
Climatic Aridity Shapes Post-Fire Interactions between Ceanothus spp. and Douglas-Fir (Pseudotsuga menziesii) across the Klamath Mountains

 ,
, 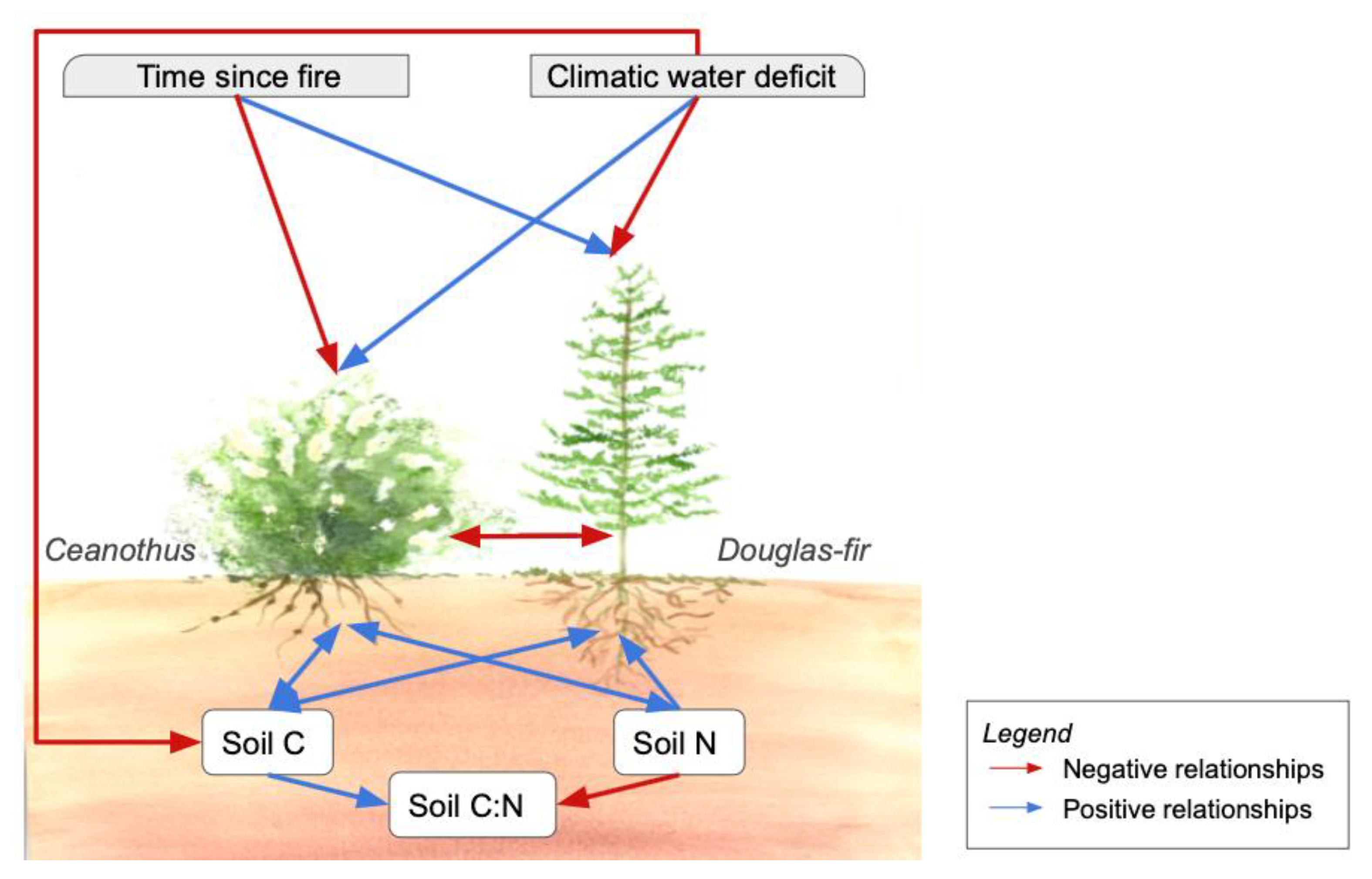{kind=link}
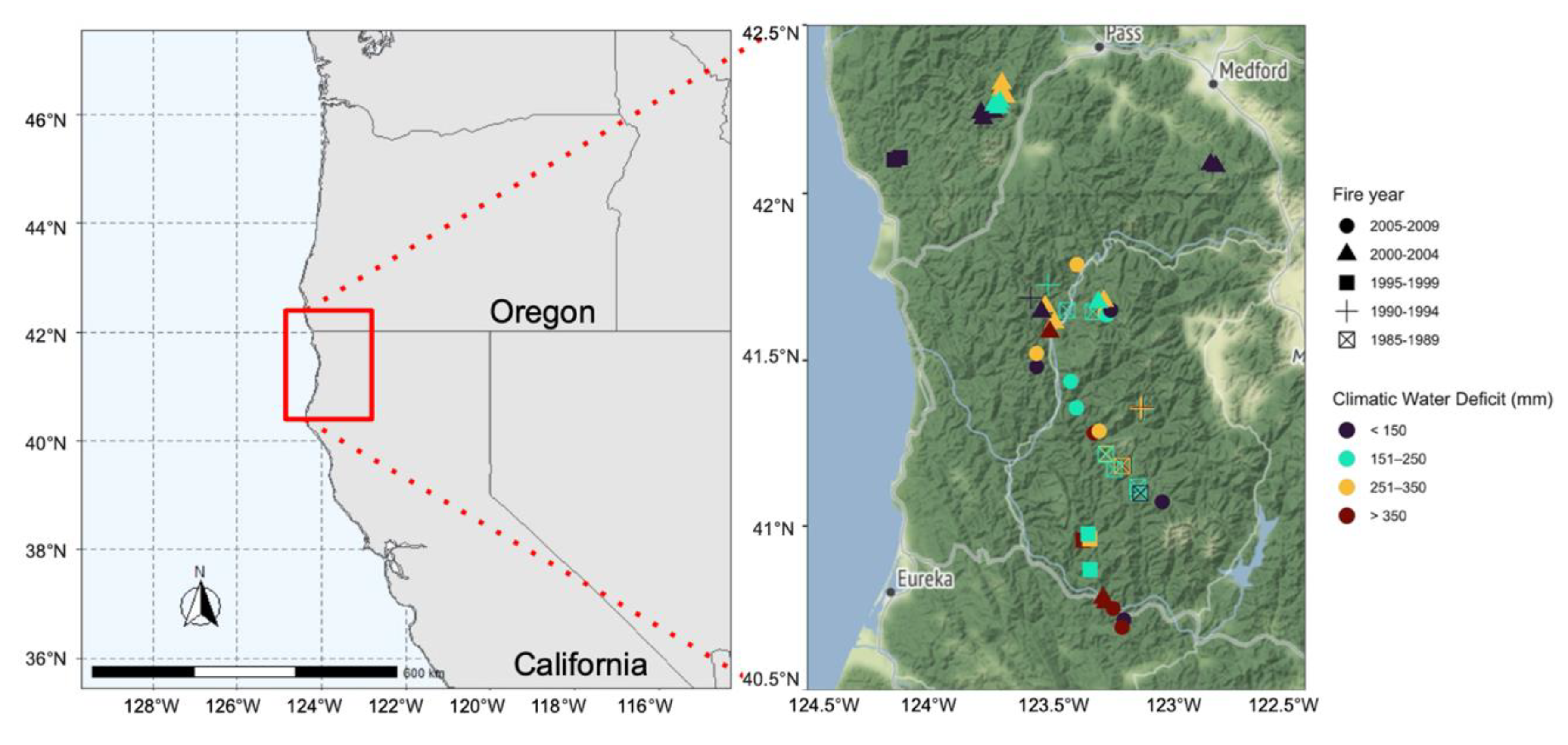{kind=link}
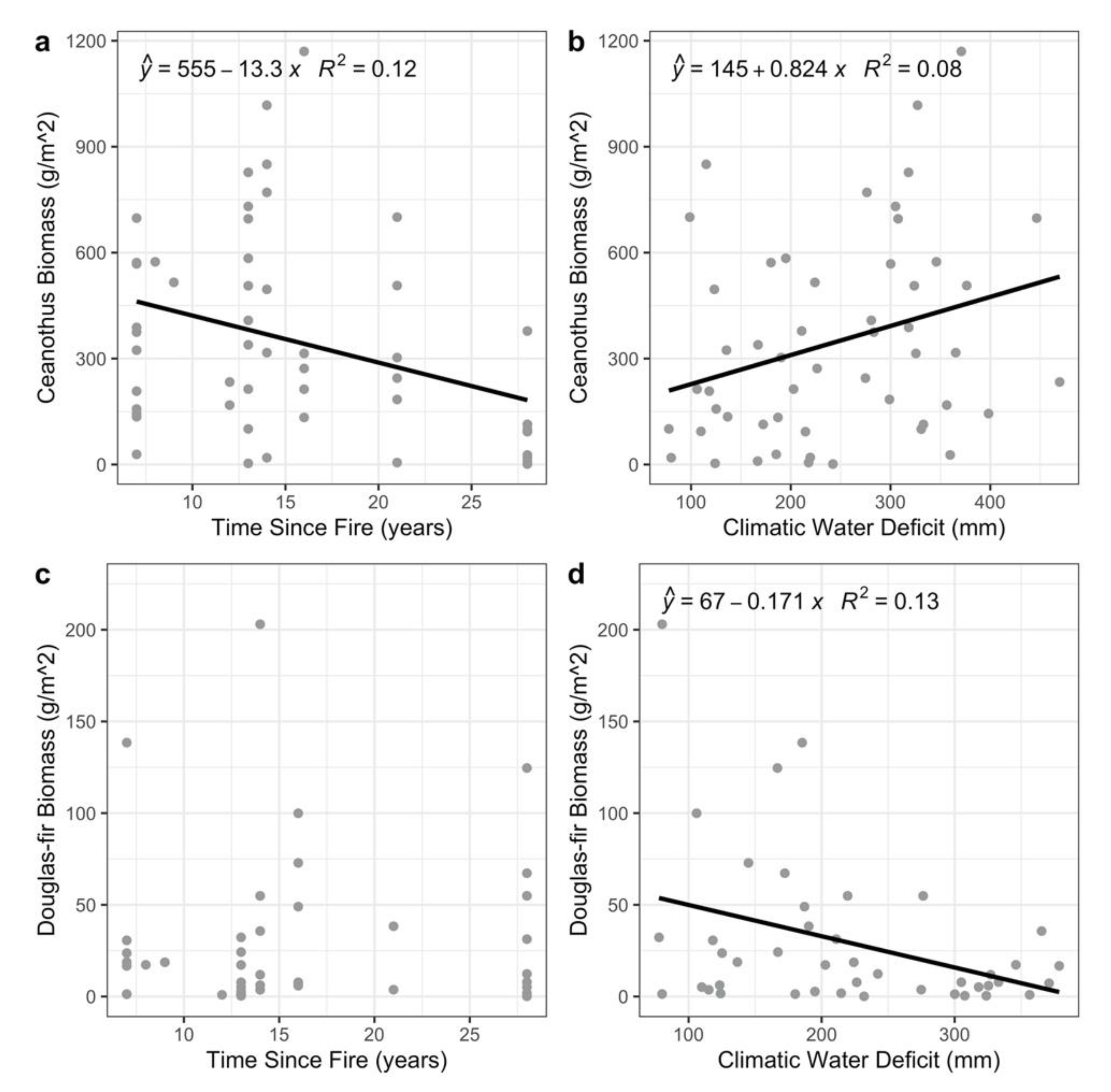{kind=link}
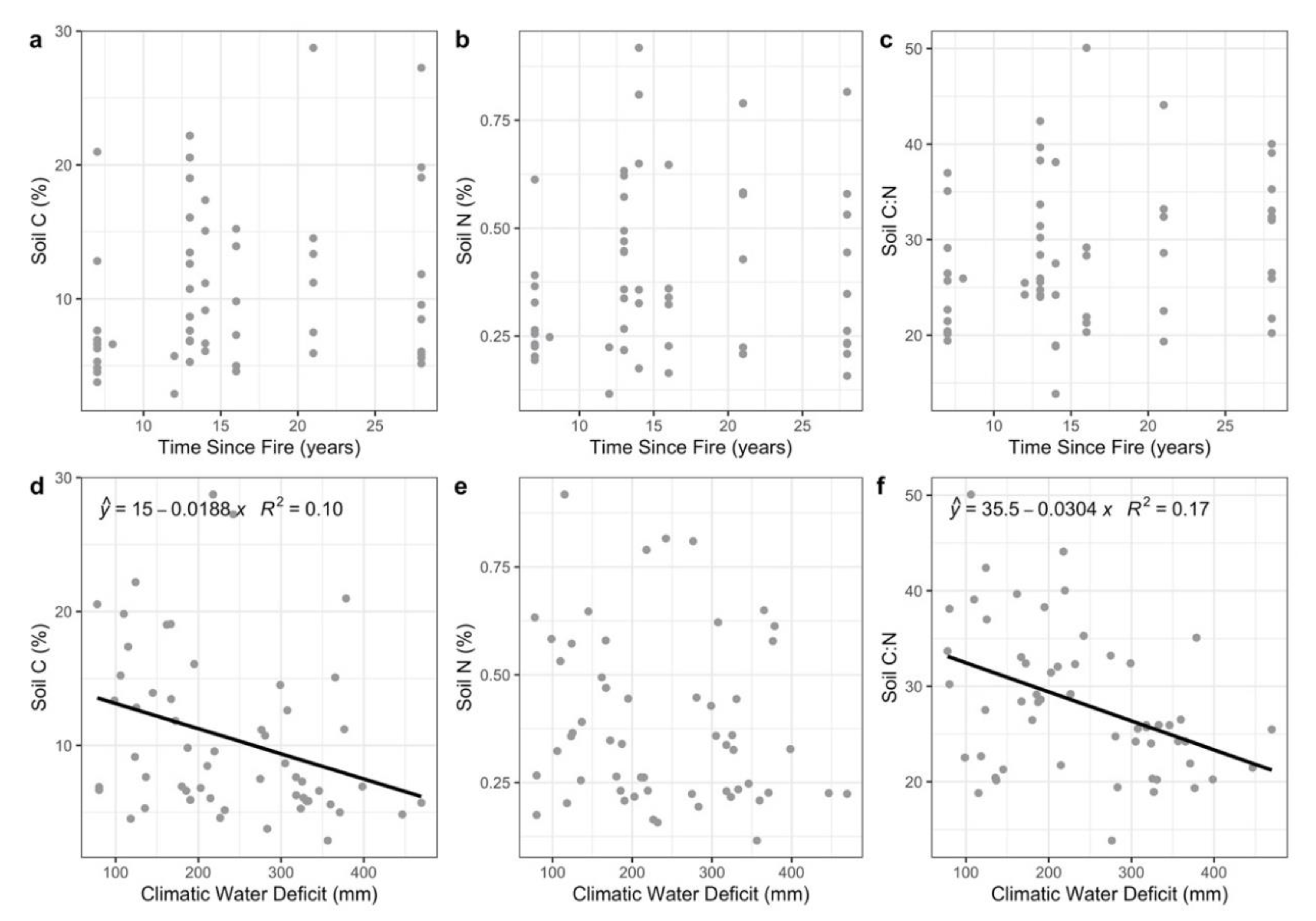{kind=link}
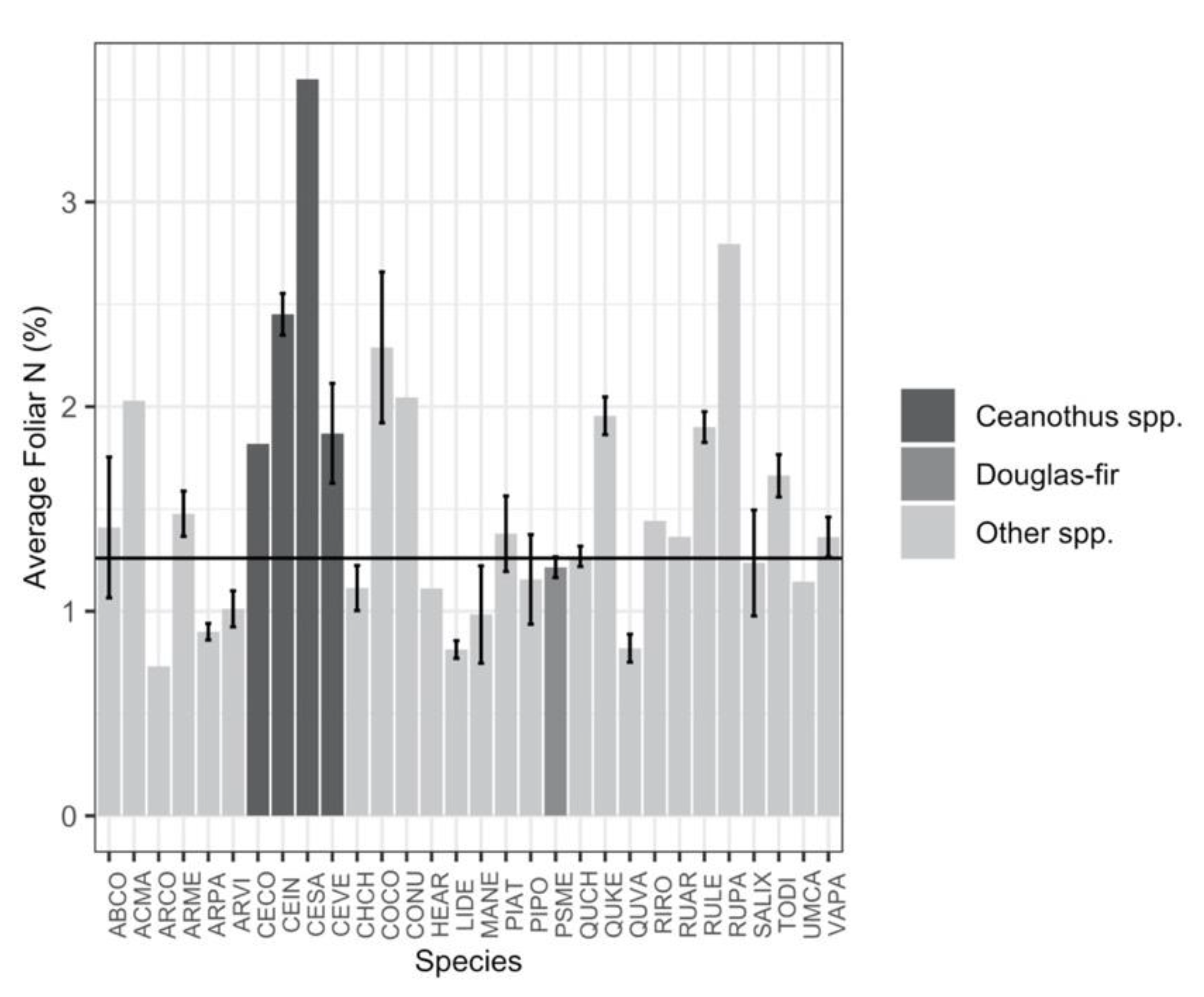{kind=link}
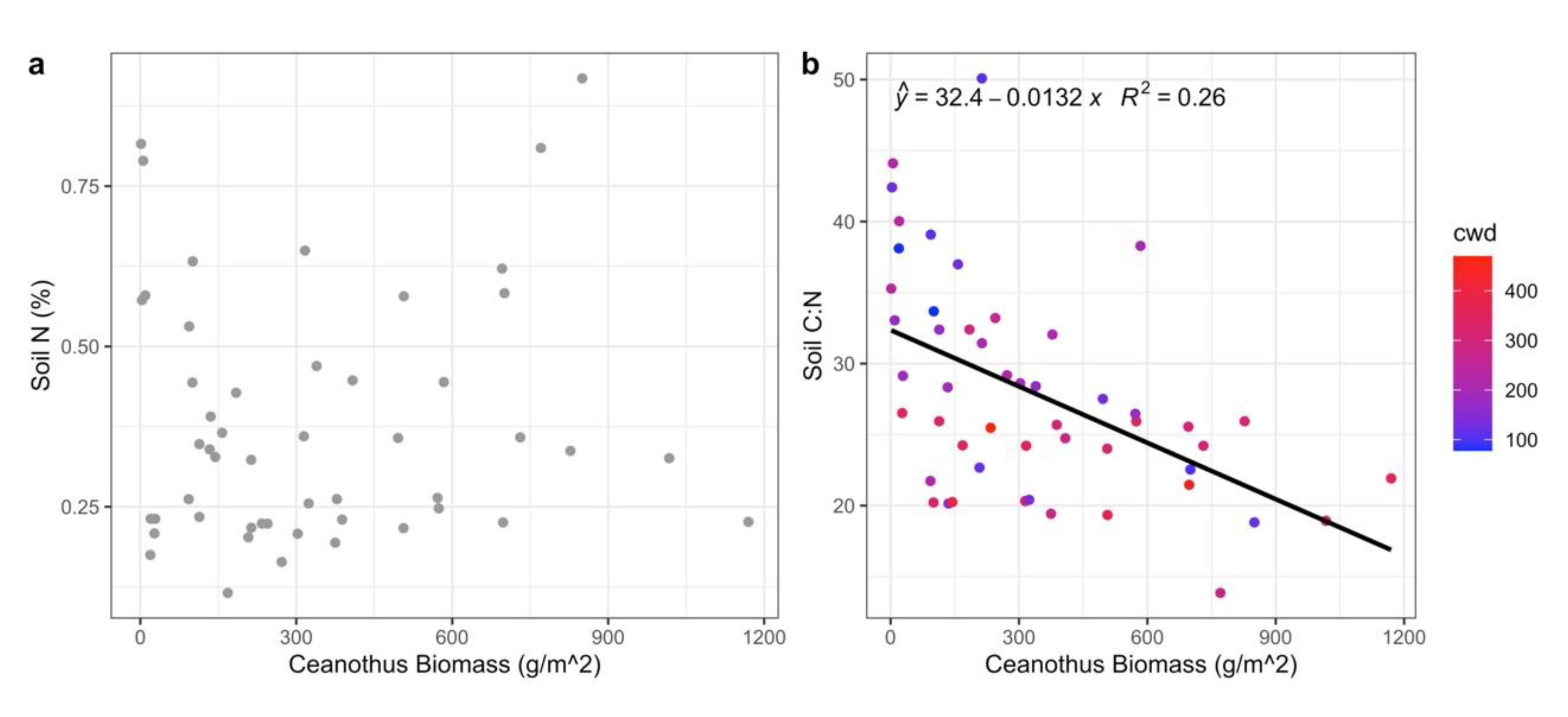{kind=link}
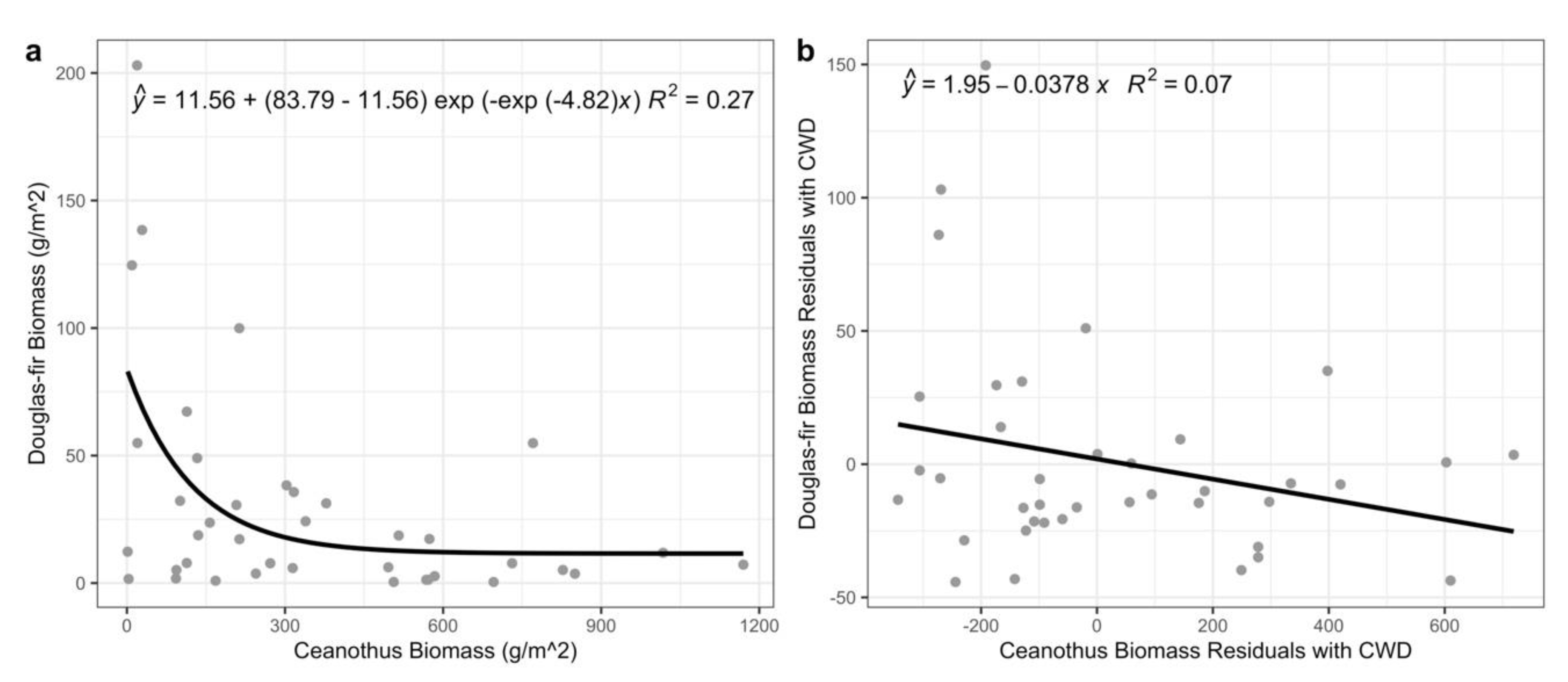{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites and Data
2.2. Field Methods
2.3. Analyses
3. Results
3.1. Ceanothus and Douglas-Fir Biomass
3.2. Soil Resources (C and N)
3.3. Ceanothus Effects on Soil Nitrogen Availability and Concomitant Effects on Douglas-Fir
3.4. Interactions between Ceanothus and Douglas-Fir
4. Discussion
4.1. Post-Fire Succession
4.2. Soil Biogeochemistry along Time since Fire and Climatic Water Deficit Gradients
4.3. Ceanothus, Climatic Water Deficit, and Implications for the Cycling of Nitrogen in the Klamath
