
Recovery of Forest Structure Following Large-Scale Windthrows in the Northwestern Amazon

 ,
, 
 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
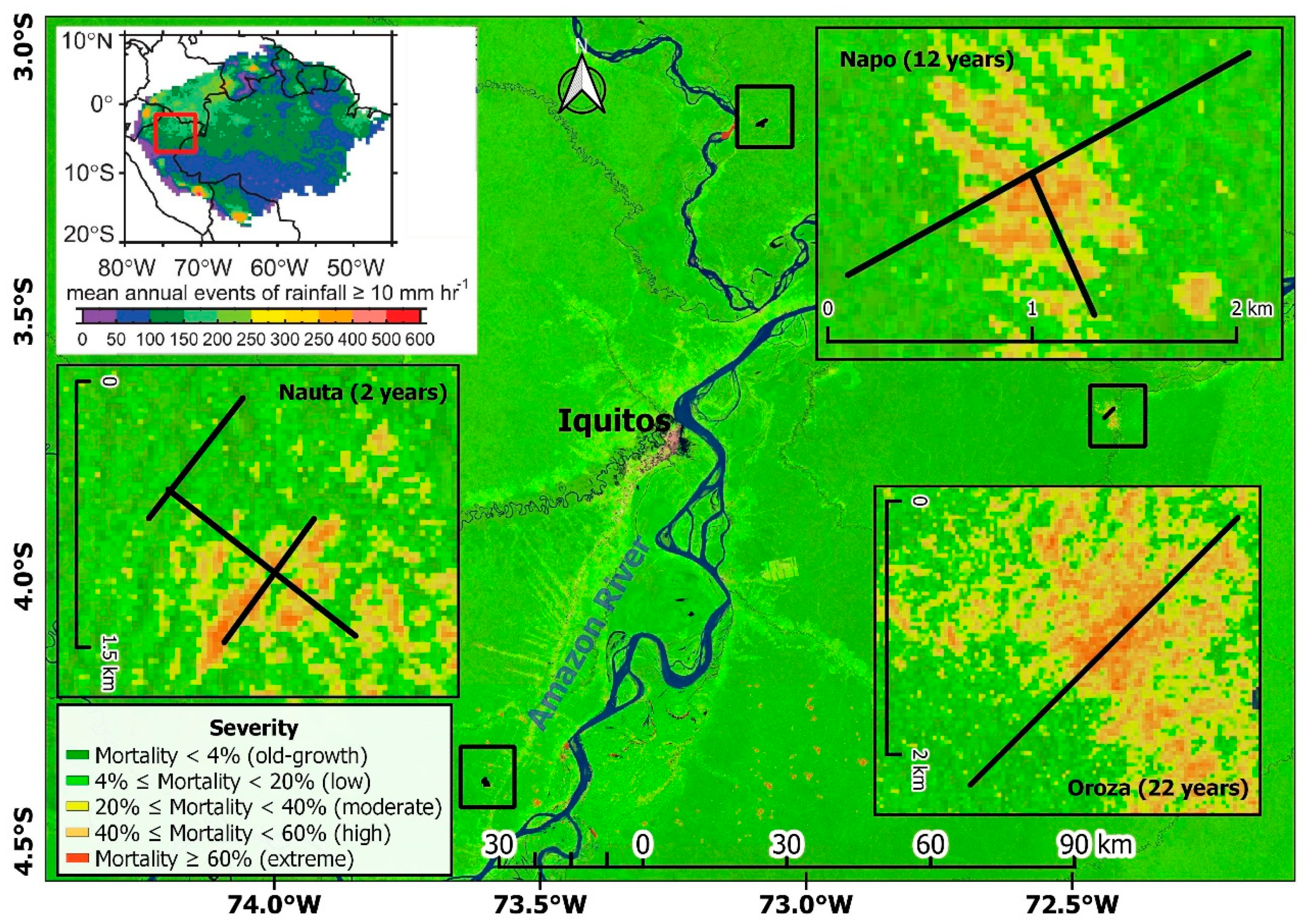
2.1. Study Area
2.2. Assessing Windthrow Severity
2.3. Forest Inventory
2.4. Data Processing and Analysis
2.5. Statistical Analysis
3. Results
3.1. Subplot Distribution by Disturbance Severity Class
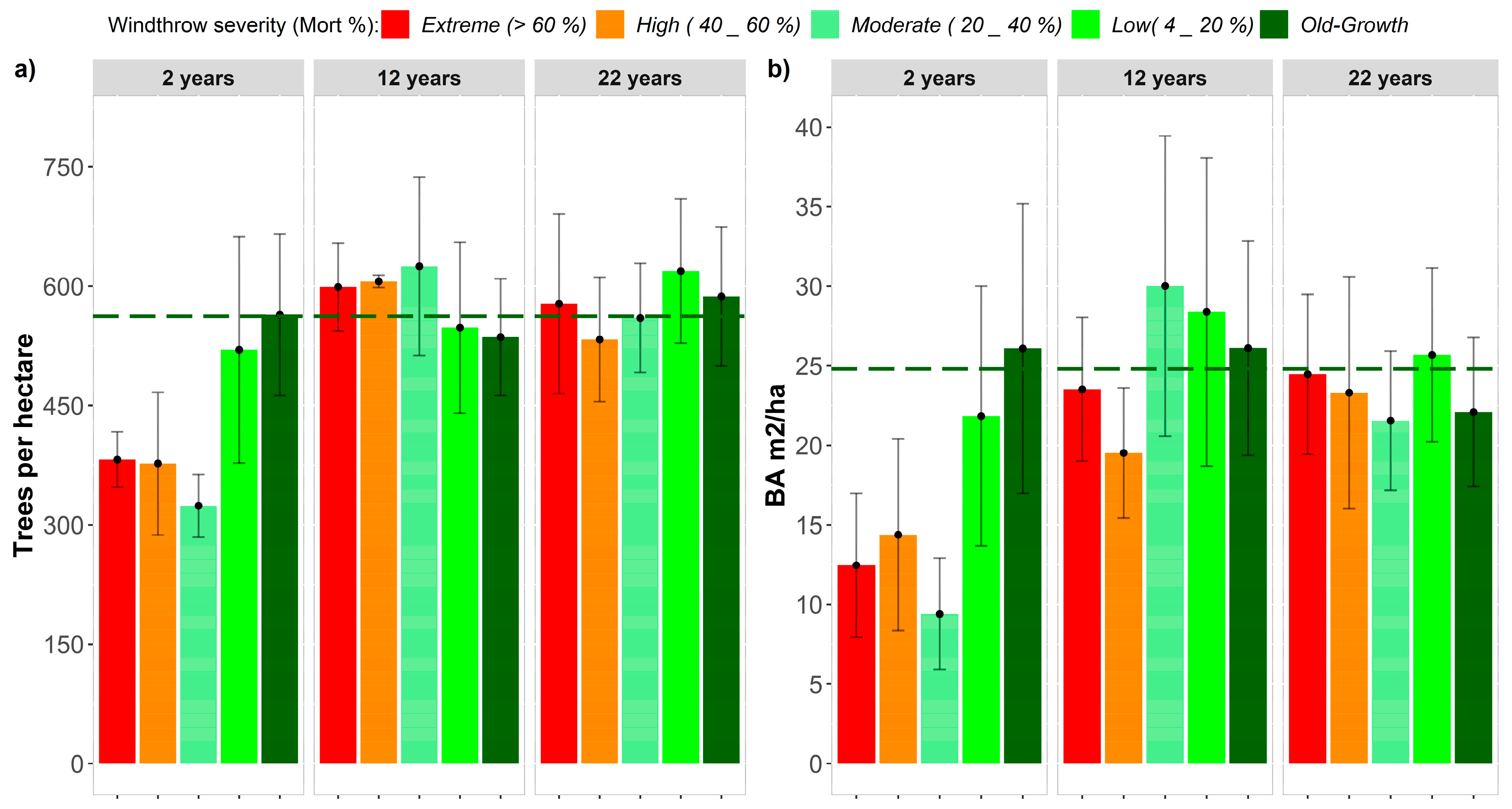
3.2. Structural Attributes of Old-Growth Forests
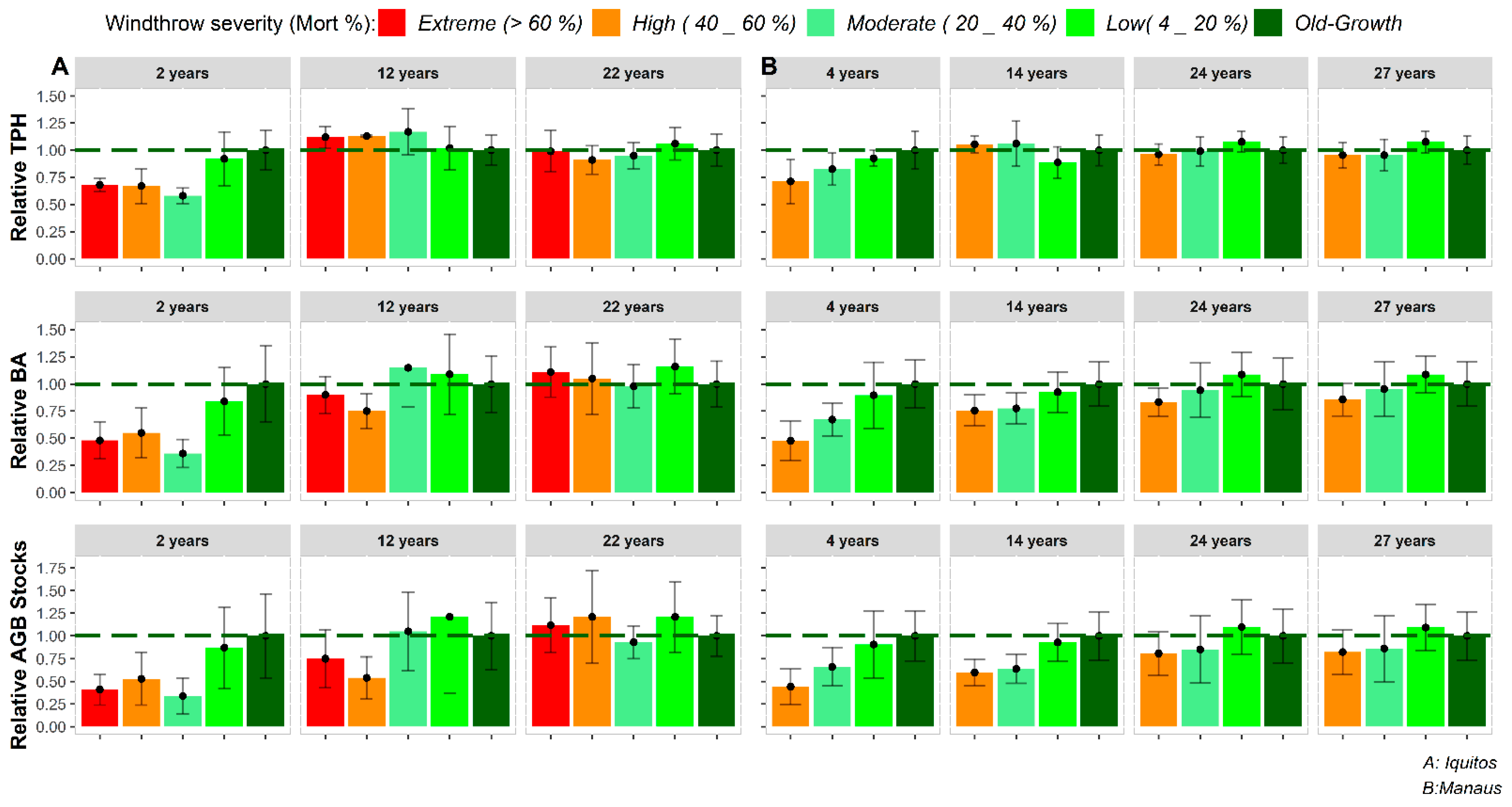
3.3. Recovery Patterns of Structural Attributes
3.3.1. Mean Diameter, Tree Density and Basal Area
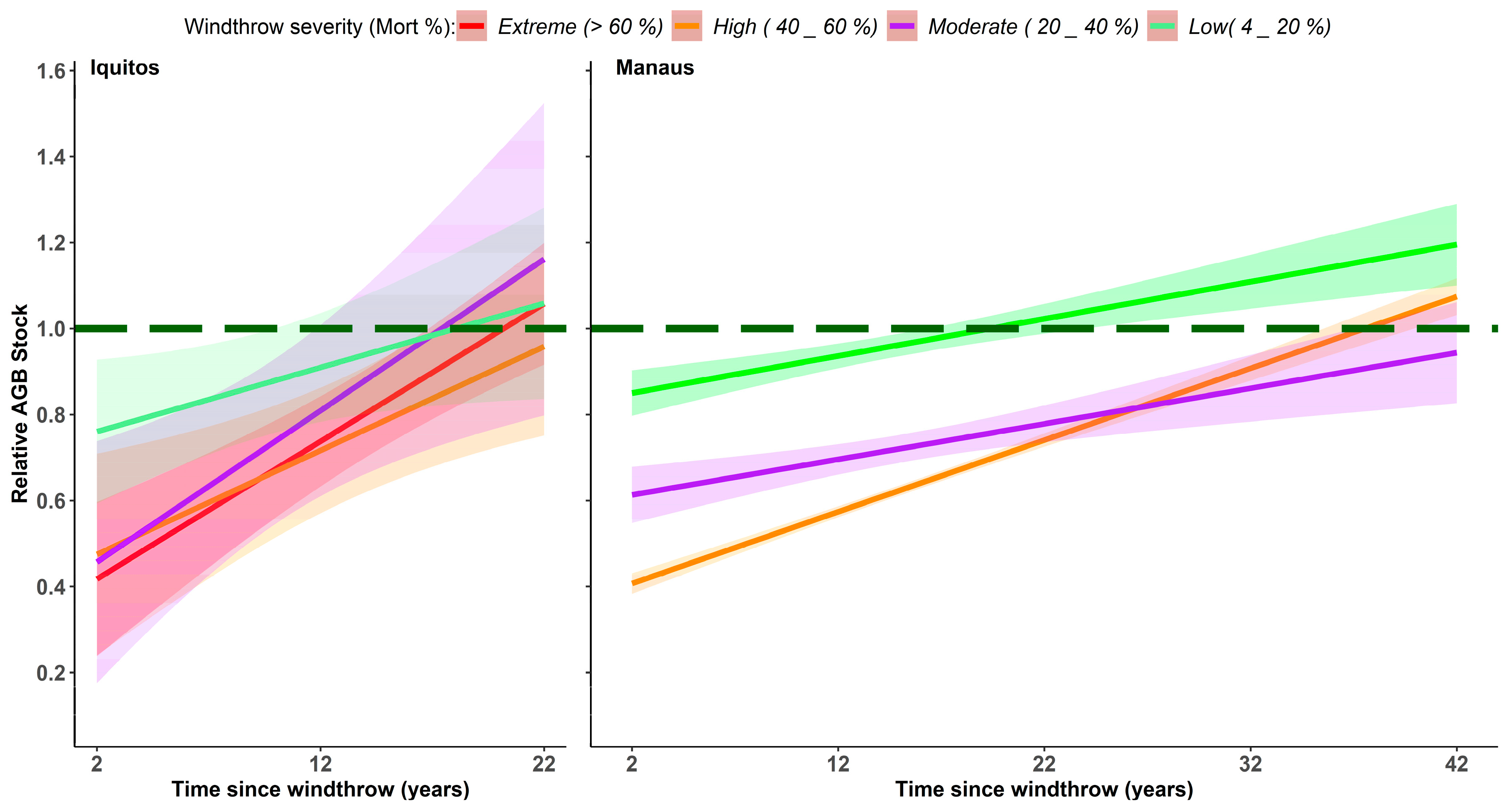
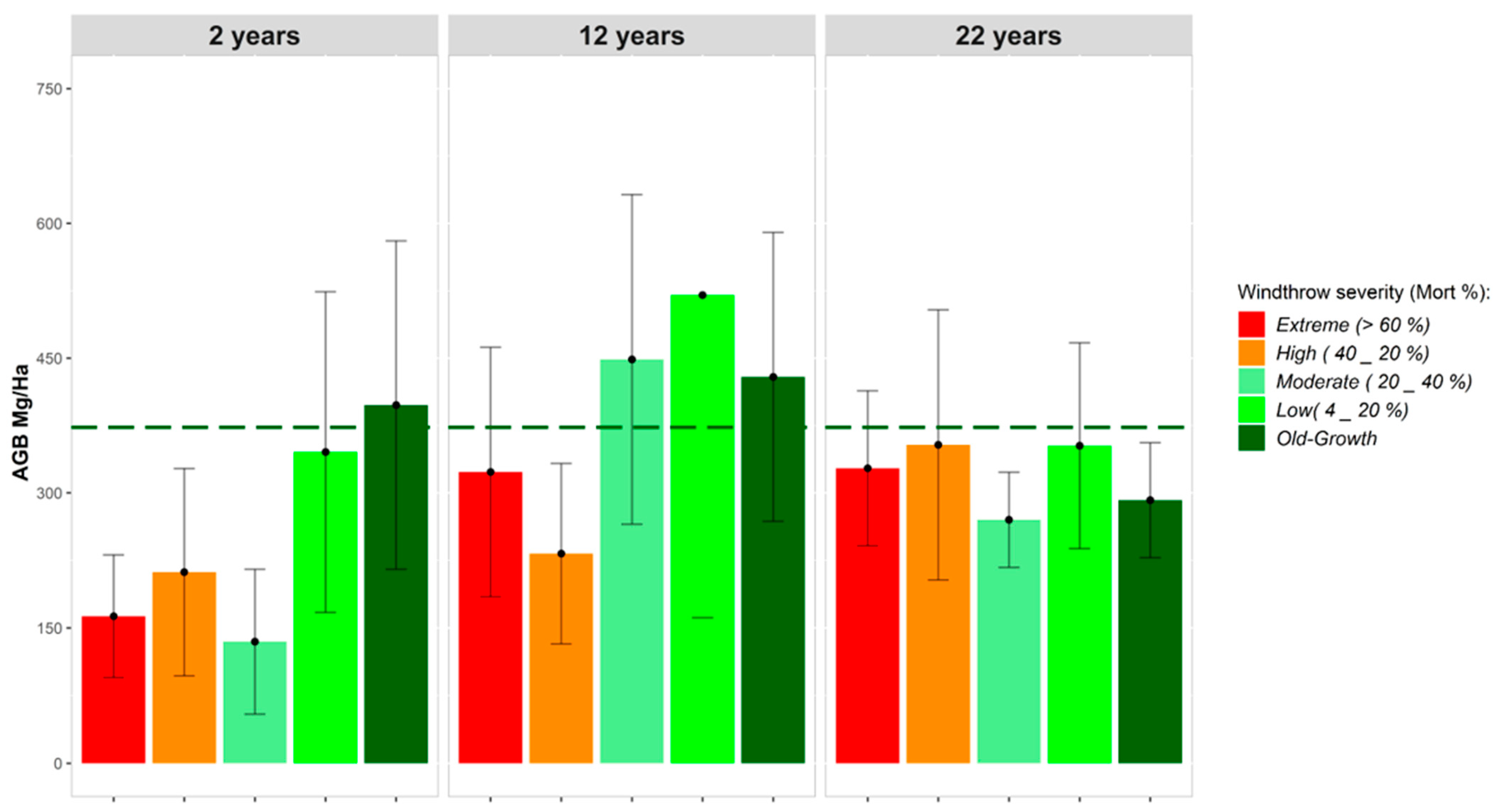
3.3.2. Aboveground Tree Biomass
4. Discussion
4.1. Patterns of Recovery after Windthrow
4.2. Time Span of Biomass Recovery
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years after Windthrow | Severity | Number of Families | Number of Genus | Number of Species |
|---|---|---|---|---|
| 2 | Extreme | 29 | 52 | 75 |
| High | 20 | 42 | 68 | |
| Moderate | 23 | 45 | 76 | |
| Low | 39 | 80 | 174 | |
| Old-growth | 46 | 112 | 245 | |
| 12 | Extreme | 34 | 86 | 125 |
| High | 26 | 53 | 79 | |
| Moderate | 41 | 95 | 161 | |
| Low | 46 | 100 | 185 | |
| Old-growth | 52 | 159 | 342 | |
| 22 | Extreme | 39 | 112 | 222 |
| High | 39 | 84 | 155 | |
| Moderate | 31 | 54 | 82 | |
| Low | 34 | 89 | 170 | |
| Old-growth | 47 | 120 | 257 |
Appendix B

Appendix C

References
- Nelson, B.; Amaral, I. Destructive Wind Effects Detected in TM Image in the Amazon Basin. In Proceedings of the International Society for Photogrammetry and Remote Sensing, Rio de Janeiro, Brazil, 26–30 September 1994; Brazil’s National Institute for Space Research: Rio de Janeiro, Brazil, 1994; pp. 339–343. [Google Scholar]
- Garstang, M.; White, S.; Shugart, H.H.; Halverson, J. Convective Cloud Downdrafts as the Cause of Large Blowdowns in the Amazon Rainforest. Meteorl. Atmos. Phys. 1998, 67, 199–212. [Google Scholar] [CrossRef]
- Negrón-Juárez, R.I.; Chambers, J.Q.; Guimaraes, G.; Zeng, H.; Raupp, C.F.M.; Marra, D.M.; Ribeiro, G.H.P.M.; Saatchi, S.S.; Nelson, B.W.; Higuchi, N. Widespread Amazon Forest Tree Mortality from a Single Cross-Basin Squall Line Event: Wind-Driven Tree Mortality in Amazonia. Geophys. Res. Lett. 2010, 37. [Google Scholar] [CrossRef]
- Mitchell, S.J. Wind as a Natural Disturbance Agent in Forests: A Synthesis. Forestry 2013, 86, 147–157. [Google Scholar] [CrossRef]
