
Gap Size in Hyrcanian Forest Affects the Lignin and N Concentrations of the Oriental Beech (Fagus orientalis Lipsky) Fine Roots but Does Not Change Their Morphological Traits in the Medium Term



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
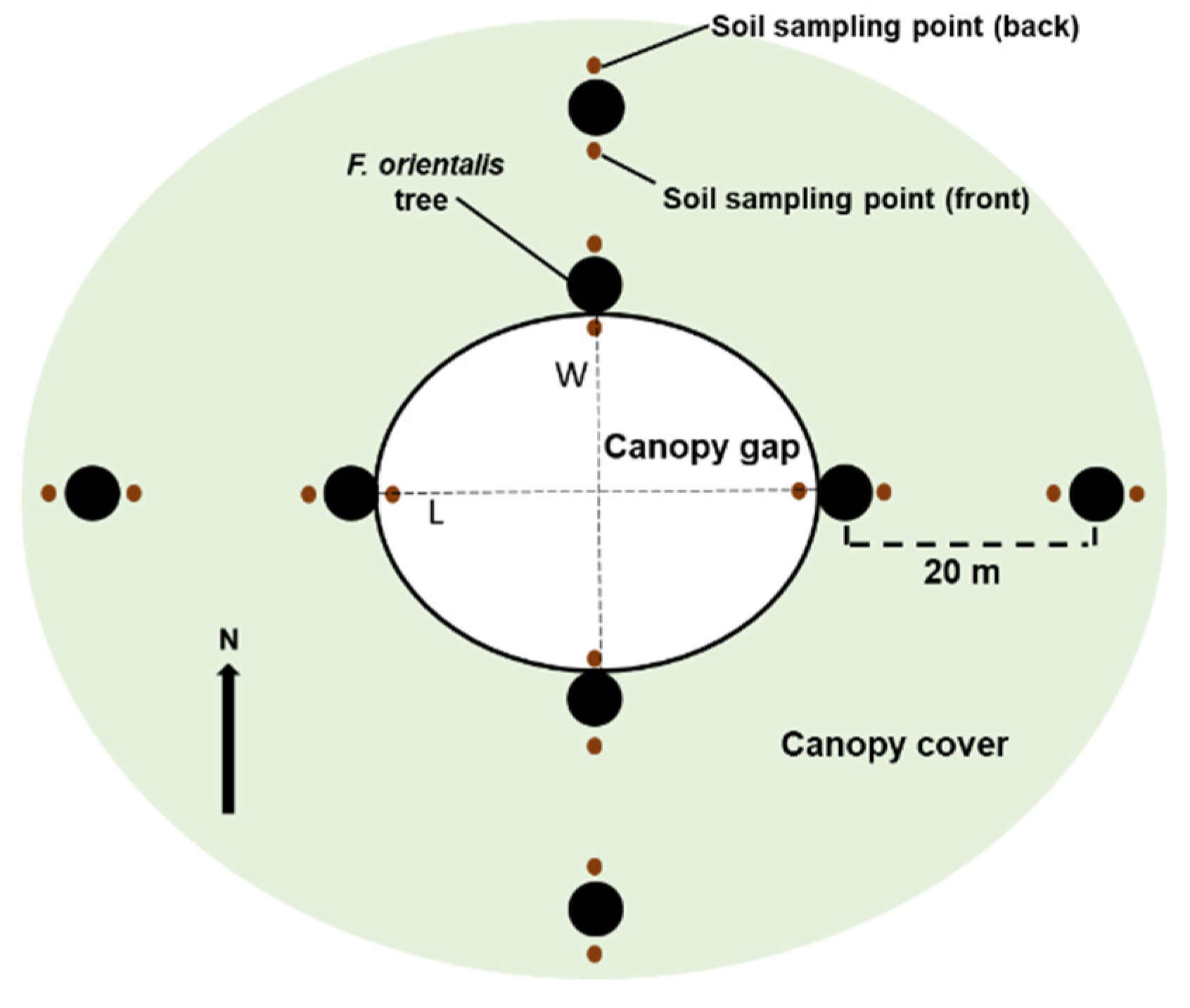
2.2. Experimental Design
2.3. Fine Root Sampling
2.4. Morphological Features of Fine Roots
2.5. Chemical Composition of Fine Roots
2.5.1. Carbon and Nitrogen
2.5.2. Cellulose and Lignin
2.6. Statistical Analyses
3. Results and Discussion
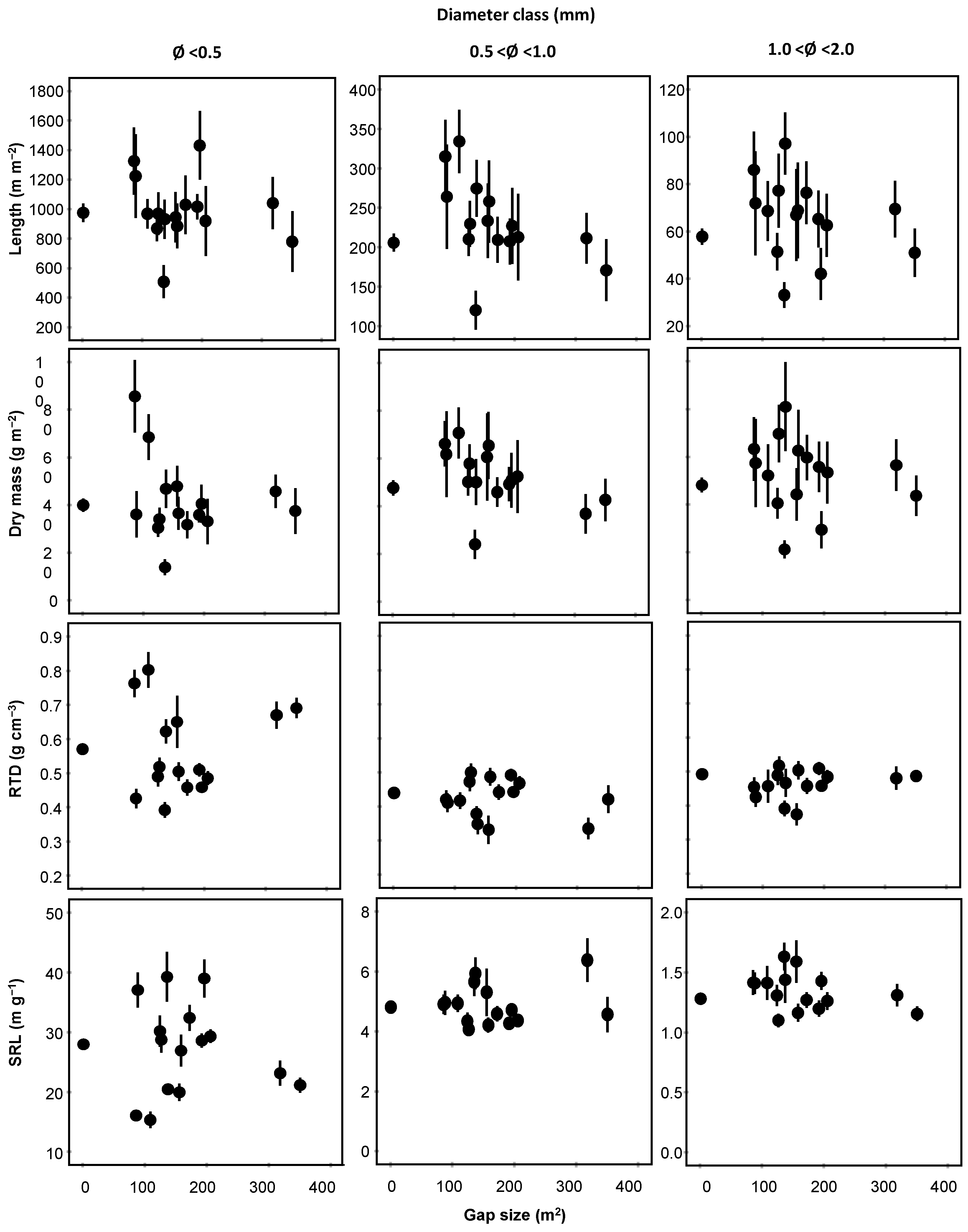
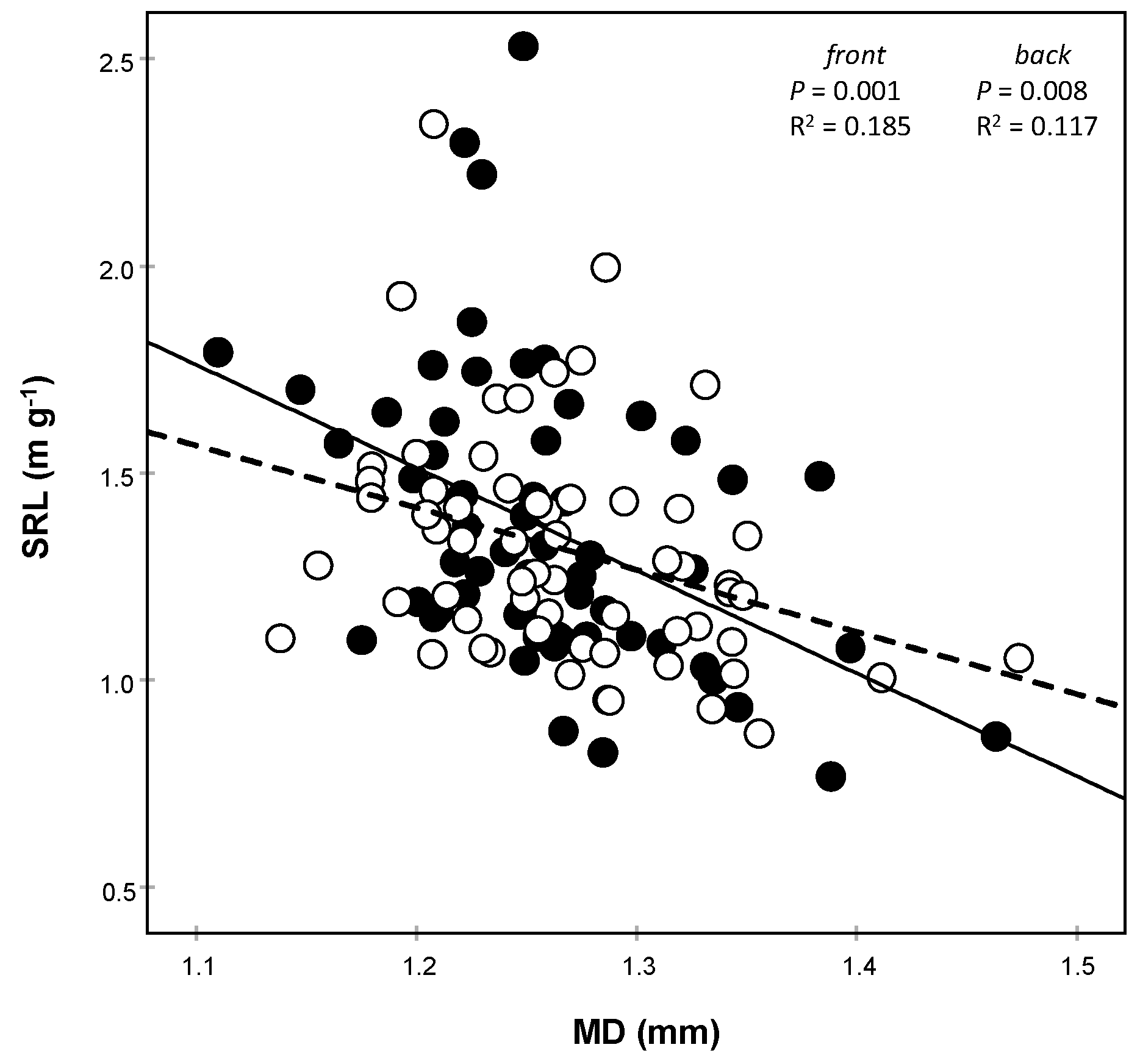
3.1. Morphological Traits
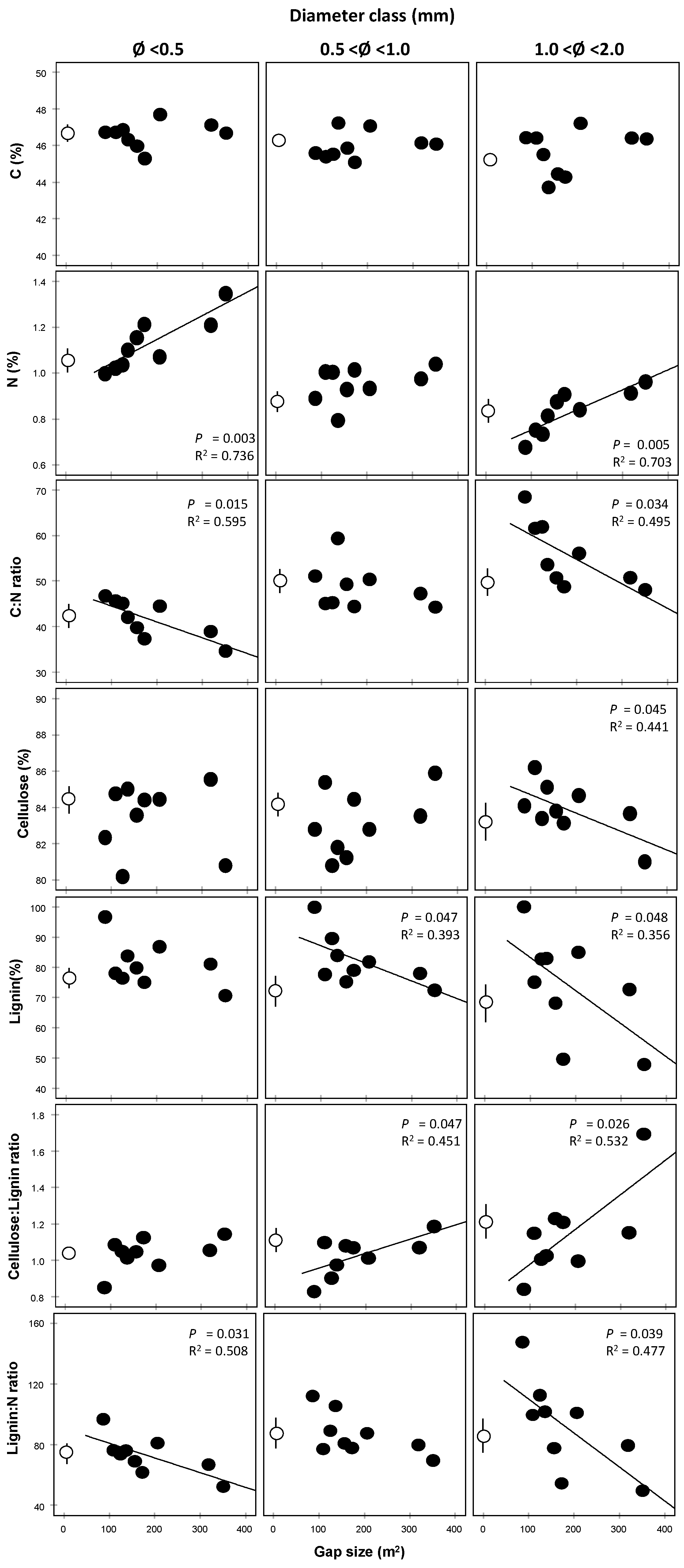
3.2. Chemical Traits
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, C.; Brunner, I.; Zong, S.; Li, M.-H. The Dynamics of Living and Dead Fine Roots of Forest Biomes Across the Northern Hemisphere. Forests 2019, 10, 953. [Google Scholar] [CrossRef]
- Finér, L.; Ohashi, M.; Noguchi, K.; Hirano, Y. Fine root production and turnover in forest ecosystems in relation to stand and environmental characteristics. For. Ecol. Manag. 2011, 262, 2008–2023. [Google Scholar] [CrossRef]
- Železnik, P.; Vilhar, U.; Starr, M.; De Groot, M.; Kraigher, H. Fine root dynamics in Slovenian beech forests in relation to soil temperature and water availability. Trees 2016, 30, 375–384. [Google Scholar] [CrossRef]
