Evaluating Soil–Root Interaction of Hybrid Larch Seedlings Planted under Soil Compaction and Nitrogen Loading

 ,
, 
 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Site and Experimental Design
2.2. Soil Environment
2.3. Seedling Performances
2.3.1. Aboveground
2.3.2. Belowground
2.4. Data Analysis
3. Results
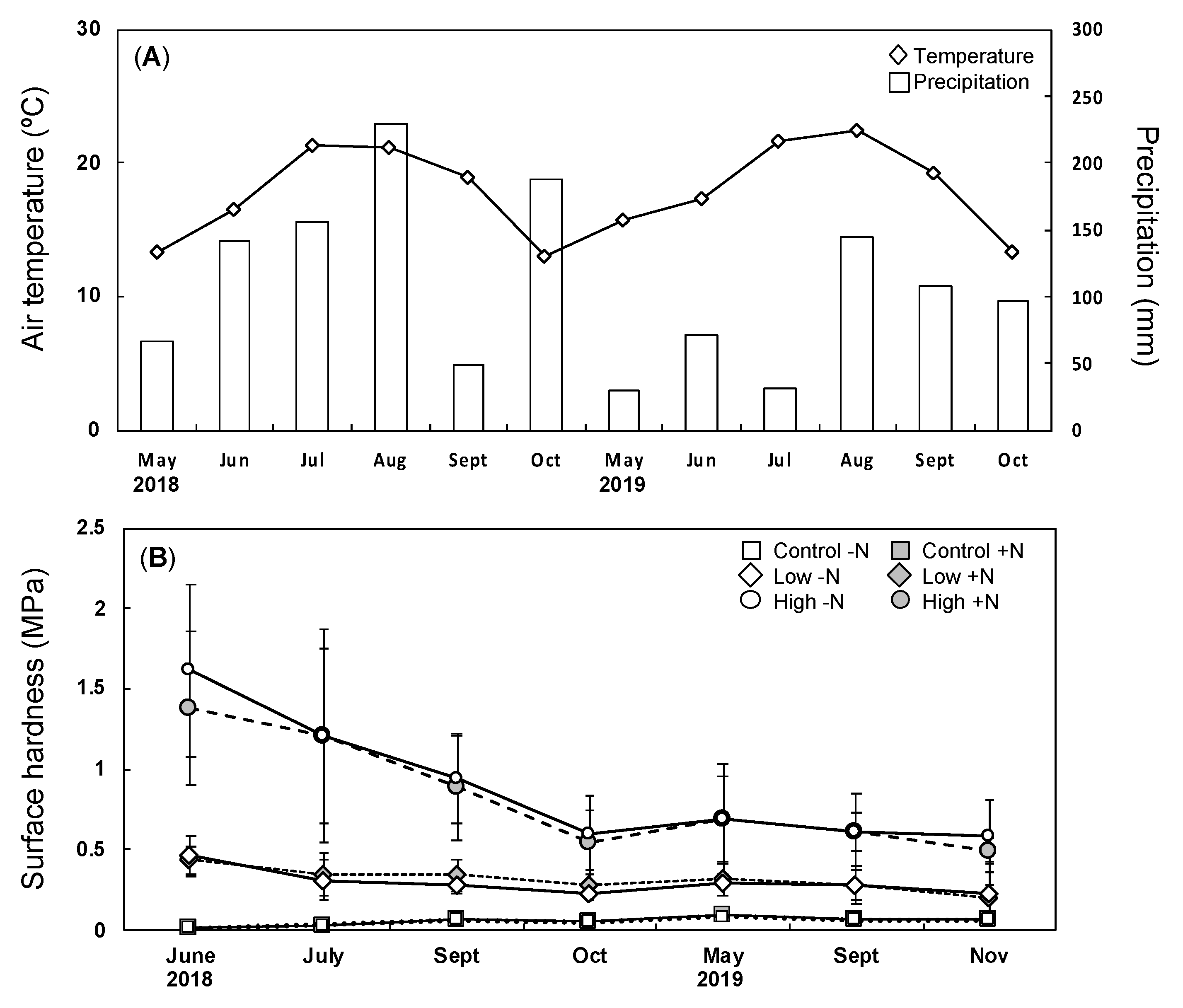
3.1. Environment at the Study Site
3.2. Aboveground Responses
3.3. Belowground Responses
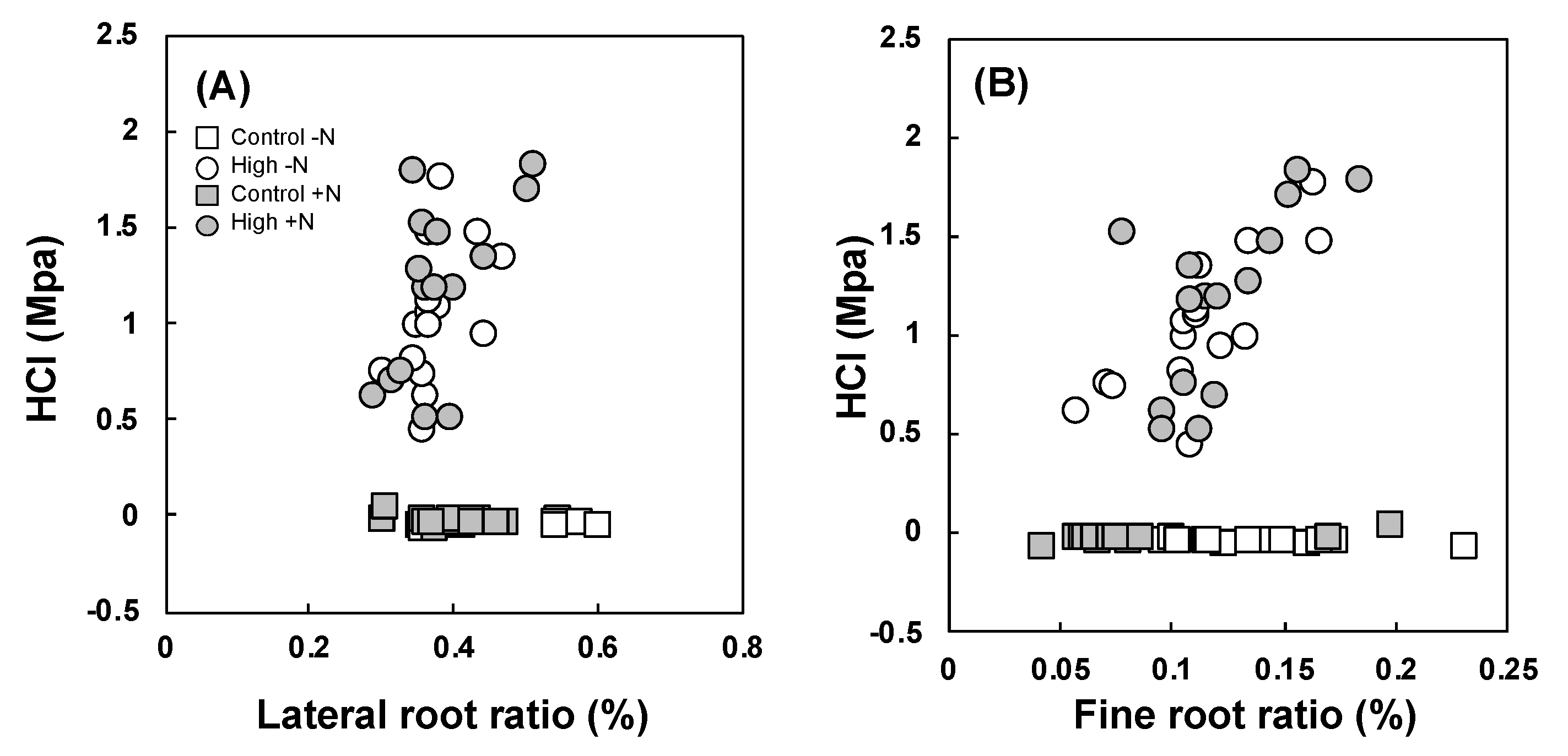
3.4. Relationship between Soil Hardness and Roots
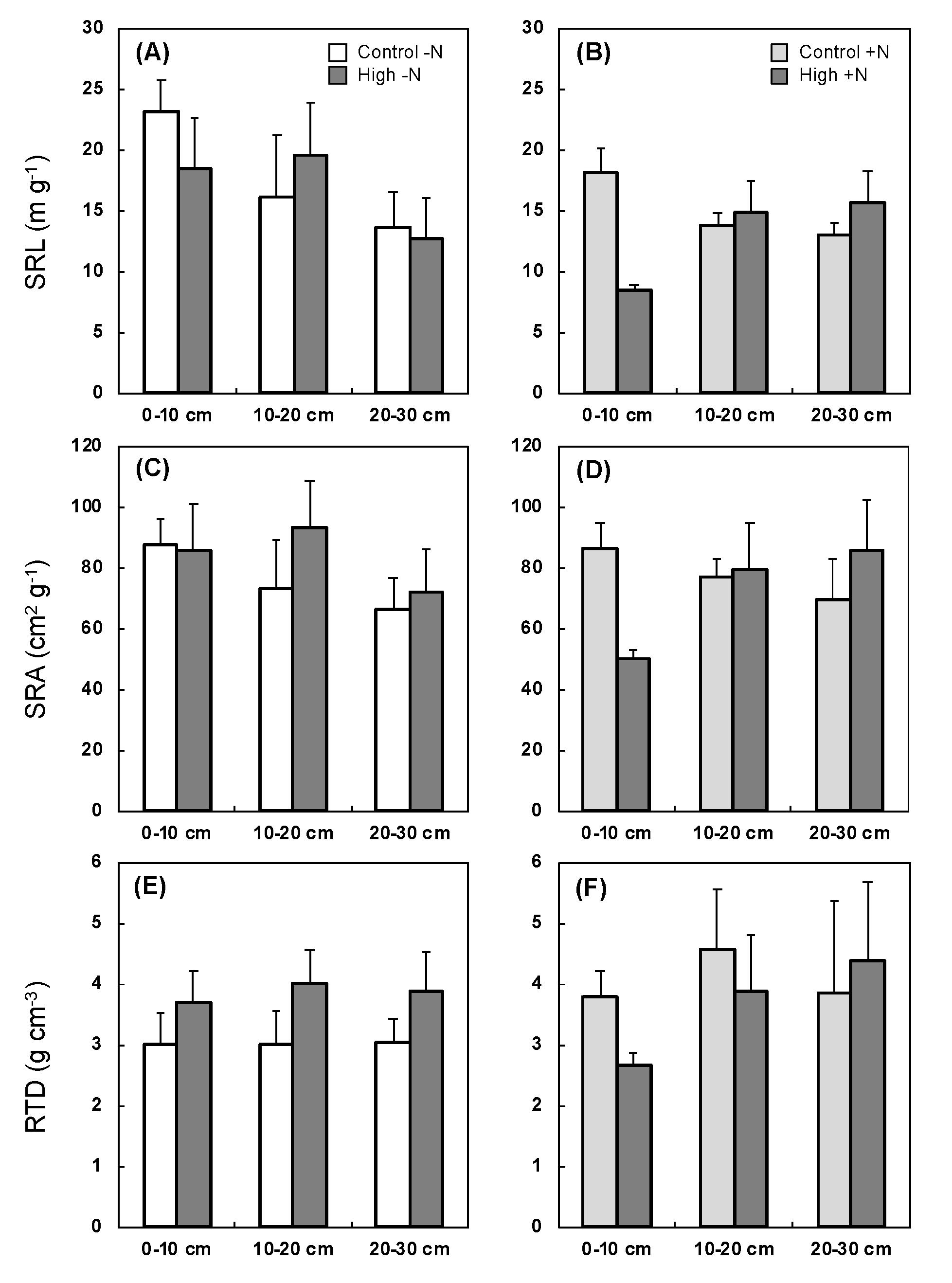
3.5. Responses of Root Morphological Traits and ECM Association
4. Discussion
4.1. Relationship between Roots and Soil
4.2. Root Morphological Developments and ECM Symbiosis
4.3. Aboveground Responses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cambi, M.; Certini, G.; Neri, F.; Marchi, E. The impact of heavy traffic on forest soils: A review. For. Ecol. Manag. 2015, 338, 124–138. [Google Scholar] [CrossRef]
- Marchi, E.; Chung, W.; Visser, R.; Abbas, D.; Nordfjell, T.; Mederski, P.S.; McEwan, A.; Brink, M.; Laschi, A. Sustainable Forest Operations (SFO): A new paradigm in a changing world and climate. Sci. Total. Environ. 2018, 634, 1385–1397. [Google Scholar] [CrossRef]
- Mariotti, B.; Hoshika, Y.; Cambi, M.; Marra, E.; Feng, Z.Z.; Paoletti, E.; Marchi, E. Vehicle-induced compaction of forest soil affects plant morphological and physiological attributes: A meta-analysis. For. Ecol. Manag. 2020, 462. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Soil compaction and growth of woody plants. Scand. J. For. Res. 1999, 14, 596–619. [Google Scholar] [CrossRef]
- Tan, X.; Chang, S.X. Soil compaction and forest litter amendment affect carbon and net nitrogen mineralization in a boreal forest soil. Soil Tillage Res. 2007, 93, 77–86. [Google Scholar] [CrossRef]