4.4. Interactions between Ceanothus and Douglas-Fir
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson-Teixeira, K.J.; Miller, A.D.; Mohan, J.E.; Hudiburg, T.W.; Duval, B.D.; DeLucia, E.H. Altered dynamics of forest recovery under a changing climate. Glob. Chang. Biol. 2013, 19, 2001–2021. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.G.; Allen, C.D.; Anderson-Teixeira, K.; Aukema, B.H.; Bond-Lamberty, B.; Chini, L.; Clark, J.S.; Dietze, M.; Grossiord, C.; Hanbury-Brown, A.; et al. Pervasive shifts in forest dynamics in a changing world. Science 2020, 368. [Google Scholar] [CrossRef]
- Anderson, K.J. Temporal patterns in rates of community change during succession. Am. Nat. 2007, 169, 780–793. [Google Scholar] [CrossRef]
- Westerling, A.L.; Turner, M.G.; Smithwick, E.A.; Romme, W.H.; Ryan, M.G. Continued warming could transform Greater Yellowstone fire regimes by mid-21st century. Proc. Natl. Acad. Sci. USA 2011, 208, 13165–13170. [Google Scholar] [CrossRef] [Green Version]
- Davis, K.T.; Dobrowski, S.Z.; Higuera, P.E.; Holden, Z.A.; Veblen, T.T.; Rother, M.T.; Parks, S.A.; Sala, A.; Maneta, M.P. Wildfires and climate change push low-elevation forests across a critical climate threshold. Proc. Natl. Acad. Sci. USA 2019, 116, 6193–6198. [Google Scholar] [CrossRef] [Green Version]
- Stevens-Rumann, C.S.; Morgan, P. Tree regeneration following wildfires in the western U.S.: A review. Fire Ecol. 2019, 15, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Kenny, S.A.; Bennett, A.F.; Clarke, M.F.; Morgan, J.W. Time-since-fire and climate interact to affect the structural recovery of an Australian semi-arid plant community. Austral Ecol. 2018, 43, 456–469. [Google Scholar] [CrossRef] [Green Version]
- Tepley, A.J.; Thomann, E.; Veblen, T.T.; Perry, G.L.W.; Holz, A.; Partisis, J.; Kitzberger, T.; Anderson-Teixeira, K.J. Influences of fire-vegetation feedbacks and post-fire recovery rates on forest landscape vulnerability to altered fire regimes. J. Ecol. 2018, 106, 1925–1940. [Google Scholar] [CrossRef] [Green Version]