- Finér, L.; Ohashi, M.; Noguchi, K.; Hirano, Y. Factors causing variation in fine root biomass in forest ecosystems. For. Ecol. Manag. 2011, 261, 265–277. [Google Scholar] [CrossRef]
- Yuan, Z.; Chen, H.Y. Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, and stand age: Literature review and meta-analyses. Crit. Rev. Plant. Sci. 2010, 29, 204–221. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef]
- Vogt, K.A.; Vogt, D.J.; Bloomfield, J. Analysis of some direct and indirect methods for estimating root biomass and production of forests at an ecosystem level. Plant Soil 1998, 200, 71–89. [Google Scholar] [CrossRef]
- Bauhus, J.; Bartsch, N. Fine-root growth in beech (Fagus sylvatica) forest gaps. Can. J. For. Res. 1996, 26, 2153–2159. [Google Scholar] [CrossRef]
- Brockway, D.G.; Outcalt, K.W. Gap-phase regeneration in longleaf pine wiregrass ecosystems. For. Ecol. Manag. 1998, 106, 125–139. [Google Scholar] [CrossRef]
- Montagnoli, A.; Terzaghi, M.; Di Iorio, A.; Scippa, G.S.; Chiatante, D. Fine-root seasonal pattern, production and turnover rate of European beech (Fagus sylvatica L.) stands in Italy Prealps: Possible implications of coppice conversion to high forest. Plant Biosyst. 2012, 146, 1012–1022. [Google Scholar] [CrossRef]
- Jones, R.H.; Mitchell, R.J.; Stevens, G.N.; Pecot, S.D. Controls of fine root dynamics across a gradient of gap sizes in a pine woodland. Oecologia 2003, 134, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Di Iorio, A.; Montagnoli, A.; Terzaghi, M.; Scippa, G.S.; Chiatante, D. Effect of tree density on root distribution in Fagus sylvatica stands: A semi-automatic digitising device approach to trench wall method. Trees Struct. Funct. 