- Batey, T. Soil compaction and soil management–a review. Soil Use Manag. 2009, 25, 335–345. [Google Scholar] [CrossRef]
- Kara, Ö.; Bolat, İ. Influence of soil compaction on microfungal community structure in two soil types in Bartin Province, Turkey. J. Basic. Microbiol. 2007, 47, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Moehring, D.M.; Rawls, I.W. Detrimental effect of wet weather logging. J. For. 1970, 68, 166–167. [Google Scholar] [CrossRef]
- Ampoorter, E.; De Frenne, P.; Hermy, M.; Verheyen, K. Effects of soil compaction on growth and survival of tree saplings: A meta-analysis. Basic Appl. Ecol. 2011, 12, 394–402. [Google Scholar] [CrossRef]
- Kormanek, M.; Banach, J.; Leńczuk, D. Influence of soil compaction on the growth of silver fir (Abies alba Mill.) under a forest canopy. Ecol. Quest. 2015, 22, 47–54. [Google Scholar] [CrossRef]
- Passioura, J.B. Soil conditions and plant growth. Plant Cell Environ. 2002, 25, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Noguchi, M.; Akibayashi, Y.; Noda, M.; Kadomatsu, M.; Sasa, K. Twenty years of community dynamics in a mixed conifer - broad-leaved forest under a selection system in northern Japan. Can. J. For. Res. 2006, 36, 1363–1375. [Google Scholar] [CrossRef]
- Yamazaki, H.; Yoshida, T. Significance and limitation of scarification treatments on early establishment of Betula maximowicziana, a tree species producing buried seeds: Effects of surface soil retention. J. For. Res. 2018, 23, 166–172. [Google Scholar] [CrossRef]
- Fleming, R.L.; Powers, R.F.; Foster, N.W.; Kranabetter, J.M.; Scott, D.A.; Ponder, F., Jr.; Berch, S.; Chapman, W.K.; Kabzems, R.D.; Ludovici, K.H.; et al. Effects of organic matter removal, soil compaction, and vegetation control on 5-year seedling performance: A regional comparison of Long-Term Soil Productivity sites. Can. J. For. Res. 2006, 36, 529–550. [Google Scholar] [CrossRef]