- Briles, C.; Whitlock, C.; Bartlein, P. Postglacial vegetation, fire, and climate history of the Siskiyou Mountains, Oregon, USA. Quat. Res. 2005, 64, 44–56. [Google Scholar] [CrossRef]
- Skinner, C.N.; Taylor, A.H.; Agee, J.K.; Briles, C.E.; Whitlock, C.L. Klamath Mountains Bioregion. In Fire in California’s Ecosystems; Van Wagtendonk, J.W., Sugihara, N.G., Stephens, S.L., Thode, A.E., Shaffer, K.E., Fites-Kaufman, J.A., Eds.; University of California Press: Oakland, CA, USA, 2006; pp. 173–196. [Google Scholar]
- Tepley, A.J.; Thompson, J.R.; Epstein, H.E.; Anderson-Teixeira, K.J. Vulnerability to forest loss through altered post-fire recovery dynamics in a warming climate in the Klamath Mountains. Glob. Chang. Biol. 2017, 23, 4117–4132. [Google Scholar] [CrossRef] [PubMed]
- Serra-Diaz, J.M.; Maxwell, C.; Lucash, M.S.; Scheller, R.M.; Laflower, D.M.; Miller, A.D.; Tepley, A.J.; Epstein, H.E.; Anderson-Teixeira, K.J.; Thompson, J.R. Disequilibrium of fire-prone forests sets the stage for a rapid decline in conifer dominance during the 21st century. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.R.; Spies, T.A.; Ganio, L.M. Reburn severity in managed and unmanaged vegetation in a large wildfire. Proc. Natl. Acad. Sci. USA 2007, 104, 10743–10748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wagtendonk, J.W.; Sugihara, N.G.; Stephens, S.L.; Thode, A.E.; Shaffer, K.E.; Fites-Kaufman, J.A.; Agee, J.K. Fire in California’s Ecosystems; University of California Press: Oakland, CA, USA, 2006. [Google Scholar]
- Odion, D.C.; Moritz, M.A.; DellaSala, D.A. Alternative community states maintained by fire in the Klamath Mountains, USA. J. Ecol. 2009, 98, 96–105. [Google Scholar] [CrossRef]
- Miller, A.D.; Thompson, J.R.; Tepley, A.J.; Anderson-Teixeira, K.J. Alternative stable equilibria and critical thresholds created by fire regimes and plant responses in a fire-prone community. Ecography 2018, 42, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Parra, A.; Moreno, J.M. Drought differentially affects the post-fire dynamics of seeders and resprouters in a Mediterranean shrubland. Sci. Total Environ. 2018, 626, 1219–1229. [Google Scholar] [CrossRef]
- Knapp, E.E.; Weatherspoon, C.P.; Skinner, C.N. Shrub seed banks in mixed conifer forests of Northern California and the role of fire in regulating abundance. Fire Ecol. 2012, 8, 32–48. [Google Scholar] [CrossRef]
- Peterson, C.E.; Hazard, J.W. Regional variation in growth response of coastal Douglas-fir to nitrogen fertilizer in the Pacific Northwest. For. Sci. 1990, 36, 625–640. [Google Scholar]
- Oakley, B.B.; North, M.P.; Franklin, J.F. Facilitative and competitive effects of N-fixing shrub on white fir saplings. For. Ecol. Manag. 2006, 223, 100–107. [Google Scholar] [CrossRef]
- Strothmann, R.O.; Roy, D.F. Regeneration of Douglas-Fir in the Klamath Mountain Region, California and Oregon; U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1984.
