Projecting the Impact of Climate Change on the Spatial Distribution of Six Subalpine Tree Species in South Korea Using a Multi-Model Ensemble Approach




Abstract
1. Introduction
2. Materials and Methods
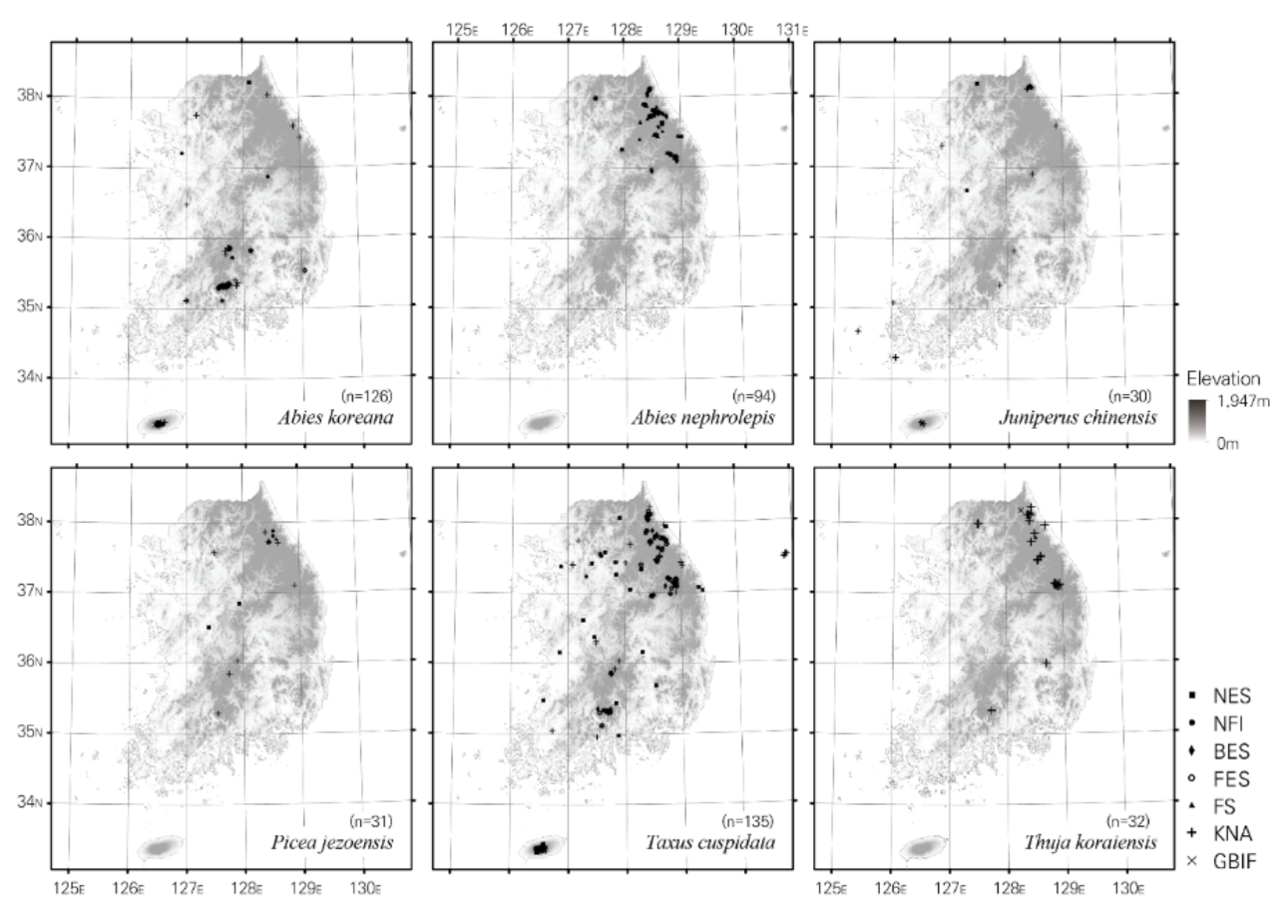
2.1. Location Data for 6 Subalpine Forest Species in South Korea
2.2. Environmental Parameters
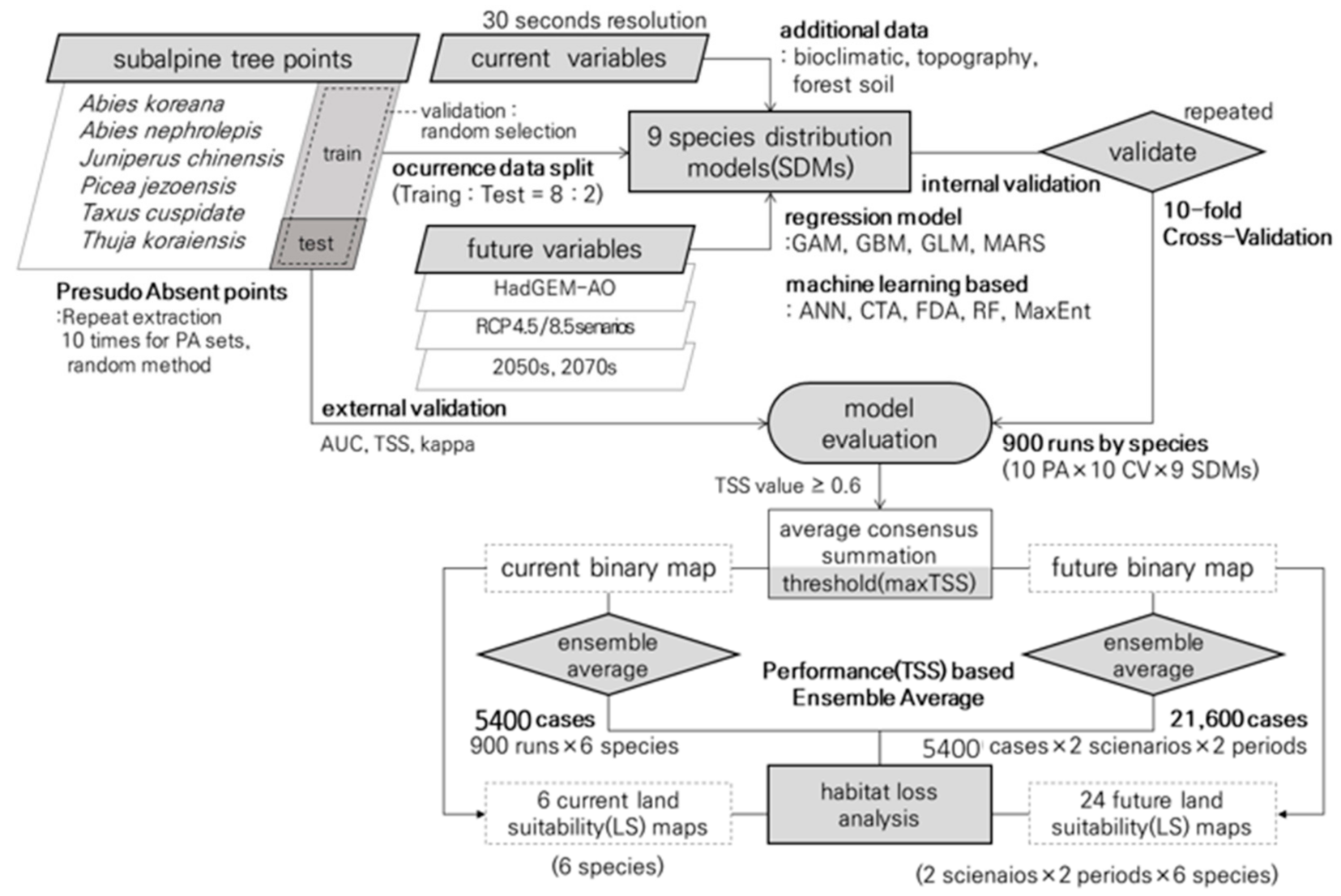
2.3. Species Distribution Ensemble Modeling and Land Suitability Analysis
3. Results
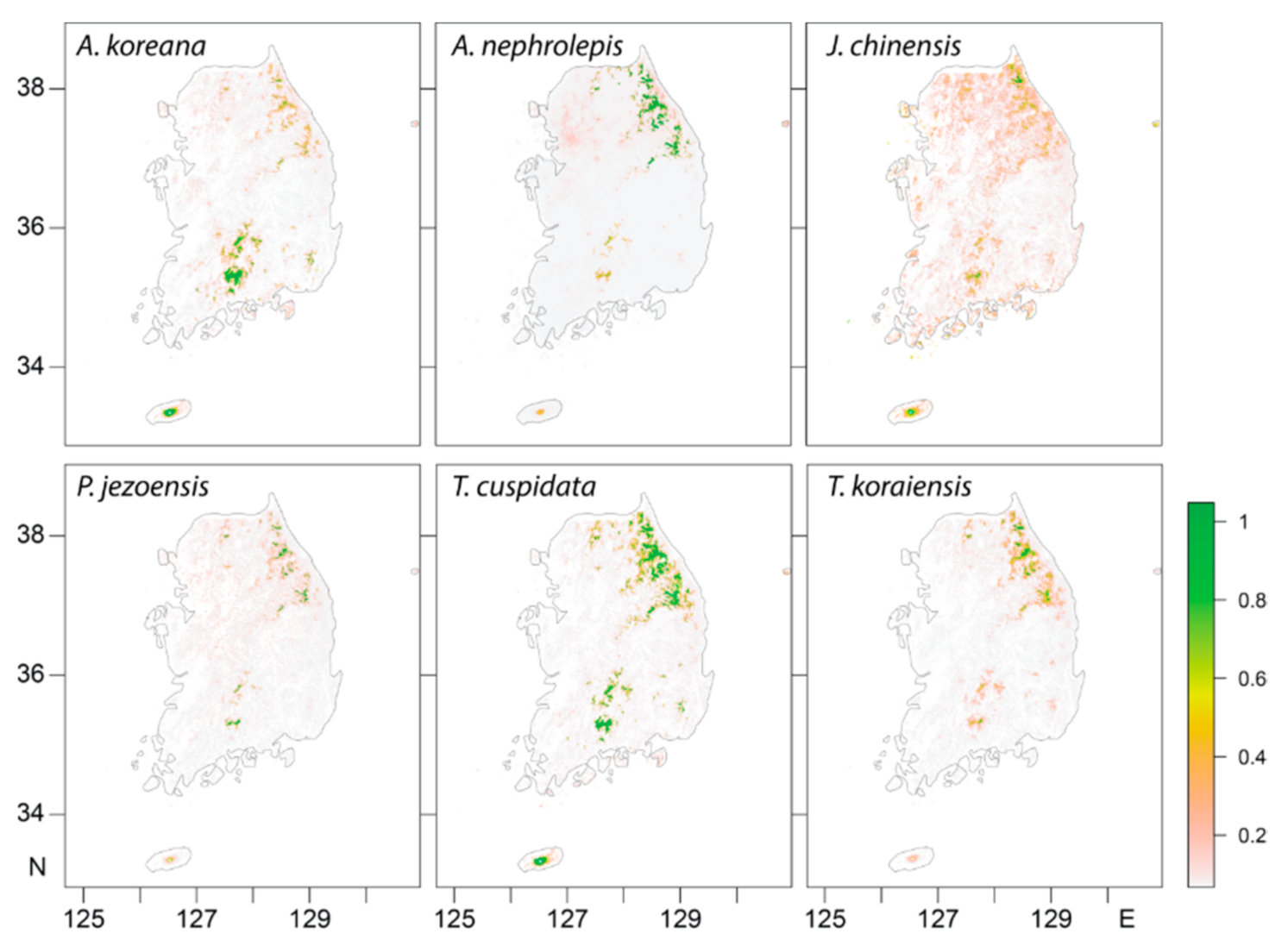
3.1. Model Accuracy and Current Potential Suitable Habitat Areas
3.2. Future Changes in the Distribution of Suitable Habitat Area and Habitat Loss