2013, 27, 1503–1513. [Google Scholar] [CrossRef]
- Ostonen, I.; Püttsepp, Ü.; Biel, C.; Alberton, O.; Bakker, M.R.; Lõhmus, K.; Majdi, H.; Metcalfe, D.; Olsthoorn, A.F.M.; Pronk, A.; et al. Specific root length as an indicator of environmental change. Plant Biosyst. 2007, 141, 426–442. [Google Scholar] [CrossRef]
- Amendola, C.; Montagnoli, A.; Terzaghi, M.; Trupiano, D.; Oliva, F.; Baronti, S.; Miglietta, F.; Chiatante, D.; Scippa, G.S. Short-term effects of biochar on grapevine fine root dynamics and arbuscular mycorrhizae production. Agric. Ecosyst. Environ. 2017, 239, 236–245. [Google Scholar] [CrossRef]
- Withington, J.M.; Reich, P.B.; Oleksyn, J.; Eissenstat, D.M. Comparisons of structure and life span in roots and leaves among temperate trees. Ecol. Monogr. 2006, 76, 381–397. [Google Scholar] [CrossRef]
- Zadworny, M.; McCormack, M.L.; Rawlik, K.; Jagodziński, A.M. Seasonal variation in chemistry, but not morphology, in roots of Quercus robur growing in different soil types. Tree Physiol. 2015, 35, 644–652. [Google Scholar] [CrossRef]
- Li, A.; Guo, D.; Wang, Z.; Liu, H. Nitrogen and phosphorus allocation in leaves, twigs, and fine roots across 49 temperate, subtropical and tropical tree species: A hierarchical pattern. Funct. Ecol. 2010, 24, 224–232. [Google Scholar] [CrossRef]