- Bormann, B.T.; Homann, P.S.; Darbyshire, R.L.; Morisette, B.A. Intense forest wildfire sharply reduces mineral soil C and N: The first direct evidence. Can. J. For. Res. 2008, 38, 2771–2783. [Google Scholar] [CrossRef]
- Oakley, B.B.; North, M.P.; Franklin, J.F. The effects of fire on soil nitrogen associated with patches of the actinorhizal shrub Ceanothus cordulatus. Plant Soil 2003, 254, 35–46. [Google Scholar] [CrossRef]
- Oakley, B.B.; North, M.P.; Franklin, J.F.; Hedlund, B.P.; Staley, J.T. Diversity and Distribution of Frankia Strains Symbiotic with Ceanothus in California. Appl. Environ. Microbiol. 2004, 70, 6444–6452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavitovski, J.; Newton, M. Ecological importance of snowbrush Ceanothus velutinus in the Oregon Cascades. Ecology 1968, 49, 1113–1145. [Google Scholar]
- Youngberg, C.T.; Wollum, A.G. Nitrogen accretion in developing Ceanothus velutinus stands. Soil Sci. Soc. Am. J. 1976, 40, 109–112. [Google Scholar] [CrossRef]
- Binkley, D.; Cromack, K.; Fredriksen, R.L. Nitrogen accretion and availability in some snowbrush ecosystems. For. Sci. 1982, 28, 720–724. [Google Scholar]
- Johnson, D.W. Soil properties beneath Ceanothus and Pine Stands in the eastern Sierra Nevada. Soil Sci. Soc. Am. J. 1995, 59, 918–924. [Google Scholar] [CrossRef]
- Johnson, D.W.; Murphy, J.F.; Susfalk, R.B.; Caldwell, T.G.; Miller, W.W.; Walker, R.F.; Powers, R.F. The effects of wildfire, salvage logging, and post-fire N-fixation on the nutrient budgets of a Sierran forest. For. Ecol. Manag. 2005, 220, 155–165. [Google Scholar] [CrossRef]
- Yelenik, S.; Perakis, S.; Hibbs, D. Regional constraints to biological nitrogen fixation in post-fire forest communities. Ecology 2013, 9, 739–750. [Google Scholar] [CrossRef] [Green Version]
- Irvine, D.R.; Hibbs, D.E.; Shatford, J.P.A. The relative importance of biotic and abiotic controls on young conifer growth after fire in the Klamath-Siskiyou Region. Northwest Sci. 2009, 83, 334–347. [Google Scholar] [CrossRef]
- Rodriguez-Barrueco, C.; Mackintosh, A.H.; Bond, G. Some effects of combined nitrogen on the nodule symbioses of Casuarina and Ceanothus. Plant Soil 1970, 33, 129–139. [Google Scholar] [CrossRef]
- Zavitovski, J.; Newton, M.; El-Hassan, B. Effects of snowbrush on growth of some conifers. J. For. 1969, 67, 242–246. [Google Scholar]
- Lopez-Ortiz, M.J.; Marcey, T.; Lucash, M.S.; Hibbs, D.; Shatford, P.A.; Thompson, J.R. Post-fire management affects species composition but not Douglas- fir regeneration in the Klamath Mountains. For. Ecol. Manag. 2019, 432, 1030–1040. [Google Scholar] [CrossRef]
- Zhang, J.; Oliver, W.W.; Busse, M.D. Growth and development of ponderosa pine on sites of contrasting productivities: Relative importance of stand density and shrub competition effects. Can. J. For. Res. 2006, 36, 2426–2438. [Google Scholar] [CrossRef]
- Hijmans, R.J.; van Etten, J. Raster: Geographic Data Analysis and Modeling, R Package Version 3.3-13. 2014.
- Lutz, J.A.; Van Wagtendonk, J.W.; Franklin, J.F. Climatic water deficit, tree species ranges, and climate change in Yosemite National Park. J. Biogeogr. 2010, 37, 936–950. [Google Scholar] [CrossRef]
- PRISM Climate Group, Oregon State University. Available online: http://prism.oregonstate.edu (accessed on 18 March 2015).
- Soil Survey Staff, Natural Resources Conservation Service, United States Department of Agriculture. Soil Survey Geographic (SSURGO). Available online: https://data.nal.usda.gov/dataset/soil-survey-geographic-database-ssurgo (accessed on 25 May 2015).
- Miller, J.D.; Knapp, E.E.; Key, C.H.; Skinner, C.N.; Isbell, C.J.; Creasy, R.M.; Sherlock, J.W. Calibration and validation of the relative differenced Normalized Burn Ratio (RdNBR) to three measures of fire severity in the Sierra Nevda and Klamath Mountains, CA, USA. Remote Sens. Environ. 2009, 113, 645–656. [Google Scholar] [CrossRef]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2006. [Google Scholar]
- Erickson, H.E.; Harrington, C.A. Conifer-Ceanothus interactions influence tree growth before and after shrub removal in a forest plantation in the western Cascade Mountains, USA. For. Ecol. Manag. 2006, 229, 183–194. [Google Scholar] [CrossRef]
- Johnson, D.W.; Walker, R.F.; McNulty, M.; Rau, B.M.; Miller, W.W. The long-term effects of wildfire and post-fire vegetation on Sierra Nevada forest soils. Forests 2012, 3, 398–416. [Google Scholar] [CrossRef] [Green Version]
- Swanson, M.E.; Franklin, J.F.; Beschta, R.L.; Crisafulli, C.M.; DellaSala, D.A.; Hutto, R.L.; Lindenmaver, D.B.; Swanson, F.J. The forgotten stage of forest succession: Early-successional ecosystems on forest sites. Front. Ecol. Environ. 2011, 9, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.R.; Wardle, D.A.; Bardgett, R.D.; Clarkson, B.D. The use of chronosequences in studies of ecological succession and soil development. J. Ecol. 2010, 98, 725–736. [Google Scholar] [CrossRef]
- Conard, S.G.; Jaramillo, A.E.; Cormack, K., Jr.; Rose, S. The Role of the Genus Ceanothus in Western Forest Ecosystems; U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and Range Experiment Station: Portland, OR, USA, 1985.
