Assisted Migration Field Tests in Canada and Mexico: Lessons, Limitations, and Challenges


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Key Assisted Migration Definitions
- (a)
- Assisted migration within current geographic range, for simplicity referred to herein as assisted migration within range, when it involves the movement of populations within a species’ current natural distribution. It has also been referred to as assisted gene flow [40] or assisted population migration [37].
- (b)
- Assisted migration outside of current geographic range, referred herein as assisted migration outside of range, when it involves the movement of populations to locations just beyond a species’ current distribution. This form of assisted migration has also been called assisted colonization [50] or assisted range expansion [37].
3. Field Tests Examined
3.1. Canada. Pinus albicaulis
Materials and Climate Data
3.2. Canada. Picea glauca × engelmannii Hybrid Complex (Interior Spruce)
Materials and Climate Data
3.3. México. Abies religiosa (Sacred Fir, Oyamel)
4. Lessons
4.1. Pinus albicaulis Can Be Migrated North of Its Current Species Distribution in Canada within the Predicted Climatic Niche If Adequate Snowpack Is Present
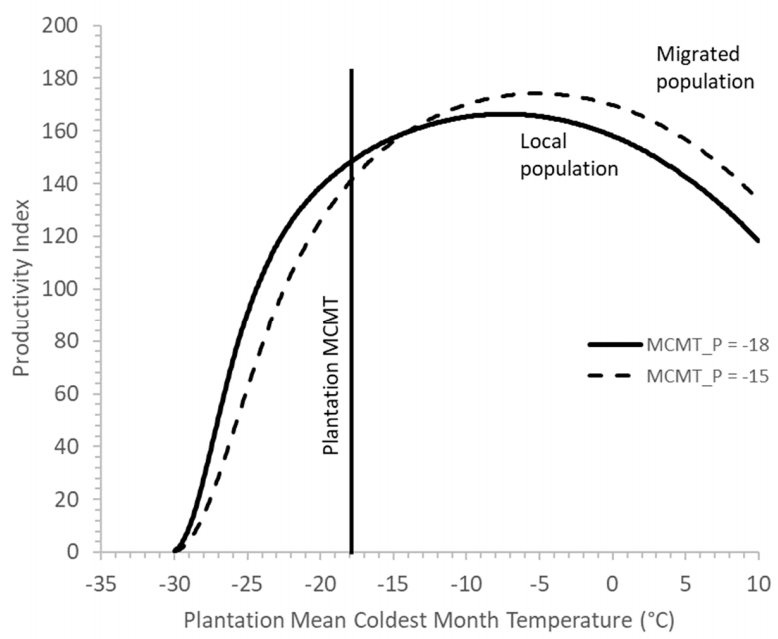
4.2. Picea glauca × engelmannii Hybrid Complex
4.3. México. Abies religiosa. The Species Performs Well When Moved 400 m Upward and Local Shrubs Are Used as Nurse Plants
5. Limitations
5.1. Canada. Pinus albicaulis
5.2. Canada. Picea glauca × engelmannii Hybrid Complex
How Far Is Too Far? Determining a Suitable Migration Distance
5.3. México. Abies religiosa. Too Short Observational Period
6. Challenges
6.1. Making Large Multisite Provenance Field Trials More Affordable
6.2. Replications over Time
6.3. The Dilemma of Species Prioritization in Megabiodiverse Countries
6.4. When Poleward Migration Is Not an Option
6.5. Tackling Uncertainty by Promoting Genetic Diversification
7. Mitigating Risks
7.1. Introduction of Species That Might Become Invasive
7.2. Mismatch of Biotic Interactions
7.3. Outbreeding Depression
7.4. Balancing the Risks: The Core Dilemma
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Anchor Points for P. glauca × engelmannii Analysis
Appendix A.2. Universal Transfer Function and Ghost Test Site for P. glauca × engelmannii Analysis
Appendix A.3. Appendix References
References
- Ledig, F.T.; Kitzmiller, J.H. Genetic strategies for reforestation in the face of global climate change. For. Ecol. Manag. 1992, 50, 153–169. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Wykoff, W.R.; Ying, C.C. Physiologic plasticity, evolution, and impacts of a changing climate on Pinus contorta. Clim. Chang. 2001, 50, 355–376. [Google Scholar] [CrossRef]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-McLane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2008, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Rehfeldt, G.E.; Jaquish, B.C.; López-Upton, J.; Sáenz-Romero, C.; St Clair, J.B.; Leites, L.P.; Joyce, D.G. Comparative genetic responses to climate for the varieties of Pinus ponderosa and Pseudotsuga menziesii: Realized climate niches. For. Ecol. Manag. 2014, 324, 126–137. [Google Scholar] [CrossRef]
- Rehfeldt, G.E. Ecological genetics of Pinus contorta from the Rocky Mountains (USA): A synthesis. Silv. Genet. 1988, 37, 131–135. [Google Scholar]