- Magnabosco Marra, D.; Higuchi, N.; Trumbore, S.E.; Ribeiro, G.H.P.M.; dos Santos, J.; Carneiro, V.M.C.; Lima, A.J.N.; Chambers, J.Q.; Negrón-Juárez, R.I.; Holzwarth, F.; et al. Predicting Biomass of Hyperdiverse and Structurally Complex Central Amazonian Forests–a Virtual Approach Using Extensive Field Data. Biogeosciences 2016, 13, 1553–1570. [Google Scholar] [CrossRef]
- Rifai, S.W.; Urquiza Muñoz, J.D.; Negrón-Juárez, R.I.; Ramírez Arévalo, F.R.; Tello-Espinoza, R.; Vanderwel, M.C.; Lichstein, J.W.; Chambers, J.Q.; Bohlman, S.A. Landscape-Scale Consequences of Differential Tree Mortality from Catastrophic Wind Disturbance in the Amazon. Ecol. Appl. 2016, 26, 2225–2237. [Google Scholar] [CrossRef]
- Negrón-Juárez, R.; Jenkins, H.; Raupp, C.; Riley, W.; Kueppers, L.; Magnabosco Marra, D.; Ribeiro, G.; Monteiro, M.; Candido, L.; Chambers, J.; et al. Windthrow Variability in Central Amazonia. Atmosphere 2017, 8, 28. [Google Scholar] [CrossRef]
- Magnabosco Marra, D.; Trumbore, S.E.; Higuchi, N.; Ribeiro, G.H.P.M.; Negrón-Juárez, R.I.; Holzwarth, F.; Rifai, S.W.; dos Santos, J.; Lima, A.J.N.; Kinupp, V.F.; et al. Windthrows Control Biomass Patterns and Functional Composition of Amazon Forests. Glob. Chang. Biol. 2018, 24, 5867–5881. [Google Scholar] [CrossRef]
- Silvério, D.V.; Brando, P.M.; Bustamante, M.M.C.; Putz, F.E.; Marra, D.M.; Levick, S.R.; Trumbore, S.E. Fire, Fragmentation, and Windstorms: A Recipe for Tropical Forest Degradation. J. Ecol. 2019, 107, 656–667. [Google Scholar] [CrossRef]
- Esquivel-Muelbert, A.; Phillips, O.L.; Brienen, R.J.W.; Fauset, S.; Sullivan, M.J.P.; Baker, T.R.; Chao, K.-J.; Feldpausch, T.R.; Gloor, E.; Higuchi, N.; et al. Tree Mode of Death and Mortality Risk Factors across Amazon Forests. Nat. Commun. 2020, 11, 5515. [Google Scholar] [CrossRef]
- Espírito-Santo, F.D.B.; Gloor, M.; Keller, M.; Malhi, Y.; Saatchi, S.; Nelson, B.; Junior, R.C.O.; Pereira, C.; Lloyd, J.; Frolking, S.; et al. Size and Frequency of Natural Forest Disturbances and the Amazon Forest Carbon Balance. Nat. Commun. 2014, 5, 3434. [Google Scholar] [CrossRef]
- Negrón-Juárez, R.I.; Holm, J.A.; Marra, D.M.; Rifai, S.W.; Riley, W.J.; Chambers, J.Q.; Koven, C.D.; Knox, R.G.; McGroddy, M.E.; Di Vittorio, A.V.; et al. Vulnerability of Amazon Forests to Storm-Driven Tree Mortality. Environ. Res. Lett. 2018, 13, 054021. [Google Scholar] [CrossRef]
- Chesson, P. Overview: Nonequilibrium community theories: Chance, variability, history, and coexistence. In Community Ecology; Diamond, J., Case, T.J., Eds.; Harper and Row: New York, NY, USA, 1986; pp. 229–239. [Google Scholar]