- Terzaghi, M.; Montagnoli, A.; Di Iorio, A.; Scippa, G.S.; Chiatante, D. Fine-root carbon and nitrogen concentration of European beech (Fagus sylvatica L.) in Italy Prealps: Possible implications of coppice conversion to high forest. Front. Plant Sci. 2013, 4, 192. [Google Scholar] [CrossRef]
- Vajari, K.A.; Jalilvand, H.; Pourmajidian, M.R.; Espahbodi, K.; Moshki, A. The effect of single-tree selection system on soil properties in an oriental beech stand of Hyrcanian forest, north of Iran. J. For. Res. 2011, 22, 591–596. [Google Scholar] [CrossRef]
- Sommerfeld, R.; Lundquist, J.; Smith, J. Characterizing the canopy gap structure of a disturbed forest using the Fourier transform. For. Ecol. Manag. 2000, 128, 101–108. [Google Scholar] [CrossRef]
- Miller, S.D.; Goulden, M.L.; da Rocha, H.R. The effect of canopy gaps on subcanopy ventilation and scalar fluxes in a tropical forest. Agric. For. Meteorol. 2007, 142, 25–34. [Google Scholar] [CrossRef]
- Vepakomma, U.; St-Onge, B.; Kneeshaw, D. Spatially explicit characterization of boreal forest gap dynamics using multi-temporal lidar data. Remote Sens. Environ. 2008, 112, 2326–2340. [Google Scholar] [CrossRef]
- Kukkonen, M.; Rita, H.; Hohnwald, S.; Nygren, A. Treefall gaps of certified, conventionally managed and natural forests as regeneration sites for Neotropical timber trees in northern Honduras. For. Ecol. Manag. 2008, 255, 2163–2176. [Google Scholar] [CrossRef]