- Aitken, S.N.; Bemmels, J.B. Time to get moving: Assisted gene flow of forest trees. Evol. Appl. 2016, 9, 271–290. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Leites, L.P.; Joyce, D.G.; Weiskittel, A.R. Role of population genetics in guiding ecological responses to climate. Glob. Chang. Biol. 2017, 24, 858–868. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Guzmán-Reyna, R.R.; Rehfeldt, G.E. Altitudinal genetic variation among Pinus oocarpa populations in Michoacán, Mexico: Implications for seed zoning, conservation, tree breeding and global warming. For. Ecol. Manag. 2006, 229, 340–350. [Google Scholar] [CrossRef]
- Alvarez-Maldini, C.; Acevedo, M.; Dumroese, R.K.; González, M.; Cartes, E. Intraspecific variation in drought response of three populations of Cryptocarya alba and Persea lingue, two native species from mediterranean central Chile. Front. Plant Sci. 2020, 11, 1–15. [Google Scholar] [CrossRef]
- Peñuelas, J.; Ogaya, R.; Boada, M.S.; Jump, A. Migration, invasion and decline: Changes in recruitment and forest structure in a warming-linked shift of European beech forest in Catalonia (NE Spain). Ecography 2007, 30, 829–837. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; Peters, M. Estimating potential habitat for 134 eastern US tree species under six climate scenarios. For. Ecol. Manag. 2008, 254, 390–406. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Crookston, N.L.; Sáenz-Romero, C.; Campbell, E.M. North American vegetation model for land-use planning in a changing climate: A solution to large classification problems. Ecol. Appl. 2012, 22, 119–141. [Google Scholar] [CrossRef]
- Rehfeldt, G.E.; Jaquish, B.C.; Sáenz-Romero, C.; Joyce, D.G.; Leites, L.P.; St Clair, J.B.; López-Upton, J. Comparative genetic responses to climate in the varieties of Pinus ponderosa and Pseudotsuga menziesii: Reforestation. For. Ecol. Manag. 2014, 324, 147–157. [Google Scholar] [CrossRef]
- Gómez-Pineda, E.; Sáenz-Romero, C.; Ortega-Rodríguez, J.M.; Blanco-García, A.; Madrigal-Sánchez, X.; Lindig-Cisneros, R.; Lopez-Toledo, L.; Pedraza-Santos, M.E.; Rehfeldt, G.E. Suitable climatic habitat changes for Mexican conifers along altitudinal gradients under climatic change scenarios. Ecol. Appl. 2020, 30, e02041. [Google Scholar] [CrossRef] [PubMed]
- Lenoir, J.; Gégout, J.-C.; Marquet, P.; De Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Alfaro-Ramírez, F.U.; Arredondo-Moreno, J.T.; Pérez-Suárez, M.; Endara-Agramont, Á.R. Pinus hartwegii Lindl. treeline ecotone: Structure and altitudinal limits at Nevado de Toluca, Mexico. Rev. Chapingo. Ser. Cienc. For. Ambiente 2017, 23, 261–273. [Google Scholar] [CrossRef]
- Trant, A.; Higgs, E.; Starzomski, B.M. A century of high elevation ecosystem change in the Canadian Rocky Mountains. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, R.I.; Fady, B.; Vendramin, G.G.; Dawson, I.K.; Fleming, R.A.; Sáenz-Romero, C.; Lindig-Cisneros, R.A.; Murdock, T.; Vinceti, B.; Navarro, C.M. The role of forest genetic resources in responding to biotic and abiotic factors in the context of anthropogenic climate change. For. Ecol. Manag. 2014, 333, 76–87. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.T. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Allen, C.; Breshears, D.; McDowell, N. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Hogg, E.H.; Michaelian, M.; Hook, T.I.; Undershultz, M.E. Recent climatic drying leads to age-independent growth reductions of white spruce stands in western Canada. Glob. Chang. Biol. 2017, 23, 5297–5308. [Google Scholar] [CrossRef] [PubMed]