- Ennos, A.R. Wind as a Ecological Factor. Trends Ecol. Evol. 1997, 12, 108–111. [Google Scholar] [CrossRef]
- Marra, D.M.; Chambers, J.Q.; Higuchi, N.; Trumbore, S.E.; Ribeiro, G.H.P.M.; dos Santos, J.; Negrón-Juárez, R.I.; Reu, B.; Wirth, C. Large-Scale Wind Disturbances Promote Tree Diversity in a Central Amazon Forest. PLoS ONE 2014, 9, e103711. [Google Scholar] [CrossRef]
- Ulanova, N.G. The Effects of Windthrow on Forests at Different Spatial Scales: A Review. For. Ecol. Manag. 2000, 135, 155–167. [Google Scholar] [CrossRef]
- Denslow, J.S. Tropical Rainforest Gaps and Tree Species Diversity. Annu. Rev. Ecol. Syst. 1987, 18, 431–451. [Google Scholar] [CrossRef]
- Baker, T.R.; Vela Díaz, D.M.; Chama Moscoso, V.; Navarro, G.; Monteagudo, A.; Pinto, R.; Cangani, K.; Fyllas, N.M.; Lopez Gonzalez, G.; Laurance, W.F.; et al. Consistent, Small Effects of Treefall Disturbances on the Composition and Diversity of Four Amazonian Forests. J. Ecol. 2016, 104, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R.; Schelhaas, M.-J.; Lexer, M.J. Unraveling the Drivers of Intensifying Forest Disturbance Regimes in Europe: Drivers of Forest Disturbance Intensification. Glob. Chang. Biol. 2011, 17, 2842–2852. [Google Scholar] [CrossRef]
- Quine, C.P.; Gardiner, B.A. Understanding How the Interaction of Wind and Trees Results in Windthrow, Stem Breakage, and Canopy Gap Formation. In Plant Disturbance Ecology; Elsevier: Amsterdam, The Netherlands, 2007; pp. 103–155. ISBN 978-0-12-088778-1. [Google Scholar]
- Espinoza Villar, J.C.; Ronchail, J.; Guyot, J.L.; Cochonneau, G.; Naziano, F.; Lavado, W.; De Oliveira, E.; Pombosa, R.; Vauchel, P. Spatio-Temporal Rainfall Variability in the Amazon Basin Countries (Brazil, Peru, Bolivia, Colombia, and Ecuador). Int. J. Climatol. 2009, 29, 1574–1594. [Google Scholar] [CrossRef]
- Marengo, J.A. Interdecadal Variability and Trends of Rainfall across the Amazon Basin. Theor. Appl. Climatol. 2004, 78, 79–96. [Google Scholar] [CrossRef]
- Quesada, C.A.; Lloyd, J.; Anderson, L.O.; Fyllas, N.M.; Schwarz, M.; Czimczik, C.I. Soils of Amazonia with Particular Reference to the Rainfor Sites. Biogeosciences 2011, 8, 1415–1440. [Google Scholar] [CrossRef]
- Gloor, M.; Brienen, R.J.W.; Galbraith, D.; Feldpausch, T.R.; Schöngart, J.; Guyot, J.-L.; Espinoza, J.C.; Lloyd, J.; Phillips, O.L. Intensification of the Amazon Hydrological Cycle over the Last Two Decades. Geophys. Res. Lett. 2013, 40, 1729–1733. [Google Scholar] [CrossRef]