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grabherr, G.; Gottfried, M.; Pauli, H. Climate change impacts in alpine environments. Geogr. Compass 2010, 4, 1133–1153. [Google Scholar] [CrossRef]
- Kayama, M.; Makoto, K.; Nomura, M.; Sasa, K.; Koike, T. Growth characteristics of Sakhalin spruce (Picea glehnii) planted on the northern Japanese hillsides exposed to strong winds. Trees 2009, 23, 145–157. [Google Scholar] [CrossRef]
- Guisan, A.; Theurillat, J.-P. Assessing alpine plant vulnerability to climate change: A modeling perspective. Integr. Assess. 2000, 1, 307–320. [Google Scholar] [CrossRef]
- Bailey, R.G. Ecosystem Geography: From Ecoregions to Sites; Springer Science & Business Media: New York, NY, USA, 2009; ISBN 0387895159. [Google Scholar]
- Freeman, B.G.; Lee-Yaw, J.A.; Sunday, J.M.; Hargreaves, A.L. Expanding, shifting and shrinking: The impact of global warming on species’ elevational distributions. Glob. Ecol. Biogeogr. 2018, 27, 1268–1276. [Google Scholar] [CrossRef]
- Korea National Park Research Institute (KNPRI). National Park Climate Change Ecosystem Monitoring; KNPRI: Wonju, Korea, 2019; pp. 105–222. [Google Scholar]
- International Union for Conservation of Nature and Natural Resources RED LIST. Available online: https://www.iucnredlist.org (accessed on 10 January 2020).