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.S.; Hunt, J.M.; Penuelas, J. Rapid climate change-related growth decline at the southern range edge of Fagus sylvatica. Glob. Chang. Biol. 2006, 12, 2163–2174. [Google Scholar] [CrossRef]
- Mátyás, C. Forecasts needed for retreating forests. Nature 2010, 464, 1271. [Google Scholar] [CrossRef] [PubMed]
- Mátyás, C.; Berki, I.; Czúcz, B.; Gálos, B.; Móricz, N.; Rasztovits, E. Future of beech in Southeast Europe from the perspective of evolutionary ecology. Acta Silv. Lignaria Hung. 2010, 6, 91–110. [Google Scholar]
- Fréjaville, T.; Fady, B.; Kremer, A.; Ducousso, A.; Benito Garzón, M. Inferring phenotypic plasticity and population responses to climate across tree species ranges using forest inventory data. Glob. Ecol. Biogeogr. 2019, 28, 1259–1271. [Google Scholar] [CrossRef]
- Gonzalez, A.; Ronce, O.; Ferriere, R.; Hochberg, M.E. Evolutionary rescue: An emerging focus at the intersection between ecology and evolution. Philosoph. Trans. R. Soc. B 2012, 368, 1–8. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W.; Sherman-Broyles, S.L. Factors influencing levels of genetic diversity in woody plant species. New For. 1992, 6, 95–124. [Google Scholar] [CrossRef]
- St Clair, J.B.; Howe, G.T. Genetic maladaptation of coastal Douglas-fir seedlings to future climates. Glob. Chang. Biol. 2007, 13, 1441–1454. [Google Scholar] [CrossRef]
- Michaelian, M.; Hogg, E.H.; Hall, R.J.; Arsenault, E. Massive mortality of aspen following severe drought along the southern edge of the Canadian boreal forest. Glob. Chang. Biol. 2011, 17, 2084–2094. [Google Scholar] [CrossRef]
- Bucharova, A. Assisted migration within species range ignores biotic interactions and lacks evidence. Restorat. Ecol. 2017, 25, 14–18. [Google Scholar] [CrossRef]
- Richardson, D.M.; Hellmann, J.J.; McLachlan, J.S.; Sax, D.F.; Schwartz, M.W.; Gonzalez, P.; Brennan, E.J.; Camacho, A.; Root, T.L.; Sala, O.E. Multidimensional evaluation of managed relocation. Proc. Natl. Acad. Sci. USA 2009, 106, 9721–9724. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, J.S.; Hellmann, J.J.; Schwartz, M.W. A framework for debate of assisted migration in an era of climate change. Conserv. Biol. 2007, 21, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; O’Neill, G.A.; Aitken, S.N. Integrating environmental and genetic effects to predict responses of tree populations to climate. Ecol. Appl. 2010, 20, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Gray, L.K.; Gylander, T.; Mbogga, M.S.; Chen, P.-y.; Hamann, A. Assisted migration to address climate change: Recommendations for aspen reforestation in western Canada. Ecol. Appl. 2011, 21, 1591–1603. [Google Scholar] [CrossRef] [PubMed]
- Gray, L.K.; Hamann, A. Strategies for reforestation under uncertain future climates: Guidelines for Alberta, Canada. PLoS ONE 2011, 6, e22977. [Google Scholar] [CrossRef]
- Leech, S.M.; Almuedo, P.L.; O’Neill, G. Assisted migration: Adapting forest management to a changing climate. J. Ecosyst. Manag. 2011, 12, 18–34. Available online: http://jem.forrex.org/index.php/jem/article/view/91/98 (accessed on 15 November 2020).