- Nemani, R.R. Climate-Driven Increases in Global Terrestrial Net Primary Production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Malhi, Y.; Davidson, E.A. Biogeochemistry and Ecology of terrestrial Ecosystems of Amazonia. In Amazonia and Global Change; American Geophysical Union: Washington, DC, USA, 2009; ISBN 978-0-87590-476-4. [Google Scholar]
- de Mello Ribeiro, G.H.P.; Suwa, R.; Marra, D.M.; Lima, A.J.N.; Kajimoto, T.; Ishizuka, M.; Higuchi, N. Allometry for Juvenile Trees in an Amazonian Forest after Wind Disturbance. JARQ 2014, 48, 213–219. [Google Scholar] [CrossRef][Green Version]
- Andrewsa, T.; Dietze, M.; Booth, R. Climate or Disturbance: Temperate Forest Structural Change and Carbon Sink Potential. bioRxiv 2018, 478693. [Google Scholar] [CrossRef]
- Sombroek, W. Spatial and Temporal Patterns of Amazon Rainfall: Consequences for the Planning of Agricultural Occupation and the Protection of Primary Forests. AMBIO J. Hum. Environ. 2001, 30, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.B.; Gillespie, A.R. Remote Sensing of Landscapes with Spectral Images: A Physical Modeling Approach; Cambridge University Press: Cambridge, UK, 2006; ISBN 978-0-511-61719-5. [Google Scholar]
- Saldarriaga, J.G.; West, D.C.; Tharp, M.L.; Uhl, C. Long-Term Chronosequence of Forest Succession in the Upper Rio Negro of Colombia and Venezuela. J. Ecol. 1988, 76, 938. [Google Scholar] [CrossRef]
- Masek, J.G.; Vermote, E.F.; Saleous, N.E.; Wolfe, R.; Hall, F.G.; Huemmrich, K.F.; Gao, F.; Kutler, J.; Lim, T.-K. A Landsat Surface Reflectance Dataset for North America, 1990–2000. IEEE Geosci. Remote Sens. Lett. 2006, 3, 68–72. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-Scale Geospatial Analysis for Everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Adams, J. Classification of Multispectral Images Based on Fractions of Endmembers: Application to Land-Cover Change in the Brazilian Amazon. Remote Sens. Environ. 1995, 52, 137–154. [Google Scholar] [CrossRef]
- Chambers, J.Q.; Negrón-Juárez, R.I.; Hurtt, G.C.; Marra, D.M.; Higuchi, N. Lack of Intermediate-Scale Disturbance Data Prevents Robust Extrapolation of Plot-Level Tree Mortality Rates for Old-Growth Tropical Forests: Clustered Gaps Tropical Tree Mortality Rates. Ecol. Lett. 2009, 12, E22–E25. [Google Scholar] [CrossRef]
- Balderas Torres, A.; Lovett, J.C. Using Basal Area to Estimate Aboveground Carbon Stocks in Forests: La Primavera Biosphere’s Reserve, Mexico. Forestry 2013, 86, 267–281. [Google Scholar] [CrossRef]