- National Institute of Forest Science (NIFS). Guidelines for Survey of Coniferous Tree Species in Endangered Alpine Areas; NIFS: Seoul, Korea, 2017.
- Ministry of Environment (MOE). Subalpine Coniferous Forest Management Measures According to Climate Change; MOE: Sejong, Korea, 2016.
- Pecchi, M.; Marchi, M.; Moriondo, M.; Forzieri, G.; Ammoniaci, M.; Bernetti, I.; Bindi, M.; Chirici, G. Potential impact of climate change on the spatial distribution of key forest tree species in Italy under RCP 4.5 for 2050s. Forests 2020, 11, 934. [Google Scholar] [CrossRef]
- Breiner, F.T.; Guisan, A.; Bergamini, A.; Nobis, M.P. Overcoming limitations of modelling rare species by using ensembles of small models. Methods Ecol. Evol. 2015, 6, 1210–1218. [Google Scholar] [CrossRef]
- Meller, L.; Cabeza, M.; Pironon, S.; Barbet-Massin, M.; Maiorano, L.; Georges, D.; Thuiller, W. Ensemble distribution models in conservation prioritization: From consensus predictions to consensus reserve networks. Divers. Distrib. 2014, 20, 309–321. [Google Scholar] [CrossRef]
- Goberville, E.; Beaugrand, G.; Hautekèete, N.C.; Piquot, Y.; Luczak, C. Uncertainties in the projection of species distributions related to general circulation models. Ecol. Evol. 2015, 5, 1100–1116. [Google Scholar] [CrossRef]
- National Institute of Ecology ECObank. Available online: https://www.nie-ecobank.kr (accessed on 12 January 2020).
- Park, H. Development and Application of Climate Change Sensitivity Assessment Method for Plants Using the Species Distribution Models. Ph.D. Thesis, Kangwon National University, Chuncheon, Korea, August 2016. [Google Scholar]
- National Arboretum Korea Biodiversity Information System. Available online: http://www.nature.go.kr (accessed on 2 February 2020).
- Global Biodiversity Information Facility. Available online: https://www.gbif.org (accessed on 25 January 2020).
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A.; Elith, J.; Dudík, M.; Ferrier, S.; Huettmann, F.; et al. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Mateo, R.G.; Felicísimo, Á.M.; Muñoz, J. Effects of the number of presences on reliability and stability of MARS species distribution models: The importance of regional niche variation and ecological heterogeneity. J. Veg. Sci. 2010, 21, 908–922. [Google Scholar] [CrossRef]
- Ter Steege, H.; Haripersaud, P.P.; Banki, O.S.; Schieving, F. A Model of botanical collectors’ behavior in the field: Never the same species twice. Am. J. Bot. 2011, 98, 31–37. [Google Scholar] [CrossRef] [PubMed]
- McPherson, J.M.; Jetz, W.; Rogers, D.J. The effects of species’ range sizes on the accuracy of distribution models: Ecological phenomenon or statistical artefact? J. Appl. Ecol. 2004, 41, 811–823. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M.; et al. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Schreuder, M.; De La Cruz, J.; Guarino, L. Using GIS to check co-ordinates of genebank accessions. Genet. Resour. Crop Evol. 1999, 46, 291–296. [Google Scholar] [CrossRef]
- WorldClim. Available online: https://worldclim.org (accessed on 20 January 2020).