- Ponder, F., Jr.; Fleming, R.L.; Berch, S.; Busse, M.D.; Elioff, J.D.; Hazlett, P.W.; Kabzems, R.D.; Kranabetter, J.M.; Morris, D.M.; Page-Dumroese, D.; et al. Effects of organic matter removal, soil compaction and vegetation control on 10th year biomass and foliar nutrition: LTSP continent-wide comparisons. For. Ecol. Manag. 2012, 278, 35–54. [Google Scholar] [CrossRef]
- Alameda, D.; Villar, R. Moderate soil compaction: Implications on growth and architecture in seedlings of 17 woody plant species. Soil Tillage Res. 2009, 103, 325–331. [Google Scholar] [CrossRef]
- Gebauer, R.; Volarik, D.; Martinkova, M. Impact of soil pressure and compaction on tracheids in Norway spruce seedlings. New For. 2011, 41, 75–88. [Google Scholar] [CrossRef]
- Jourgholami, M.; Khoramizadeh, A.; Zenner, E.K. Effects of soil compaction on seedling morphology, growth, and architecture of chestnut-leaved oak (Quercus castaneifolia). iForest 2016, 10, 145–153. [Google Scholar] [CrossRef]
- Cambi, M.; Mariotti, B.; Fabiano, F.; Maltoni, A.; Tani, A.; Foderi, C.; Laschi, A.; Marchi, E. Early response of Quercus robur seedlings to soil compaction following germination. Land Degrad. Dev. 2018, 29, 916–925. [Google Scholar] [CrossRef]
- Shah, A.N.; Tanveer, M.; Shahzad, B.; Yang, G.; Fahad, S.; Ali, S.; Bukhari, M.A.; Tung, S.A.; Hafeez, A.; Souliyanonh, B. Soil compaction effects on soil health and crop productivity. Environ. Sci. Pollut. Res. 2017, 24, 10056–10067. [Google Scholar] [CrossRef]
- Reisinger, T.W.; Pope, P.E.; Hammond, S.C. Natural recovery of compacted soils in an upland hardwood forest in Indiana. North. J. Appl. For. 1992, 9, 138–141. [Google Scholar] [CrossRef]
- Bardgett, R. Organism interactions and soil processes. In The Biology of Soil: A Community and Ecosystem Approach; Crawley, M.J., Little, C., Southwood, T.R.E., Ulfstrand, S., Eds.; Oxford University Press: Oxford, UK, 2005; pp. 57–85. [Google Scholar]