- Hewitt, N.; Klenk, N.; Smith, A.L.; Bazely, D.R.; Yan, N.; Wood, S.; MacLellan, J.I.; Lipsig-Mumme, C.; Henriques, I. Taking stock of the assisted migration debate. Biol. Conserv. 2011, 144, 2560–2572. [Google Scholar] [CrossRef]
- Pedlar, J.H.; McKenney, D.W.; Aubin, I.; Beardmore, T.; Beaulieu, J.; Iverson, L.; O’Neill, G.A.; Winder, R.S.; Ste-Marie, C. Placing forestry in the assisted migration debate. BioScience 2012, 62, 835–842. [Google Scholar] [CrossRef]
- Aitken, S.N.; Whitlock, M.C. Assisted gene flow to facilitate local adaptation to climate change. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 367–388. [Google Scholar] [CrossRef]
- Lu, P.; Parker, W.H.; Cherry, M.; Colombo, S.; Parker, W.C.; Man, R.; Roubal, N. Survival and growth patterns of white spruce (Picea glauca [M oench] Voss) rangewide provenances and their implications for climate change adaptation. Ecol. Evol. 2014, 4, 2360–2374. [Google Scholar] [CrossRef] [PubMed]
- McKenney, D.W.; Pedlar, J.H.; Lawrence, K.; Papadopol, P.; Campbell, K.; Hutchinson, M.F. Change and evolution in the plant hardiness zones of Canada. BioScience 2014, 64, 341–350. [Google Scholar] [CrossRef]
- Castellanos-Acuña, D.; Lindig-Cisneros, R.; Sáenz-Romero, C. Altitudinal assisted migration of Mexican pines as an adaptation to climate change. Ecosphere 2015, 6, 1–16. [Google Scholar] [CrossRef]
- Koralewski, T.E.; Wang, H.-H.; Grant, W.E.; Byram, T.D. Plants on the move: Assisted migration of forest trees in the face of climate change. For. Ecol. Manag. 2015, 344, 30–37. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Mendoza-Maya, E.; Gómez-Pineda, E.; Blanco-García, A.; Endara-Agramont, A.R.; Lindig-Cisneros, R.; López-Upton, J.; Trejo-Ramírez, O.; Wehenkel, C.; Cibrián-Tovar, D. Recent evidence of Mexican temperate forest decline and the need for ex situ conservation, assisted migration, and translocation of species ensembles as adaptive management to face projected climatic change impacts in a megadiverse country. Can. J. For. Res. 2020, 50, 843–854. [Google Scholar] [CrossRef]
- Wang, T.; Hamann, A.; Yanchuk, A.; O’neill, G.; Aitken, S. Use of response functions in selecting lodgepole pine populations for future climates. Glob. Chang. Biol. 2006, 12, 2404–2416. [Google Scholar] [CrossRef]
- Nigh, G. Mitigating the effects of climate change on lodgepole pine site height in British Columbia, Canada, with a transfer function. For. Int. J. For. Res. 2014, 87, 377–387. [Google Scholar] [CrossRef][Green Version]
- O’Neill, G.; Wang, T.; Ukrainetz, N.; Charleson, L.; McAuley, L.; Yanchuk, A.; Zedel, S. A Proposed Climate-Based Seed Transfer System for British Columbia; Technical Report 099; Ministry of Forests, Lands and Natural Resource Operations: Victoria, BC, Canada, 2017.
- Peterson St-Laurent, G.; Hagerman, S.; Findlater, K.M.; Kozak, R. Public trust and knowledge in the context of emerging climate-adaptive forestry policies. J. Environ. Manag. 2019, 242, 474–486. [Google Scholar] [CrossRef]
- Ledig, F.T.; Rehfeldt, G.E.; Sáenz-Romero, C.; Flores-López, C. Projections of suitable habitat for rare species under global warming scenarios. Am. J. Bot. 2010, 97, 970–987. [Google Scholar] [CrossRef]
- Ukrainetz, N.K.; O’Neill, G.A.; Jaquish, B. Comparison of fixed and focal point seed transfer systems for reforestation and assisted migration: A case study for interior spruce in British Columbia. Can. J. For. Res. 2011, 41, 1452–1464. [Google Scholar] [CrossRef]
- Warwell, M.V.; Rehfeldt, G.E.; Crookston, N.L. Modeling contemporary climate profiles of whitebark pine (Pinus albicaulis) and predicting responses to global warming. In Proceedings of the Conference Whitebark Pine: A Pacific Coast Perspective; Goheen, E., Ed.; R6-NR-FHP-2007-01; US Department of Agriculture, Forest Service: Ashland, OR, USA, 2007; pp. 139–142. Available online: https://www.fs.usda.gov/treesearch/pubs/33997 (accessed on 15 November 2020).