- Fick, S.; Hijmans, R. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017. [Google Scholar] [CrossRef]
- Busby, J. BIOCLIM-a bioclimate analysis and prediction system. Plant Prot. Q. 1991, 6, 8–9. [Google Scholar]
- Choi, J.; Lee, S. Climate Change Impact Assessment of Abies nephrolepis (Trautv.) Maxim. in Subalpine Ecosystem using Ensemble Habitat Suitability Modeling. J. Korea Soc. Environ. Restor. Technol. 2018, 21, 103–118. [Google Scholar] [CrossRef]
- Wright, J.S.; Sobel, A.; Galewsky, J. Diagnosis of zonal mean relative humidity changes in a warmer climate. J. Clim. 2010, 23, 4556–4569. [Google Scholar] [CrossRef]
- Wolter, P.T.; Mladenoff, D.J.; Host, G.E.; Crow, T.R. Using Multi-Temporal Landsat Imagery. Photogramm. Eng. Remote Sens. 1995, 61, 1129–1143. [Google Scholar]
- Han, N.; Cha, D.; Kwon, J.; Cho, H. A Study on Geotop Classification and Geodiversity in Mt. Jeombong Experimental Forest. J. Korea Soc. Environ. Restor. Technol. 2015, 18, 179–190. [Google Scholar]
- Gorokhovich, Y.; Voustianiouk, A. Accuracy assessment of the processed SRTM-based elevation data by CGIAR using field data from USA and Thailand and its relation to the terrain characteristics. Remote Sens. Environ. 2006, 104, 409–415. [Google Scholar] [CrossRef]
- Wilson, J.P.; Gallant, J.C. Terrain Analysis: Principles and Applications; Wiley: Hoboken, NJ, USA, 2000. [Google Scholar]
- SRTM Data. Available online: http://srtm.csi.cgiar.org/srtmdata (accessed on 20 February 2020).
- Forest Spatial Information Service. Available online: http://map.forest.go.kr (accessed on 20 January 2020).
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Tiago, P.; Pereira, H.M.; Capinha, C. Using citizen science data to estimate climatic niches and species distributions. Basic Appl. Ecol. 2017, 20, 75–85. [Google Scholar] [CrossRef]
- Lobo, J.M.; Tognelli, M.F. Exploring the effects of quantity and location of pseudo-absences and sampling biases on the performance of distribution models with limited point occurrence data. J. Nat. Conserv. 2011, 19, 1–7. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Phillips, S.; Leathwick, J.; Elith, J. Dismo: Species Distribution Modeling, R Package Version 1.1-4; 2020. Available online: http://CRAN.R-project.org/package=dismo (accessed on 20 October 2020).
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F. Biomod2: Ensemble Platform for Species Distribution Modeling; R Package Version 3.3-7.1; 2019. Available online: http://CRAN.R-project.org/package=biomod2 (accessed on 20 February 2020).
- Leroy, B.; Meynard, C.N.; Bellard, C.; Courchamp, F. virtualspecies, an R package to generate virtual species distributions. Ecography 2016, 39, 599–607. [Google Scholar] [CrossRef]
- Robinson, H.S.; Weckworth, B. Landscape Ecology: Linking Landscape Metrics to Ecological Processes. In Snow Leopards; McCarthy, T., Mallon, D.B.T.-S.L., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 395–405. [Google Scholar]
- Turner, M.G.; Gardner, R.H. Introduction to Landscape Ecology and Scale BT. In Landscape Ecology in Theory and Practice: Pattern and Process; Turner, M.G., Gardner, R.H., Eds.; Springer: New York, NY, USA, 2015; pp. 1–32. [Google Scholar]
- Connor, T.; Viña, A.; Winkler, J.A.; Hull, V.; Tang, Y.; Shortridge, A.; Yang, H.; Zhao, Z.; Wang, F.; Zhang, J.; et al. Interactive spatial scale effects on species distribution modeling: The case of the giant panda. Sci. Rep. 2019, 9, 14563. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of ocurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Liu, C.; White, M.; Newell, G. Selecting thresholds for the prediction of species occurrence with presence-only data. J. Biogeogr. 2013, 40, 778–789. [Google Scholar] [CrossRef]
- Lee, W.T.; Yim, Y.J. The Phytogeography; Kangwon National University Press: Chuncheon, Korea, 2002. [Google Scholar]
- Lee, D.; Jin, D.; Lee, J.; Choi, I.; Choi, B. Floristic study of Mt. Namdeogyu. Korean J. Plant Taxon. 2013, 43, 69–79. [Google Scholar] [CrossRef]
- National Institute of Forest Science (NIFS). Endangered Alpine Coniferous Species in Korea; NIFS: Seoul, Korea, 2019.