- Zenner, E.K.; Fauskee, J.T.; Berger, A.L.; Puettmann, K.I. Impacts of skidding traffic intensity on soil disturbance, soil recovery, and aspen regeneration in north central Minnesota. North. J. Appl. For. 2007, 24, 177–183. [Google Scholar] [CrossRef]
- Meyer, C.; Lüscher, P.; Schulin, R. Recovery of forest soil from compaction in skid tracks planted with black alder (Alnus glutinosa (L.) Gaertn.). Soil. Tillage Res. 2014, 143, 7–16. [Google Scholar] [CrossRef]
- Fernandez, J.L.F.; Hartmann, P.; von Wilpert, K. Planting of alder trees at the edge of skid trails helps to stabilize forest topsoil structure against damage caused by heavy forestry machines. Soil Tillage Res. 2019, 187, 214–218. [Google Scholar] [CrossRef]
- Rodríguez-González, P.M.; Campelo, F.; Albuquerque, A.; Rivaes, R.; Ferreira, T.; Pereira, J.S. Sensitivity of black alder (Alnus glutinosa [L.] Gaertn.) growth to hydrological changes in wetland forests at the rear edge of the species distribution. Plant Ecol. 2014, 215, 233–245. [Google Scholar] [CrossRef]
- Schwarz, M.; Lehmann, P.; Or, D. Quantifying lateral root reinforcement in steep slopes - from a bundle of roots to tree stands. Earth Surf. Process. Landf. 2010, 35, 354–367. [Google Scholar] [CrossRef]
- Sinnett, D.; Morgan, G.; Williams, M.; Hutchings, T.R. Soil penetration resistance and tree root development. Soil Use Manag. 2008, 24, 273–280. [Google Scholar] [CrossRef]
- Correa, J.; Postma, J.A.; Watt, M.; Wojciechowski, T. Soil compaction and the architectural plasticity of root systems. J. Exp. Bot. 2019, 70, 6019–6034. [Google Scholar] [CrossRef]
- Aust, W.M.; Burger, J.A.; McKee, W.H., Jr.; Scheerer, G.A.; Tippett, M.D. Bedding and fertilization ameliorate effects of designated wet-weather skid trails after four years for loblolly pine (Pinus taeda) plantations. South. J. Appl. For. 1998, 22, 222–226. [Google Scholar] [CrossRef]