- McLane, S.C.; Aitken, S.N. Whitebark pine (Pinus albicaulis) assisted migration potential: Testing establishment north of the species range. Ecol. Appl. 2012, 22. [Google Scholar] [CrossRef] [PubMed]
- Seliger, B.J.; McGill, B.J.; Svenning, J.C.; Gill, J.L. Widespread underfilling of the potential ranges of North American trees. J. Biogeogr. 2020. [Google Scholar] [CrossRef]
- Statistics Canada. Human Activity and the Environment: Forests in Canada 2017; Catalogue No. 16 201 X; Statistics Canada: Ottawa, QC, Canada, 2018.
- O’Neill, G.A.; Stoehr, M.; Jaquish, B. Quantifying safe seed transfer distance and impacts of tree breeding on adaptation. For. Ecol. Manag. 2014, 328, 122–130. [Google Scholar] [CrossRef]
- Wang, T.; Hamann, A.; Spittlehouse, D.; Carroll, C. Locally downscaled and spatially customizable climate data for historical and future periods for North America. PLoS ONE 2016, 11, e0156720. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, G.A.; Hamann, A.; Wang, T. Accounting for population variation improves estimates of the impact of climate change on species’ growth and distribution. J. Appl. Ecol. 2008, 45, 1040–1049. [Google Scholar] [CrossRef]
- St. Clair, J.B.; Howe, G.T.; Kling, J.G. The 1912 Douglas-Fir heredity study: Long-term effects of climatic transfer distance on growth and survival. J. For. 2020, 118, 1–13. [Google Scholar] [CrossRef]
- Rzedowski, J. Vegetación de México; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Ciudad de México, México, 2006. [Google Scholar]
- Sánchez-Velásquez, L.R.; Domínguez-Hernández, D.; Pineda-López, M.d.R.; Lara-González, R. Does Baccharis conferta shrub act as a nurse plant to the Abies religiosa seedling? Open For. Sci. J. 2011, 4, 67–70. [Google Scholar] [CrossRef]
- Guzmán-Santiago, J.C.; Aguirre-Calderón, O.A.; Jiménez-Pérez, J.; Vargas-Larreta, B. Volume estimation of Abies religiosa (Kunth) Schltdl. & Cham. In different states Mexico. Colomb. For. 2020, 23, 99–113. [Google Scholar] [CrossRef]
- Anderson, J.; Brower, L. Freeze-protection of overwintering monarch butterflies in Mexico: Critical role of the forest as a blanket and an umbrella. Ecol. Entomol. 1996, 21, 107–116. [Google Scholar] [CrossRef]
- Montoya, E.; Guzmán-Plazola, R.A.; López-Mata, L. Fragmentation dynamics in an Abies religiosa forest of central Mexico. Can. J. For. Res. 2020, 50, 680–688. [Google Scholar] [CrossRef]
- Carbajal-Navarro, A.; Navarro-Miranda, E.; Blanco, A.; Cruzado-Vargas, A.L.; Gómez-Pineda, E.; Zamora-Sánchez, C.; Pineda-García, F.; O’Neill, G.; Gómez-Romero, M.; Lindig-Cisneros, R. Ecological restoration of Abies religiosa forests using nurse plants and assisted migration in the Monarch Butterfly Biosphere Reserve, Mexico. Front. Ecol. Evol. 2019, 7, 421. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Rehfeldt, G.E.; Crookston, N.L.; Duval, P.; St-Amant, R.; Beaulieu, J.; Richardson, B.A. Spline models of contemporary, 2030, 2060 and 2090 climates for Mexico and their use in understanding climate-change impacts on the vegetation. Clim. Chang. 2010, 102, 595–623. [Google Scholar] [CrossRef]