- Cannon, J.B.; Brewer, J.S. Effects of Tornado Damage, Prescribed Fire, and Salvage Logging on Natural Oak (Quercus spp.) Regeneration in a Xeric Southern USA Coastal Plain Oak and Pine Forest. Nat. Areas J. 2013, 33, 39–49. [Google Scholar] [CrossRef]
- Izbicki, B.J.; Alexander, H.D.; Paulson, A.K.; Frey, B.R.; McEwan, R.W.; Berry, A.I. Prescribed fire and natural canopy gap disturbances: Impacts on upland oak regeneration. For. Ecol. Manag. 2020, 465, 118107. [Google Scholar] [CrossRef]
- Gagnon, J.L.; Jokela, E.J.; Moser, W.; Huber, D.A. Characteristics of gaps and natural regeneration in mature longleaf pine flatwoods ecosystems. For. Ecol. Manag. 2004, 187, 373–380. [Google Scholar] [CrossRef]
- Yamamoto, S.-I. Forest Gap Dynamics and Tree Regeneration. J. For. Res. 2000, 5, 223–229. [Google Scholar] [CrossRef]
- Schnabel, F.; Donoso, P.J.; Winter, C. Short-term effects of single-tree selection cutting on stand structure and tree species composition in Valdivian rainforests of Chile. New Zeal. J. For. Sci. 2017, 47, 21. [Google Scholar] [CrossRef]
- Muscolo, A.; Bagnato, S.; Sidari, M.; Mercurio, R. A review of the roles of forest canopy gaps. J. For. Res. 2014, 25, 725–736. [Google Scholar] [CrossRef]
- Zhu, J.J.; Tan, H.; Li, F.Q.; Chen, M.; Zhang, J.X. Microclimate regimes following gap formation in a montane secondary forest of eastern Liaoning Province, China. J. For. Res. 2007, 18, 167–173. [Google Scholar] [CrossRef]
