.00 |
| Water use efficiency | 45.78 | 45.83 | 42.63 | 35.79 | 42.51 | |
| Intercellular CO2 concentration | 38.86 | 37.72 | 50.16 | 37.59 | 41.08 | |
| Transpiration rate | 37.09 | 36.35 | 45.84 | 34.04 | 38.33 | |
| Limiting value of stomata | 28.51 | 26.71 | 31.63 | 31.75 | 29.65 | |
| Stomatal conductance | 21.65 | 17.12 | 24.22 | 24.71 | 21.93 | |
| Morphology | Leaf area | 18.05 | 15.33 | 18.62 | 25.12 | 19.28 |
| Leaf length | 11.63 | 14.27 | 15.24 | 17.13 | 14.57 | |
| Leaf width | 12.85 | 10.24 | 12.06 | 14.59 | 12.43 | |
| Specific leaf area | 5.89 | 8.35 | 11.10 | 17.09 | 10.61 | |
| Photosynthetic pigment content | Chl a | 10.45 | 7.96 | 9.04 | 14.54 | 10.50 |
| Car | 10.02 | 6.40 | 7.53 | 13.64 | 9.40 | |
| Chl (a + b) | 8.52 | 6.79 | 7.36 | 12.29 | 8.74 | |
| Chl b | 6.62 | 6.11 | 5.48 | 9.09 | 6.82 | |
| Leaf position average | 21.18 | 19.97 | 23.71 | 23.96 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Z.; Liu, Q.; An, B.; Wu, X.; Sun, L.; Wu, P.; Liu, B.; Ma, X. Effects of Planting Density on Morphological and Photosynthetic Characteristics of Leaves in Different Positions on Cunninghamia lanceolata Saplings. Forests 2021, 12, 853. https://doi.org/10.3390/f12070853
Huang Z, Liu Q, An B, Wu X, Sun L, Wu P, Liu B, Ma X. Effects of Planting Density on Morphological and Photosynthetic Characteristics of Leaves in Different Positions on Cunninghamia lanceolata Saplings. Forests. 2021; 12(7):853. https://doi.org/10.3390/f12070853
Chicago/Turabian StyleHuang, Zhijun, Qingqing Liu, Bo An, Xiaojian Wu, Linjun Sun, Pengfei Wu, Bo Liu, and Xiangqing Ma. 2021. "Effects of Planting Density on Morphological and Photosynthetic Characteristics of Leaves in Different Positions on Cunninghamia lanceolata Saplings" Forests 12, no. 7: 853. https://doi.org/10.3390/f12070853
APA StyleHuang, Z., Liu, Q., An, B., Wu, X., Sun, L., Wu, P., Liu, B., & Ma, X. (2021). Effects of Planting Density on Morphological and Photosynthetic Characteristics of Leaves in Different Positions on Cunninghamia lanceolata Saplings. Forests, 12(7), 853. https://doi.org/10.3390/f12070853