- Simcock, R.C.; Parfitt, R.L.; Skinner, M.F.; Dando, J.; Graham, J.D. The effects of soil compaction and fertilizer application on the establishment and growth of Pinus radiata. Can. J. For. Res. 2006, 36, 1077–1086. [Google Scholar] [CrossRef]
- Jordan, D.; Ponder, F.; Hubbard, V.C. Effects of soil compaction, forest leaf litter and nitrogen fertilizer on two oak species and microbial activity. Appl. Soil. Ecol. 2003, 23, 33–41. [Google Scholar] [CrossRef]
- Picchio, R.; Tavankar, F.; Nikooy, M.; Pignatti, G.; Venanzi, R.; Lo Monaco, A. Morphology, growth and architecture responses of beech (Fagus orientalis Lipsky) and maple tree (Acer velutinum Boiss.) seedlings to soil compaction stress caused by mechanized logging operations. Forests 2019, 10, 771. [Google Scholar] [CrossRef]
- Ryu, K.; Watanabe, M.; Shibata, H.; Takagi, K.; Nomura, M.; Koike, T. Ecophysiological responses of the larch species in northern Japan to environmental changes as a basis for afforestation. Landsc. Ecol. Eng. 2009, 5, 99–106. [Google Scholar] [CrossRef]
- Abaimov, A. Geographical distribution and genetics of Siberian larch species. In Permafrost Ecosystems; Osawa, A., Zyryanova, O., Matsuura, Y., Kajimoto, T., Wein, R., Eds.; Springer Ecol. Studies 209: Dordrecht, The Netherlands, 2010; pp. 41–58. [Google Scholar]
- Kita, K.; Fujimoto, T.; Uchiyama, K.; Kuromaru, M.; Akutsu, H. Estimated amount of carbon accumulation of hybrid larch in three 31-year-old progeny test plantations. J. Wood. Sci. 2009, 55, 425–434. [Google Scholar] [CrossRef]
- Ministry of Agriculture, Forestry and Fisheries (2017) Annual Report on Forest and Forestry in Japan, Fiscal Year 2017 (Summary). Available online: https://www.maff.go.jp/e/data/publish/attach/pdf/index-95.pdf (accessed on 22 June 2020).