- Chave, J.; Réjou-Méchain, M.; Búrquez, A.; Chidumayo, E.; Colgan, M.S.; Delitti, W.B.C.; Duque, A.; Eid, T.; Fearnside, P.M.; Goodman, R.C.; et al. Improved Allometric Models to Estimate the Aboveground Biomass of Tropical Trees. Glob. Chang. Biol. 2014, 20, 3177–3190. [Google Scholar] [CrossRef] [PubMed]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a Worldwide Wood Economics Spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- De Oliveira, M.M.; Higuchi, N.; Celes, C.H.; Higuchi, F.G. Tamanho e Formas de Parcelas Para Inventários Florestais de Espécies Arbóreas na Amazônia Central. Ciênc. Florest. 2014, 24, 645–653. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Feldpausch, T.R.; Banin, L.; Phillips, O.L.; Baker, T.R.; Lewis, S.L.; Quesada, C.A.; Affum-Baffoe, K.; Arets, E.J.M.M.; Berry, N.J.; Bird, M.; et al. Height-Diameter Allometry of Tropical Forest Trees. Biogeosciences 2011, 8, 1081–1106. [Google Scholar] [CrossRef]
- ter Steege, H.; Pitman, N.C.A.; Phillips, O.L.; Chave, J.; Sabatier, D.; Duque, A.; Molino, J.-F.; Prévost, M.-F.; Spichiger, R.; Castellanos, H.; et al. Continental-Scale Patterns of Canopy Tree Composition and Function across Amazonia. Nature 2006, 443, 444–447. [Google Scholar] [CrossRef]
- Chao, K.-J.; Phillips, O.L.; Gloor, E.; Monteagudo, A.; Torres-Lezama, A.; Martínez, R.V. Growth and Wood Density Predict Tree Mortality in Amazon Forests. J. Ecol. 2008, 96, 281–292. [Google Scholar] [CrossRef]
- Aigbe, H.I.; Omokhua, G.E. Modeling Diameter Distribution of the Tropical Rainforest in Oban Forest Reserve. JEE 2014, 5, 130. [Google Scholar] [CrossRef]
- Muller-Landau, H.C.; Condit, R.S.; Harms, K.E.; Marks, C.O.; Thomas, S.C.; Bunyavejchewin, S.; Chuyong, G.; Co, L.; Davies, S.; Foster, R.; et al. Comparing Tropical Forest Tree Size Distributions with the Predictions of Metabolic Ecology and Equilibrium Models. Ecol. Lett. 2006, 9, 589–602. [Google Scholar] [CrossRef]
- Niklas, K.J.; Midgley, J.J.; Rand, R.H. Tree Size Frequency Distributions, Plant Density, Age and Community Disturbance: Tree Size Distributions. Ecol. Lett. 2003, 6, 405–411. [Google Scholar] [CrossRef]
- Chambers, J.Q.; Negron-Juarez, R.I.; Marra, D.M.; Di Vittorio, A.; Tews, J.; Roberts, D.; Ribeiro, G.H.P.M.; Trumbore, S.E.; Higuchi, N. The Steady-State Mosaic of Disturbance and Succession across an Old-Growth Central Amazon Forest Landscape. Proc. Natl. Acad. Sci. USA 2013, 110, 3949–3954. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R.V.; Phillips, O.L. Allpahuayo: Floristics, Structure, and Dynamics of a High-Diversity Forest in Amazonian Peru. Ann. Mo. Bot. Gard. 2000, 87, 499. [Google Scholar] [CrossRef]