- Kim, E.; Lee, J.; Park, G.; Lim, J. Change of Subalpine Coniferous Forest Area over the Last 20 Years. J. Korean Soc. For. Sci. 2019, 108, 10–20. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | NES 1 | BES 2 | KNA 3 | NFI 4 | FES 5 | GBIF 6 | FS 7 | Total |
|---|---|---|---|---|---|---|---|---|
| Abies koreana | 9 (11) | 30 (30) | 33 (37) | 10 (15) | 40 (45) | 4 (7) | - | 126 (145) |
| Abies nephrolepis | 9 (9) | 27 (45) | - | 7 (10) | 14 (37) | 0 (1) | 37 (239) | 94 (341) |
| Juniperus chinensis | 3 (7) | 1 (1) | 12 (16) | - | 12 (41) | 2 (4) | - | 30 (69) |
| Picea jezoensis | 3 (3) | - | 10 (16) | 2 (4) | - | - | 16 (16) | 31 (39) |
| Taxus cuspidata | 36 (74) | 19 (25) | 41 (67) | 12 (50) | 24 (57) | 3 (14) | - | 135 (287) |
| Thuja koraiensis | - | - | 24 (28) | 1 (1) | - | 6 (19) | 1 (1) | 32 (49) |
| Total | 60 (104) | 77 (101) | 120 (164) | 32 (80) | 90 (180) | 15 (45) | 54 (256) | 448 (930) |
| Code | Description | Unit | Contribution (%) |
|---|---|---|---|
| Bio01 | Annual mean temperature | °C | 9.7 ± 5.4 |
| Bio02 | Mean diurnal temperature range | °C | 11.0 ± 5.8 |
| Bio04 | Temperature seasonality | °C | 5.5 ± 2.0 |
| Bio12 | Annual precipitation | mm | 8.5 ± 4.4 |
| Bio13 | Precipitation of wettest month | mm | 7.5 ± 3.7 |
| Bio14 | Precipitation of driest month | mm | 5.2 ± 3.2 |
| DEM | Elevation | m | 54.2 ± 0.1 |
| LF | Landforms index from TPI * | - | 4.9 ± 1.5 |
| SP | Slope position index from TPI * | - | 6.7 ± 1.8 |
| SLP | Slope | ° | 5.3 ± 0.7 |
| ASP | Cos(Aspect(rad)) | - | 5.1 ± 1.3 |
| FSD | Forest soil depth | cm | 6.7 ± 3.7 |
| FST | Forest soil texture | - | 5.4 ± 2.3 |
| FSM | Forest soil moisture | % | 4.6 ± 2.1 |
| Species | Models | AUC (0–1) | kappa (0–1) | TSS (0–1) |
|---|---|---|---|---|
| All species | ANN 1 | 0.903 ± 0.062 | 0.738 ± 0.110 | 0.783 ± 0.112 |
| CTA 2 | 0.864 ± 0.065 | 0.635 ± 0.117 | 0.733 ± 0.117 | |
| FDA 3 | 0.895 ± 0.070 | 0.783 ± 0.103 | 0.779 ± 0.117 | |
| GAM 4 | 0.847 ± 0.074 | 0.643 ± 0.120 | 0.679 ± 0.141 | |
| GBM 5 | 0.929 ± 0.054 | 0.807 ± 0.100 | 0.830 ± 0.096 | |
| GLM 6 | 0.881 ± 0.071 | 0.673 ± 0.117 | 0.737 ± 0.137 | |
| MARS 7 | 0.881 ± 0.070 | 0.705 ± 0.115 | 0.742 ± 0.137 | |
| MAXENT 8 | 0.863 ± 0.081 | 0.706 ± 0.134 | 0.719 ± 0.156 | |
| RF 9 | 0.926 ± 0.051 | 0.808 ± 0.094 | 0.828 ± 0.092 | |
| Abies koreana | All models | 0.947 ± 0.035 | 0.858 ± 0.005 | 0.860 ± 0.064 |
| Abies nephrolepis | 0.946 ± 0.037 | 0.846 ± 0.072 | 0.879 ± 0.073 | |
| Juniperus chinensis | 0.761 ± 0.122 | 0.476 ± 0.189 | 0.539 ± 0.208 | |
| Picea jezoensis | 0.906 ± 0.077 | 0.767 ± 0.122 | 0.811 ± 0.144 | |