- Reisinger, T.W.; Simmons, G.L.; Pope, P.E. The impact of timber harvesting on soil properties and seedling growth in the south. South. J. Appl. For. 1988, 12, 58–67. [Google Scholar] [CrossRef]
- Cambi, M.; Hoshika, Y.; Mariotti, B.; Paoletti, E.; Picchio, R.; Venanzi, R.; Marchi, E. Compaction by a forest machine affects soil quality and Quercus robur L. seedling performance in an experimental field. For. Ecol. Manag. 2017, 384, 406–414. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M. Mycorrhizal types differ in ecophysiology and alter plant nutrition and soil processes. Biol. Rev. 2019, 94, 1857–1880. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Agathokleous, E.; Qu, L.; Fujita, S.; Watanabe, M.; Tamai, Y.; Mao, Q.; Koyama, A.; Koike, T. Effects of simulated nitrogen deposition on ectomycorrhizae community structure in hybrid larch and its parents grown in volcanic ash soil: The role of phosphorous. Sci. Total Environ. 2018, 618, 905–915. [Google Scholar] [CrossRef]
- Sugai, T.; Watanabe, T.; Kita, K.; Koike, T. Nitrogen loading increases the ozone sensitivity of larch seedlings with higher sensitivity to nitrogen loading. Sci. Total Environ. 2019, 663, 587–595. [Google Scholar] [CrossRef]
- Fujita, S.; Wang, X.N.; Kita, K.; Koike, T. Effects of nitrogen loading under low and high phosphorus conditions on above- and below-ground growth of hybrid larch F-1 seedlings. iForest 2018, 11, 32. [Google Scholar] [CrossRef]
- Binkley, D.; Fisher, R.F. Soil management—Harvesting, site preparation, conversion, and drainage. In Ecology and Management of Forest Soils, 5th ed.; Binkley, D., Fisher, R.F., Eds.; Temple-Inland Inc.: Hoboken, NJ, USA, 2013; pp. 213–234. [Google Scholar]
- WRB, I.W.G. World reference base for soil resources. In World Soil Resources Report No. 103; FAO: Rome, Italy, 2006. [Google Scholar]
- Research committee of Japanese Institute of Landscape Architecture. Research Committee of Japanese Institute of Landscape Architecture: Ground Maintenance Manual in Landscape Planting. J. Jpn. Inst. Landsc. Archit. 2000, 63, 224–241. [Google Scholar]
- Ministry of the Environment (2019) Annual Report on the Environment in Japan 2019. Available online: https://www.env.go.jp/en/wpaper/2019/index.html (accessed on 22 June 2020).
- Yamaguchi, M.; Otani, Y.; Li, P.; Nagao, H.; Lenggoro, I.W.; Ishida, A.; Yazaki, K.; Noguchi, K.; Nakaba, S.; Yamane, K.; et al. Effects of long-term exposure to ammonium sulfate particles on growth and gas exchange rates of Fagus crenata, Castanopsis sieboldii, Larix kaempferi and Cryptomeria japonica seedlings. Atmos. Environ. 2014, 97, 493–500. [Google Scholar] [CrossRef]
- R. Core. Team: A language and environment for statistical computing. R Found. Stat. Comput. Vienna Austria. Available online: https://www.r-project.org (accessed on 2 August 2020).
- Haynes, R.J.; Naidu, R. Influence of lime, fertilizer and manure applications on soil organic matter content and soil physical conditions: A review. Nutr. Cycl. Agroecosyst. 1998, 51, 123–137. [Google Scholar] [CrossRef]
- Gregory, A.S.; Watts, C.W.; Whalley, W.R.; Kuan, H.L.; Griffths, B.S.; Hallett, P.D.; Whitmore, A.P. Physical resilience of soil to field compaction and the interactions with plant growth and microbial community structure. Eur. J. Soil. Sci. 2007, 58, 1221–1232. [Google Scholar] [CrossRef]
- Ampoorter, E.; De Schrijver, A.; De Frenne, P.; Hermy, M.; Verheyen, K. Experimental assessment of ecological restoration options for compacted forest soils. Ecol. Eng. 2011, 37, 1734–1746. [Google Scholar] [CrossRef]