- Peterson, S.-L.G.; Hagerman, S.; Kozak, R. What risks matter? Public views about assisted migration and other climate-adaptive reforestation strategies. Clim. Chang. 2018, 151, 573–587. [Google Scholar] [CrossRef]
- Barrows, C.W.; Ramirez, A.R.; Sweet, L.C.; Morelli, T.L.; Millar, C.I.; Frakes, N.; Rodgers, J.; Mahalovich, M.F. Validating climate-change refugia: Empirical bottom-up approaches to support management actions. Front. Ecol. Environ. 2020, 18, 298–306. [Google Scholar] [CrossRef]
- Palmer, C.; Larson, B.M. Should we move the whitebark pine? Assisted migration, ethics and global environmental change. Environ. Values 2014, 23, 641–662. [Google Scholar] [CrossRef]
- Pickles, B.J.; Twieg, B.D.; O’Neill, G.A.; Mohn, W.W.; Simard, S.W. Local adaptation in migrated interior Douglas-fir seedlings is mediated by ectomycorrhizas and other soil factors. New Phytol. 2015, 207, 858–871. [Google Scholar] [CrossRef]
- Brower, L.P.; Williams, E.H.; Jaramillo-López, P.; Kust, D.R.; Slayback, D.A.; Ramírez, M.I. Butterfly mortality and salvage logging from the march 2016 storm in the Monarch Butterfly Biosphere Reserve in Mexico. Am. Entomol. 2017, 63, 151–164. [Google Scholar] [CrossRef][Green Version]
- Campbell, R.K. Use of phenology for examining provenance transfers in reforestation of Douglas-fir. J. Appl. Ecol. 1974. [Google Scholar] [CrossRef]
- Raymond, C.; Lindgren, D. Genetic flexibility-a model for determining the range of suitable environments for a seed source. Silv. Genet. 1990, 39, 112–120. [Google Scholar]
- Mátyás, C. Modeling climate change effects with provenance test data. Tree Physiol. 1994, 14, 797–804. [Google Scholar] [CrossRef]
- Carter, K. Provenance tests as indicators of growth response to climate change in 10 north temperate tree species. Can. J. For. Res. 1996, 26, 1089–1095. [Google Scholar] [CrossRef]
- Leites, L.P.; Robinson, A.P.; Rehfeldt, G.E.; Marshall, J.D.; Crookston, N.L. Height-growth response to climatic changes differs among populations of Douglas-fir: A novel analysis of historic data. Ecol. Appl. 2012, 22, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Breed, M.F.; Stead, M.G.; Ottewell, K.M.; Gardner, M.G.; Lowe, A.J. Which provenance and where? Seed sourcing strategies for revegetation in a changing environment. Conserv. Genet. 2013, 14, 1–10. [Google Scholar] [CrossRef]
- Sáenz-Romero, C.; Lamy, J.B.; Ducousso, A.; Musch, B.; Ehrenmann, F.; Delzon, S.; Cavers, S.; Chałupka, W.; Dağdaş, S.; Hansen, J.K. Adaptive and plastic responses of Quercus petraea populations to climate across Europe. Glob. Chang. Biol. 2017, 23, 2831–2847. [Google Scholar] [CrossRef] [PubMed]
- Berlin, M.; Persson, T.; Jansson, G.; Haapanen, M.; Ruotsalainen, S.; Bärring, L.; Andersson Gull, B. Scots pine transfer effect models for growth and survival in Sweden and Finland. Silv. Fenn. 2016, 50, 1562. [Google Scholar] [CrossRef]
- O’Neill, G.; Carlson, M.; Berger, V.; Ukrainetz, N. Assisted Migration Adaptation Trial: Workplan; BC Ministry of Forests, Lands and Natural Resource Operations: Vernon, BC, Canada, 2013.