- Lewis, S.L.; Phillips, O.L.; Baker, T.R.; Lloyd, J.; Malhi, Y.; Almeida, S.; Higuchi, N.; Laurance, W.F.; Neill, D.A.; Silva, J.N.M.; et al. Concerted Changes in Tropical Forest Structure and Dynamics: Evidence from 50 South American Long-Term Plots. Phil. Trans. R. Soc. Lond. B 2004, 359, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Chazdon, R.L.; Letcher, S.G.; van Breugel, M.; Martínez-Ramos, M.; Bongers, F.; Finegan, B. Rates of Change in Tree Communities of Secondary Neotropical Forests Following Major Disturbances. Phil. Trans. R. Soc. B 2007, 362, 273–289. [Google Scholar] [CrossRef]
- Sheil, D.; Burslem, D.F.R.P. Disturbing Hypotheses in Tropical Forests. Trends Ecol. Evol. 2003, 18, 18–26. [Google Scholar] [CrossRef]
- Poorter, L.; Bongers, F.; Aide, T.M.; Almeyda Zambrano, A.M.; Balvanera, P.; Becknell, J.M.; Boukili, V.; Brancalion, P.H.S.; Broadbent, E.N.; Chazdon, R.L.; et al. Biomass Resilience of Neotropical Secondary Forests. Nature 2016, 530, 211–214. [Google Scholar] [CrossRef]
- Chazdon, R.L. Tropical Forest Recovery: Legacies of Human Impact and Natural Disturbances. Perspect. Plant Ecol. Evol. Syst. 2003, 6, 51–71. [Google Scholar] [CrossRef]
- D’Oliveira, M.V.N.; Alvarado, E.C.; Santos, J.C.; Carvalho, J.A. Forest Natural Regeneration and Biomass Production after Slash and Burn in a Seasonally Dry Forest in the Southern Brazilian Amazon. For. Ecol. Manag. 2011, 261, 1490–1498. [Google Scholar] [CrossRef]
- Elias, F.; Ferreira, J.; Lennox, G.D.; Berenguer, E.; Ferreira, S.; Schwartz, G.; de Oliveira Melo, L.; Reis Júnior, D.N.; Nascimento, R.O.; Ferreira, F.N.; et al. Assessing the Growth and Climate Sensitivity of Secondary Forests in Highly Deforested Amazonian Landscapes. Ecology 2020, 101. [Google Scholar] [CrossRef]
- Veldkamp, E.; Schmidt, M.; Powers, J.S.; Corre, M.D. Deforestation and Reforestation Impacts on Soils in the Tropics. Nature Rev. Earth Environ. 2020, 1, 590–605. [Google Scholar] [CrossRef]
- Scatena, F.N.; Moya, S.; Estrada, C.; Chinea, J.D. The First Five Years in the Reorganization of Aboveground Biomass and Nutrient Use Following Hurricane Hugo in the Bisley Experimental Watersheds, Luquillo Experimental Forest, Puerto Rico. Biotropica 1996, 28, 424. [Google Scholar] [CrossRef]
- Everham, E.M.; Brokaw, N.V.L. Forest Damage and Recovery from Catastrophic Wind. Bot. Rev. 1996, 62, 113–185. [Google Scholar] [CrossRef]