- Fernández, J.F.; Rubin, L.; Hartmann, P.; Puhlmann, H.; von Wilpert, K. Initial recovery of soil structure of a compacted forest soil can be enhanced by technical treatments and planting. For. Ecol. Manag. 2019, 431, 54–62. [Google Scholar] [CrossRef]
- Lambers, H.; Oliveira, R.S. Growth and allocation. In Plant Physiological Ecology; Lambers, H., Oliveira, R.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 385–449. [Google Scholar]
- Comas, L.H.; Callahan, H.S.; Midford, P.E. Patterns in root traits of woody species hosting arbuscular and ectomycorrhizas: Implications for the evolution of belowground strategies. Ecol. Evol. 2014, 4, 2979–2990. [Google Scholar] [CrossRef]
- Rewald, B.; Rechenmacher, A.; Godbold, D.L. It’s complicated: Intraroot system variability of respiration and morphological traits in four deciduous tree species. Plant Physiol. 2014, 166, 736–745. [Google Scholar] [CrossRef]
- Barbier, F.F.; Dun, E.A.; Kerr, S.C.; Chabikwa, T.G.; Beveridge, C.A. An Update on the Signals Controlling Shoot Branching. Trends Plant Sci. 2019, 24, 220–236. [Google Scholar] [CrossRef]
- Tracy, S.R.; Black, C.R.; Roberts, J.A.; Mooney, S.J. Soil compaction: A review of past and present techniques for investigating effects on root growth. J. Sci. Food. Agric. 2011, 91, 1528–1537. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Control | Low Compaction | High Compaction | ANOVA | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Aboveground Traits | −N | +N | −N | +N | −N | +N | C | N | C × N | |
| Stem (cm) | Current | 157.59 ± 2.91 | 164.7 ± 6.08 | 160.09 ± 3.67 | 152.92 ± 5.97 | 151.25 ± 3.64 | 140.93 ± 8.05 | * | N.S. | N.S. |
| 1-year | 42.64 ± 1.50 | 46.2 ± 2.13 | 43.94 ± 2.28 | 45.32 ± 2.39 | 35.91 ± 3.68 | 36.82 ± 4.54 | ** | N.S. | N.S. | |
| 2-year | 20.27 ± 0.75 | 20.53 ± 1.17 | 18.80 ± 2.18 | 20.14 ± 0.48 | 20.04 ± 0.74 | 20.29 ± 0.74 | N.S. | N.S. | N.S. | |
| Diameter (mm) | Current | 16.47 ± 1.89 | 19.13 ± 2.10 | 16.96 ± 2.28 | 16.71 ± 2.35 | 15.90 ± 1.66 | 14.21 ± 2.82 | *** | N.S. | ** |
| 1-year | 18.33 ± 6.20 | 22.97 ± 2.10 | 20.60 ± 2.09 | 20.10 ± 2.15 | 18.17 ± 2.91 | 16.68 ± 4.03 | ** | N.S. | ** | |
| Collar | 30.33 ± 2.85 | 34.45 ± 2.59 | 29.96 ± 3.44 | 29.75 ± 3.33 | 26.52 ± 3.07 | 23.34 ± 5.12 | *** | N.S. | ** | |
| Total (g) | 281.54 ± 16.89 | 386.49 ± 25.86 | 275.11 ± 14.70 | 277.53 ± 20.32 | 225.02 ± 17.04 | 213.15 ± 24.24 | *** | N.S. | * | |
| Stem (g) | All | 166.56 ± 8.26 | 225.01 ± 14.25 | 171.18 ± 9.97 | 169.08 ± 10.48 | 135.86 ± 10.08 | 130.92 ± 12.14 | *** | N.S. | * |
| Current | 56.27 ± 4.50 | 79.81 ± 6.79 | 61.56 ± 4.41 | 57.67 ± 4.46 | 50.51 ± 3.62 | 45.95 ± 4.36 | ** | N.S. | * | |
| Branch (g) | All | 51.27 ± 5.11 | 73.91 ± 6.44 | 46.78 ± 2.81 | 51.16 ± 5.46 | 38.29 ± 3.19 | 33.45 ± 5.41 | *** | * | * |
| Current | 13.26 ± 2.15 | 25.52 ± 4.34 | 15.14 ± 1.70 | 14.62 ± 2.92 | 17.61 ± 1.81 | 11.61 ± 2.41 | N.S. | N.S. | * | |
| Needle (g) | All | 63.70 ± 5.54 | 87.56 ± 7.27 | 57.13 ± 3.63 | 57.28 ± 5.90 | 50.86 ± 4.90 | 48.77 ± 8.05 | *** | N.S. | N.S. |
| Current | 50.53 ± 4.48 | 72.16 ± 6.52 | 47.11 ± 3.08 | 46.17 ± 4.94 | 43.61 ± 4.26 | 40.78 ± 6.43 | ** | N.S. | N.S. | |
| Current stem | BPI (m−1) | 16.18 ± 1.88 | 18.51 ± 1.68 | 16.54 ± 1.68 | 15.70 ± 1.61 | 17.82 ± 1.80 | 10.27 ± 1.32 | N.S. | N.S. | * |
| Maximum branch | Length (cm) | 67.05 ± 10.88 | 78.31 ± 18.05 | 92.24 ± 23.31 | 90.14 ± 26.95 | 69.31 ± 11.46 | 37.13 ± 3.22 | N.S. | N.S. | N.S. |