| Taxus cuspidata | 0.860 ± 0.050 | 0.681 ± 0.087 | 0.668 ± 0.091 | |
| Thuja koraiensis | 0.905 ± 0.078 | 0.703 ± 0.143 | 0.796 ± 0.155 |
| Species | Elevation of Habitat (Mean ± SD, m) | TSS-Weighted Average | Equal Value Average | CA-Based Average | |||
|---|---|---|---|---|---|---|---|
| Area (km2) | Fraction of Country (%) | Area (km2) | Fraction of Country (%) | Area (km2) | Fraction of Country (%) | ||
| Abies koreana | 1382 ± 352 | 1820.1 | 1.82 | 1823.0 | 1.82 | 2852.1 | 2.85 |
| Abies nephrolepis | 1297 ± 186 | 3021.7 | 3.01 | 1795.8 | 1.79 | 3021.7 | 3.02 |
| Juniperus chinensis | 1331 ± 340 | 307.4 | 0.31 | 334.3 | 0.33 | 649.7 | 0.65 |
| Picea jezoensis | 1338 ± 214 | 448.6 | 0.45 | 497.4 | 0.50 | 618.3 | 0.62 |
| Taxus cuspidata | 1092 ± 448 | 674.4 | 0.67 | 674.4 | 0.67 | 1120.2 | 1.12 |
| Thuja koraiensis | 1240 ± 263 | 1129.8 | 1.12 | 1065.7 | 1.06 | 1129.8 | 1.13 |
| Species | Areal Loss in 2050s (2041–2060) (%) | Areal Loss in 2070s (2061–2080) (%) | ||||
|---|---|---|---|---|---|---|
| RCP 4.5 | RCP 8.5 | Δ | RCP 4.5 | RCP 8.5 | Δ | |
| Abies koreana | 23.9 ± 24.4 | 28.4 ± 27.8 | 4.5 | 36.5 ± 25.4 | 36.7 ± 35.6 | 0.2 |
| Abies nephrolepis | 56.3 ± 33.2 | 57.7 ± 35.5 | 1.4 | 65.2 ± 34.7 | 62.2 ± 39.8 | −3.0 |
| Juniperus chinensis | 37.2 ± 26.0 | 39.0 ± 29.5 | 1.8 | 43.9 ± 26.9 | 40.6 ± 33.2 | −3.3 |
| Picea jezoensis | 22.1 ± 25.6 | 21.6 ± 26.5 | −0.5 | 25.9 ± 30.3 | 23.7 ± 33.2 | −2.2 |
| Taxus cuspidata | 13.2 ± 16.6 | 18.0 ± 22.5 | 4.8 | 17.7 ± 20.1 | 26.7 ± 32.2 | 9.0 |
| Thuja koraiensis | 59.5 ± 30.6 | 65.9 ± 34.2 | 6.4 | 62.7 ± 33.8 | 67.1 ± 39.1 | 4.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Jung, H.; Choi, J. Projecting the Impact of Climate Change on the Spatial Distribution of Six Subalpine Tree Species in South Korea Using a Multi-Model Ensemble Approach. Forests 2021, 12, 37. https://doi.org/10.3390/f12010037
Lee S, Jung H, Choi J. Projecting the Impact of Climate Change on the Spatial Distribution of Six Subalpine Tree Species in South Korea Using a Multi-Model Ensemble Approach. Forests. 2021; 12(1):37. https://doi.org/10.3390/f12010037
Chicago/Turabian StyleLee, Sanghyuk, Huicheul Jung, and Jaeyong Choi. 2021. "Projecting the Impact of Climate Change on the Spatial Distribution of Six Subalpine Tree Species in South Korea Using a Multi-Model Ensemble Approach" Forests 12, no. 1: 37. https://doi.org/10.3390/f12010037
APA StyleLee, S., Jung, H., & Choi, J. (2021). Projecting the Impact of Climate Change on the Spatial Distribution of Six Subalpine Tree Species in South Korea Using a Multi-Model Ensemble Approach. Forests, 12(1), 37. https://doi.org/10.3390/f12010037