- Boose, E.R.; Serrano, M.I.; Foster, D.R. Landscape and Regional Impacts of Hurricanes in Puerto Rico. Ecol. Monogr. 2004, 74, 335–352. [Google Scholar] [CrossRef]
- Silva, J.N.M.; de Carvalho, J.O.P.; Lopes, J.d.C.A.; de Almeida, B.F.; Costa, D.H.M.; de Oliveira, L.C.; Vanclay, J.K.; Skovsgaard, J.P. Growth and Yield of a Tropical Rain Forest in the Brazilian Amazon 13 Years after Logging. For. Ecol. Manag. 1995, 71, 267–274. [Google Scholar] [CrossRef]
- Miller, S.D.; Goulden, M.L.; Hutyra, L.R.; Keller, M.; Saleska, S.R.; Wofsy, S.C.; Figueira, A.M.S.; da Rocha, H.R.; de Camargo, P.B. Reduced Impact Logging Minimally Alters Tropical Rainforest Carbon and Energy Exchange. Proc. Natl. Acad. Sci. USA 2011, 108, 19431–19435. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, E.M.; Fearnside, P.M.; Nelson, B.W.; Barbosa, R.I.; Keizer, E.W.H. Estimates of Forest Biomass in the Brazilian Amazon: New Allometric Equations and Adjustments to Biomass from Wood-Volume Inventories. For. Ecol. Manag. 2008, 256, 1853–1867. [Google Scholar] [CrossRef]
- Houghton, R.A.; Lawrence, K.T.; Hackler, J.L.; Brown, S. The Spatial Distribution of Forest Biomass in the Brazilian Amazon: A Comparison of Estimates. Glob. Chang. Biol. 2001, 7, 731–746. [Google Scholar] [CrossRef]
- Lima, A.J.N.; Suwa, R.; de Mello Ribeiro, G.H.P.; Kajimoto, T.; dos Santos, J.; da Silva, R.P.; de Souza, C.A.S.; de Barros, P.C.; Noguchi, H.; Ishizuka, M.; et al. Allometric Models for Estimating Above- and below-Ground Biomass in Amazonian Forests at São Gabriel Da Cachoeira in the Upper Rio Negro, Brazil. For. Ecol. Manag. 2012, 277, 163–172. [Google Scholar] [CrossRef]
- Espírito, F.D.B.; Keller, M.; Braswell, B.; Nelson, B.W.; Frolking, S.; Vicente, G. Storm Intensity and Old-growth Forest Disturbances in the Amazon Region. Geophys. Res. Lett. 2010, 37, 6. [Google Scholar] [CrossRef]
- Rozendaal, D.M.A.; Phillips, O.L.; Lewis, S.L.; Affum-Baffoe, K.; Alvarez-Davila, E.; Andrade, A.; Aragão, L.E.O.C.; Araujo-Murakami, A.; Baker, T.R.; Bánki, O.; et al. Competition Influences Tree Growth, but Not Mortality, across Environmental Gradients in Amazonia and Tropical Africa. Ecology 2020, 101, e03052. [Google Scholar] [CrossRef]
- Stephenson, N.L.; van Mantgem, P.J.; Bunn, A.G.; Bruner, H.; Harmon, M.E.; O’Connell, K.B.; Urban, D.L.; Franklin, J.F. Causes and Implications of the Correlation between Forest Productivity and Tree Mortality Rates. Ecol. Monogr. 2011, 81, 527–555. [Google Scholar] [CrossRef]