- Kooch, Y.; Bayranvand, M. Composition of tree species can mediate spatial variability of C and N cycles in mixed beech forests. For. Ecol. Manag. 2017, 401, 55–64. [Google Scholar] [CrossRef]
- Guner, S.; Yagci, V.; Tilki, F.; Celik, N. The effects of initial planting density on above-and below-ground biomass in a 25-year-old Fagus orientalis Lipsky plantation in Hopa, Turkey. Sci. Res. Essays 2010, 5, 1856–1860. [Google Scholar]
- Brassard, B.W.; Chen, H.Y.H.; Bergeron, Y. Influence of environmental variability on root dynamics in northern forests. CRC Crit. Rev. Plant Sci. 2009, 28, 179–197. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; DeForest, J.L.; Burton, A.J.; Allen, M.F.; Ruess, R.W.; Hendrick, R.L. Fine root architecture of nine North American trees. Ecol. Monogr. 2002, 72, 293–309. [Google Scholar] [CrossRef]
- Guo, D.; Xia, M.; Wei, X.; Chang, W.; Liu, Y.; Wang, Z. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytol. 2008, 180, 673–683. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; Zak, D.R.; Loya, W.M.; Karberg, N.J.; King, J.S.; Burton, A.J. The Contribution of Root-Rhizosphere Interactions to Biogeochemical Cycles in a Changing World. In The Rhizosphere: An Ecological Perspective; Cardon, Z.G., Whitbeck, J.L., Eds.; Elsevier Academic Press: Burlington, MA, USA, 2007; pp. 155–178. ISBN 9780120887750. [Google Scholar]