- Montwé, D.; Spiecker, H.; Hamann, A. Five decades of growth in a genetic field trial of Douglas-fir reveal trade-offs between productivity and drought-tolerance. Tree Genet. Genomes 2015, 11, 29. [Google Scholar] [CrossRef]
- Laflower, D.M.; Hurteau, M.D.; Koch, G.W.; North, M.P.; Hungate, B.A. Climate-driven changes in forest succession and the influence of management on forest carbon dynamics in the Puget Lowlands of Washington State, USA. For. Ecol. Manag. 2016, 362, 194–204. [Google Scholar] [CrossRef] [Green Version]
- McNabb, D.H.; Cromack, K., Jr. Dinitrogen N-fixation by a mature Ceanothus velutinus (Dougl.) stand in the western Oregon Cascades. Can. J. Microbiol. 1983, 29, 1014–1021. [Google Scholar] [CrossRef]
- Rother, M.T.; Veblen, T.T. Limited conifer regeneration following wildfires in dry ponderosa pine forests of the Colorado Front Range. Ecosphere 2016, 7, e01594. [Google Scholar] [CrossRef]
- Adams, M.A.; Turnbull, T.L.; Sprent, J.I.; Buchmann, N. Legumes are different: Leaf nitrogen, photosynthesis, and water use efficiency. Proc. Natl. Acad. Sci. USA 2016, 113, 4098–4103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köster, K.; Köster, E.; Orumaa, A.; Parro, K.; Jōgiste, K.; Berninger, F.; Pumpanen, J.; Metslaid, M. How time since forest fire affects stand structure, soil physical-chemical properties and soil CO2 efflux in hemiboreal Scots Pine forest fire chronosequence? Forests 2016, 7, 201. [Google Scholar] [CrossRef] [Green Version]
- Marion, G.M.; Black, C.H. Potentially available nitrogen and phosphorus along a chaparral fire cycle chronosequence. Soil Sci. Soc. Am. J. 1988, 52, 1155–1162. [Google Scholar] [CrossRef]
- Page-Dumrose, D.; Jurgensen, M.F.; Harvey, A.E. Fire and fire-suppression impacts on forest-soil carbon. In The Potential of U.S. Forest Soils to Sequester Carbon and Mitigate the Greenhouse Effect; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Pellegrini, A.F.A.; Jackson, R.B. The long and short of it: A review of the timescales of how fire affects soils using the pulse-press framework. Adv. Ecol. Res. 2020, 62, 147–171. [Google Scholar]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Fire effects on temperate forest soil C and N storage. Ecol. Appl. 2011, 21, 1189–1201. [Google Scholar] [CrossRef] [Green Version]
- Perakis, S.S.; Tepley, A.J.; Compton, J.E. Disturbance and topography shape nitrogen availability and δ15 N over long-term forest succession. Ecosystems 2015, 18, 573–588. [Google Scholar] [CrossRef]
- Cross, A.; Perakis, S.S. Tree species and soil nutrient profiles in old-growth forests of the Oregon Coast Range. Can. J. For. Res. 2011, 41, 195–210. [Google Scholar] [CrossRef]
- Homann, P.S.; Bormann, B.T.; Darbyshire, R.L.; Morrissette, B.A. Forest soil carbon and nitrogen losses associated with wildfire and prescribed fire. Soil Sci. Soc. Am. J. 2011, 75, 1926–1934. [Google Scholar] [CrossRef]
- Borchers, J.G.; Perry, D.A. The influence of soil texture and aggregation on carbon and nitrogen dynamics in southwest Oregon forests and clearcuts. Can. J. For. Res. 1992, 22, 298–305. [Google Scholar] [CrossRef]
- Morford, S.L.; Houlton, B.Z.; Dahlgren, R.A. Geochemical and tectonic uplift controls on rock nitrogen inputs across terrestrial ecosystems. Glob. Biogeochem. Cycles 2016, 30, 333–349. [Google Scholar] [CrossRef] [Green Version]
- Post, W.M.; Emanuel, W.R.; Zinke, P.J.; Stangenberger, A.G. Soil carbon pools and world life zones. Nature 1982, 298, 156–159. [Google Scholar] [CrossRef]
- Post, W.M.; Pastor, J.; Zinke, P.J.; Stangenberger, A.G. Global patterns of soil nitrogen storage. Nature 1985, 317, 613–616. [Google Scholar] [CrossRef]
- McLauchlan, K.K.; Higuera, P.E.; Gavin, D.G.; Perakis, S.S.; Mack, M.C.; Alexander, H.; Battles, J.; Biondi, F.; Buma, B.; Colombaroli, D.; et al. Reconstructing disturbances and their biogeochemical consequences over multiple timescales. BioScience 2014, 64, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Waksman, S.A.; Tenney, F.G. The composition of natural organic materials and their decomposition in the soil. Soil Sci. 1927, 24, 257–384. [Google Scholar]
- Erickson, H.E.; Soto, P.; Johnson, D.W.; Roath, B.; Hunsaker, C. Effects of vegetation patches on soil nutrient pools and fluxes within a mixed-conifer forest. For. Sci. 2005, 51, 211–220. [Google Scholar]
- Busse, M.D.; Cochran, P.H.; Barrett, J.W. Changes in ponderosa pine productivity following removal of understory vegetation. Soil Sci. Soc. Am. J. 1996, 60, 1614–1621. [Google Scholar] [CrossRef] [Green Version]
- Binkley, D. How nitrogen-fixing trees change soil carbon. In Tree Species Effects on Soils: Implications for Global Change; Springer: Dordrecht, The Netherlands, 2005; pp. 155–164. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cinoğlu, D.; Epstein, H.E.; Tepley, A.J.; Anderson-Teixeira, K.J.; Thompson, J.R.; Perakis, S.S. Climatic Aridity Shapes Post-Fire Interactions between Ceanothus spp. and Douglas-Fir (Pseudotsuga menziesii) across the Klamath Mountains. Forests 2021, 12, 1567. https://doi.org/10.3390/f12111567
Cinoğlu D, Epstein HE, Tepley AJ, Anderson-Teixeira KJ, Thompson JR, Perakis SS. Climatic Aridity Shapes Post-Fire Interactions between Ceanothus spp. and Douglas-Fir (Pseudotsuga menziesii) across the Klamath Mountains. Forests. 2021; 12(11):1567. https://doi.org/10.3390/f12111567
Chicago/Turabian StyleCinoğlu, Damla, Howard E. Epstein, Alan J. Tepley, Kristina J. Anderson-Teixeira, Jonathan R. Thompson, and Steven S. Perakis. 2021. "Climatic Aridity Shapes Post-Fire Interactions between Ceanothus spp. and Douglas-Fir (Pseudotsuga menziesii) across the Klamath Mountains" Forests 12, no. 11: 1567. https://doi.org/10.3390/f12111567
APA StyleCinoğlu, D., Epstein, H. E., Tepley, A. J., Anderson-Teixeira, K. J., Thompson, J. R., & Perakis, S. S. (2021). Climatic Aridity Shapes Post-Fire Interactions between Ceanothus spp. and Douglas-Fir (Pseudotsuga menziesii) across the Klamath Mountains. Forests, 12(11), 1567. https://doi.org/10.3390/f12111567