- Shutyaev, A.; Giertych, M. Height growth variation in a comprehensive Eurasian provenance experiment of (Pinus sylvestris L.). Silv. Genet. 1998, 46, 332–348. [Google Scholar]
- Shutyaev, A.; Giertych, M. Genetic subdivisions of the range of Scots pine (Pinus sylvestris L.) based on a transcontinental provenance experiment. Silv. Genet. 2000, 49, 137–151. [Google Scholar]
- Sáenz-Romero, C.; Kremer, A.; Nagy, L.; Újvári-Jármay, É.; Ducousso, A.; Kóczán-Horváth, A.; Hansen, J.K.; Mátyás, C. Common garden comparisons confirm inherited differences in sensitivity to climate change between forest tree species. PeerJ 2019, 7, e6213. [Google Scholar] [CrossRef]
- Marris, E. Planting the forest of the future: While conservation biologists debate whether to move organisms threatened by the warming climate, one forester in British Columbia is already doing it. Nature 2009, 459, 906–908. [Google Scholar] [CrossRef]
- Fu, Y.-B.; Clarke, G.P.Y.; Namkoong, G.; Yanchuk, A.D. Incomplete block designs for genetic testing: Statistical efficiencies of estimating family means. Can. J. For. Res. 1998, 28, 977–986. [Google Scholar] [CrossRef]
- Yeaman, S.; Hodgins, K.A.; Lotterhos, K.E.; Suren, H.; Nadeau, S.; Degner, J.C.; Nurkowski, K.A.; Smets, P.; Wang, T.; Gray, L.K. Convergent local adaptation to climate in distantly related conifers. Science 2016, 353, 1431–1433. [Google Scholar] [CrossRef] [PubMed]
- Sáenz-Romero, C.; Rehfeldt, G.E.; Duval, P.; Lindig-Cisneros, R.A. Abies religiosa habitat prediction in climatic change scenarios and implications for monarch butterfly conservation in Mexico. For. Ecol. Manag. 2012, 275, 98–106. [Google Scholar] [CrossRef]
- Etterson, J.R.; Cornett, M.W.; White, M.A.; Kavajecz, L.C. Assisted migration across fixed seed zones detects adaptation lags in two major North American tree species. Ecol. Appl. 2020, e02092. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Pedlar, J.; Peters, M.; McKenney, D.; Iverson, L.; Matthews, S.; Adams, B. Combining US and Canadian forest inventories to assess habitat suitability and migration potential of 25 tree species under climate change. Divers. Distrib. 2020, 26, 1142–1159. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef]
- Lin, B.B. Resilience in agriculture through crop diversification: Adaptive management for environmental change. BioScience 2011, 61, 183–193. [Google Scholar] [CrossRef]
- Markowitz, H.M. Portafolio selection. Finance 1952, 7, 77–91. [Google Scholar]
- Yanchuk, A.D. A quantitative framework for breeding and conservation of forest tree genetic resources in British Columbia. Can. J. For. Res. 2001, 31, 566–576. [Google Scholar] [CrossRef]
- Ministry of Forests, Lands, Natural Resource Operations and Rural Development. Chief Forester Standards for Seed Use 2020; Ministry of Forests, Lands, Natural Resource Operations: Victoria, BC, Canada, 2020. [Google Scholar]
- Sgrò, C.M.; Lowe, A.J.; Hoffmann, A.A. Building evolutionary resilience for conserving biodiversity under climate change. Evol. Appl. 2011, 4, 326–337. [Google Scholar] [CrossRef]
- Crowe, K.A.; Parker, W.H. Using portfolio theory to guide reforestation and restoration under climate change scenarios. Clim. Chang. 2008, 89, 355–370. [Google Scholar] [CrossRef]
- Ricciardi, A.; Simberloff, D. Assisted colonization is not a viable conservation strategy. Trends Ecol. Evol. 2009, 24, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Seabrook, L.; Mcalpine, C.A.; Bowen, M.E. Restore, repair or reinvent: Options for sustainable landscapes in a changing climate. Landsc. Urban Plan. 2011, 100, 407–410. [Google Scholar] [CrossRef]