| Diameter (mm) | 4.77 ± 0.32 | 5.01 ± 0.38 | 4.47 ± 0.20 | 4.26 ± 0.43 | 4.44 ± 0.26 | 3.64 ± 0.20 | N.S. | N.S. | N.S. | |
| Total (g) | 52.59 ± 4.64 | 74.28 ± 6.76 | 48.92 ± 2.96 | 47.89 ± 5.05 | 45.52 ± 4.51 | 41.72 ± 6.46 | ** | N.S. | N.S. | |
| Needle (g) | 2.06 ± 0.39 | 2.12 ± 0.44 | 1.80 ± 0.24 | 1.72 ± 0.31 | 1.91 ± 0.37 | 0.93 ± 0.08 | N.S. | N.S. | N.S. | |
| NPI (g m−1) | 3.00 ± 0.20 | 3.11 ± 0.32 | 2.51 ± 0.17 | 2.36 ± 0.22 | 2.61 ± 0.17 | 2.50 ± 0.09 | * | N.S. | N.S. | |
| Control | High Compaction | ANOVA | ||||||
|---|---|---|---|---|---|---|---|---|
| Traits | Root Type | −N | +N | −N | +N | C | N | C × N |
| Root density (kg m−3) | Total | 0.90 ± 0.08 | 1.16 ± 0.18 | 0.54 ± 0.07 | 0.68 ± 0.08 | *** | *** | N.S. |
| Rootstock | 0.37 ± 0.06 | 0.60 ± 0.20 | 0.28 ± 0.09 | 0.33 ± 0.03 | *** | *** | * | |
| Lateral | 0.41 ± 0.07 | 0.46 ± 0.09 | 0.2 ± 0.02 | 0.26 ± 0.06 | *** | N.S. | N.S. | |
| Fine | 0.12 ± 0.02 | 0.09 ± 0.02 | 0.06 ± 0.01 | 0.08 ± 0.02 | *** | N.S. | *** | |
| Root proportion to total root biomass (%) | Rootstock | 41.02 ± 6.27 | 51.9 ± 3.31 | 50.79 ± 4.78 | 49.32 ± 5.41 | N.S. | N.S. | * |
| Lateral | 13.6 ± 2.82 | 8.47 ± 3.12 | 11.29 ± 2.63 | 12.27 ± 1.96 | * | N.S. | N.S. | |
| Fine | 45.38 ± 5.08 | 39.63 ± 3.03 | 37.92 ± 2.45 | 38.41 ± 4.34 | N.S. | N.S. | * | |
| SRL | SRA | RTD | |||||
|---|---|---|---|---|---|---|---|
| N Loading | Variable | χ2 | p value | χ2 | p Value | χ2 | p Value |
| −N | Distance (D) | 4.23 | * | 1.79 | N.S. | 0.04 | N.S. |
| Compaction (C) | 0.06 | N.S. | 0.51 | N.S. | 3.90 | * | |
| D × C | 0.27 | N.S. | 0.08 | N.S. | 0.03 | N.S. | |
| +N | D | 0.27 | N.S. | 0.67 | N.S. | 0.85 | N.S. |
| C | 1.92 | N.S. | 0.38 | N.S. | 0.29 | N.S. | |
| D × C | 11.75 | *** | 5.18 | * | 0.74 | N.S. | |
| Control | High Compaction | ||||||
|---|---|---|---|---|---|---|---|
| −N | +N | −N | +N | ||||
| Wilcoxina | 33.87 | Wilcoxina | 31.42 | Wilcoxina | 34.07 | Suillus | 42.73 |
| Suillus | 23.44 | Suillus | 19.14 | Suillus | 16.31 | Wilcoxina | 15.86 |
| Laccaria | 10.10 | Dothideomycetes | 17.06 | Inocybe | 11.82 | Tetracladium | 10.77 |
| Helicorhoidion | 5.41 | Epicoccum | 8.81 | Tomentella | 8.34 | Saitozyma | 6.32 |
| Articulospora | 3.46 | Tomentella | 5.14 | Tricholoma | 7.08 | Cryptococcus | 3.45 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sugai, T.; Yokoyama, S.; Tamai, Y.; Mori, H.; Marchi, E.; Watanabe, T.; Satoh, F.; Koike, T. Evaluating Soil–Root Interaction of Hybrid Larch Seedlings Planted under Soil Compaction and Nitrogen Loading. Forests 2020, 11, 947. https://doi.org/10.3390/f11090947
Sugai T, Yokoyama S, Tamai Y, Mori H, Marchi E, Watanabe T, Satoh F, Koike T. Evaluating Soil–Root Interaction of Hybrid Larch Seedlings Planted under Soil Compaction and Nitrogen Loading. Forests. 2020; 11(9):947. https://doi.org/10.3390/f11090947
Chicago/Turabian StyleSugai, Tetsuto, Satoko Yokoyama, Yutaka Tamai, Hirotaka Mori, Enrico Marchi, Toshihiro Watanabe, Fuyuki Satoh, and Takayoshi Koike. 2020. "Evaluating Soil–Root Interaction of Hybrid Larch Seedlings Planted under Soil Compaction and Nitrogen Loading" Forests 11, no. 9: 947. https://doi.org/10.3390/f11090947
APA StyleSugai, T., Yokoyama, S., Tamai, Y., Mori, H., Marchi, E., Watanabe, T., Satoh, F., & Koike, T. (2020). Evaluating Soil–Root Interaction of Hybrid Larch Seedlings Planted under Soil Compaction and Nitrogen Loading. Forests, 11(9), 947. https://doi.org/10.3390/f11090947