- Zelnik, Y.R.; Arnoldi, J.-F.; Loreau, M. The Impact of Spatial and Temporal Dimensions of Disturbances on Ecosystem Stability. Front. Ecol. Evol. 2018, 6, 224. [Google Scholar] [CrossRef] [PubMed]




| Model | Equation | Reference |
|---|---|---|
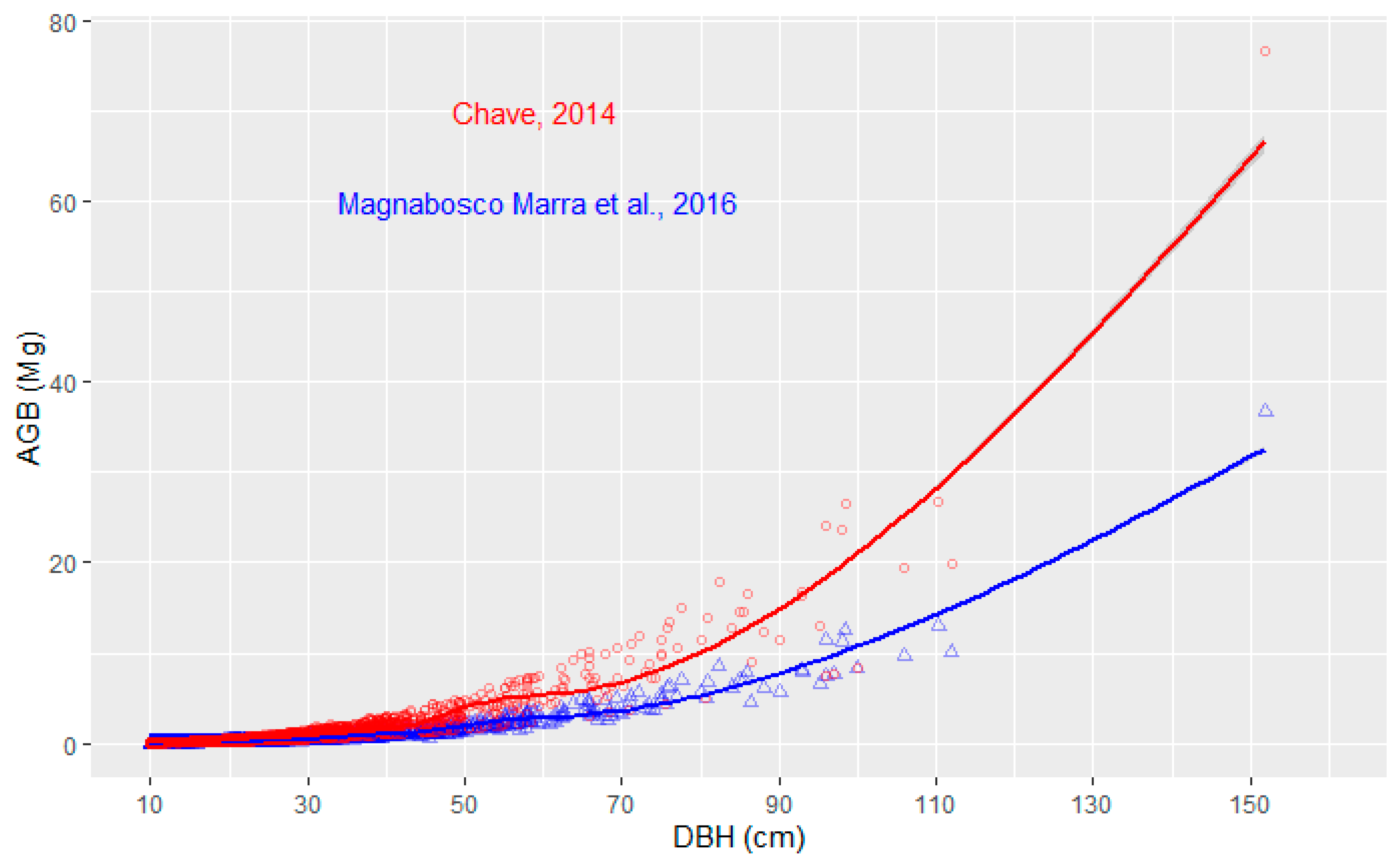
| Model 1 | AGBest = exp [−1.803 − 0.97 E + 0.976 ln (WD) + 2.673 ln (D) − 0.229 [ln(D)]2] | Equation (7) in [37] |
| Model 2 | AGBest = 0.230 D2.406·(WD)0.88 | Equation (33) in [5] |
| Model 3 | AGBest = 0.183 D2.328 | Equation (13) in [5] |
| Mortality (%) | Disturbance Severity | Years Since Windthrow | Total Number of Subplots by Severity | ||
|---|---|---|---|---|---|
| 2 Years | 12 Years | 22 Years | |||
| ≤4 | Old-growth | 39 (1.17 ha) | 46 (1.38 ha) | 35 (1.03 ha) | 120 |
| >4 & ≤20 | Low | 27 (0.81 ha) | 20 (0.6 ha) | 17 (0.51 ha) | 64 |
| >20 & ≤40 | Moderate | 14 (0.42 ha) | 14 (0.42 ha) | 7 (0.21 ha) | 35 |
| >40 & ≤60 | High | 10 (0.30 ha) | 7 (0.21 ha) | 15 (0.45 ha) | 32 |
| >60 (max:94) | Extreme | 13 (0.39 ha) | 13 (0.39 ha) | 26 (0.78 ha) | 52 |
| Total subplots by years | 103 | 100 | 100 | 303 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urquiza Muñoz, J.D.; Magnabosco Marra, D.; Negrón-Juarez, R.I.; Tello-Espinoza, R.; Alegría-Muñoz, W.; Pacheco-Gómez, T.; Rifai, S.W.; Chambers, J.Q.; Jenkins, H.S.; Brenning, A.; et al. Recovery of Forest Structure Following Large-Scale Windthrows in the Northwestern Amazon. Forests 2021, 12, 667. https://doi.org/10.3390/f12060667
Urquiza Muñoz JD, Magnabosco Marra D, Negrón-Juarez RI, Tello-Espinoza R, Alegría-Muñoz W, Pacheco-Gómez T, Rifai SW, Chambers JQ, Jenkins HS, Brenning A, et al. Recovery of Forest Structure Following Large-Scale Windthrows in the Northwestern Amazon. Forests. 2021; 12(6):667. https://doi.org/10.3390/f12060667
Chicago/Turabian StyleUrquiza Muñoz, J. David, Daniel Magnabosco Marra, Robinson I. Negrón-Juarez, Rodil Tello-Espinoza, Waldemar Alegría-Muñoz, Tedi Pacheco-Gómez, Sami W. Rifai, Jeffrey Q. Chambers, Hillary S. Jenkins, Alexander Brenning, and et al. 2021. "Recovery of Forest Structure Following Large-Scale Windthrows in the Northwestern Amazon" Forests 12, no. 6: 667. https://doi.org/10.3390/f12060667
APA StyleUrquiza Muñoz, J. D., Magnabosco Marra, D., Negrón-Juarez, R. I., Tello-Espinoza, R., Alegría-Muñoz, W., Pacheco-Gómez, T., Rifai, S. W., Chambers, J. Q., Jenkins, H. S., Brenning, A., & Trumbore, S. E. (2021). Recovery of Forest Structure Following Large-Scale Windthrows in the Northwestern Amazon. Forests, 12(6), 667. https://doi.org/10.3390/f12060667