- Mueller, J.M.; Hellmann, J.J. An assessment of invasion risk from assisted migration. Conserv. Biol. 2008, 22, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Daehler, C.C. The taxonomic distribution of invasive angiosperm plants: Ecological insights and comparison to agricultural weeds. Biol. Conserv. 1998, 84, 167–180. [Google Scholar] [CrossRef]
- Kowarik, I. Time lags in biological invasions with regard to the success and failure of alien species. In Plant Invasions: General Aspects and Special Problems; Pyšek, P., Rejmánek, K.P.M., Wade, M., Eds.; SPB Academic Publishing: Amsterdam, The Netherlands, 1995; pp. 15–38. [Google Scholar]
- Binggeli, P. Time−Lag between Introduction, Establishment and Rapid Spread of Introduced Environmental Weeds. In Proceedings of the III International Weed Science Congress, MS no. 8. International Weed Science Society, Oxford, UK, 6–11 June 2001. [Google Scholar]
- Crooks, J.A. Lag times and exotic species: The ecology and management of biological invasions in slow-motion1. Ecoscience 2005, 12, 316–329. [Google Scholar] [CrossRef]
- Camarretta, N.; Harrison, P.A.; Bailey, T.; Davidson, N.; Lucieer, A.; Hunt, M.; Potts, B.M. Stability of species and provenance performance when translocated into different community assemblages. Restor. Ecol. 2020, 28, 447–458. [Google Scholar] [CrossRef]
- Remke, M.J.; Hoang, T.; Kolb, T.; Gehring, C.; Johnson, N.C.; Bowker, M.A. Familiar soil conditions help Pinus ponderosa seedlings cope with warming and drying climate. Restor. Ecol. 2020, 28, S344–S354. [Google Scholar] [CrossRef]
- García-Hernández, M.d.l.Á.; Toledo-Aceves, T. Is there potential in elevational assisted migration for the endangered Magnolia vovidesii? J. Nat. Conserv. 2020, 53, 125782. [Google Scholar] [CrossRef]
- World Meteorological Organization. WMO Provisional Statement on the State of the Global Climate in 2019; WMO: Geneva, Switzerland, 2020; p. 34. [Google Scholar]
- Guzmán-Aguilar, G.; Carbajal-Navarro, A.; Sáenz-Romero, C.; Herrerías-Diego, Y.; López-Toledo, L.; Blanco-García, A. Abies religiosa Seedling limitations for passive restoration practices at the monarch butterfly biosphere reserve in Mexico. Front. Ecol. 2020, 13. [Google Scholar] [CrossRef]
- Romahn-Hernández, L.F.; Rodríguez-Trejo, D.A.; Villanueva-Morales, A.; Monterroso-Rivas, A.I.; Pérez-Hernández, M.J. Altitudinal range: Forest vigor factor and determinant in the natural regeneration of the sacred fir. Entreciencias 2020, 8, 1–17. [Google Scholar] [CrossRef]
- MacLauchlan, M.; Buxton, K. Overview of Forest Health Conditions in Southern British Columbia; Ministry of Forests, Lands and Natural Resource Operations and Rural Development: Kamloops, BC, Canada, 2018.




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sáenz-Romero, C.; O'Neill, G.; Aitken, S.N.; Lindig-Cisneros, R. Assisted Migration Field Tests in Canada and Mexico: Lessons, Limitations, and Challenges. Forests 2021, 12, 9. https://doi.org/10.3390/f12010009
Sáenz-Romero C, O'Neill G, Aitken SN, Lindig-Cisneros R. Assisted Migration Field Tests in Canada and Mexico: Lessons, Limitations, and Challenges. Forests. 2021; 12(1):9. https://doi.org/10.3390/f12010009
Chicago/Turabian StyleSáenz-Romero, Cuauhtémoc, Greg O'Neill, Sally N. Aitken, and Roberto Lindig-Cisneros. 2021. "Assisted Migration Field Tests in Canada and Mexico: Lessons, Limitations, and Challenges" Forests 12, no. 1: 9. https://doi.org/10.3390/f12010009
APA StyleSáenz-Romero, C., O'Neill, G., Aitken, S. N., & Lindig-Cisneros, R. (2021). Assisted Migration Field Tests in Canada and Mexico: Lessons, Limitations, and Challenges. Forests, 12(1), 9. https://doi.org/10.3390/f12010009