- Withington, J.M.; Elkin, A.D.; Bułaj, B.; Olesiński, J.; Tracy, K.N.; Bouma, T.J.; Oleksyn, J.; Anderson, L.J.; Modrzyński, J.; Reich, P.B.; et al. The impact of material used for minirhizotron tubes for root research. New Phytol. 2003, 160, 533–544. [Google Scholar] [CrossRef]
- Long, Y.; Kong, D.; Chen, Z.; Zeng, H. Variation of the linkage of root function with root branch order. PLoS ONE 2013, 8, e57153. [Google Scholar] [CrossRef]
- Iversen, C.M. Using root form to improve our understanding of root function. New Phytol. 2014, 203, 707–709. [Google Scholar] [CrossRef]
- Montagnoli, A.; Terzaghi, M.; Giussani, B.; Scippa, G.S.; Chiatante, D. An integrated method for high-resolution definition of new diameter-based fine root sub-classes of Fagus sylvatica L. Ann. For. Sci. 2018, 75, 76. [Google Scholar] [CrossRef]
- Montagnoli, A.; Terzaghi, M.; Di Iorio, A.; Scippa, G.S.; Chiatante, D. Fine-root morphological and growth traits in a Turkey-oak stand in relation to seasonal changes in soil moisture in the Southern Apennines, Italy. Ecol. Res. 2012, 27, 1015–1025. [Google Scholar] [CrossRef]
- Montagnoli, A.; Di Iorio, A.; Terzaghi, M.; Trupiano, D.; Scippa, G.S.; Chiatante, D. Influence of soil temperature and water content on fine-root seasonal growth of European beech natural forest in Southern Alps, Italy. Eur. J. For. Res. 2014, 133, 957–968. [Google Scholar] [CrossRef]
- Montagnoli, A.; Dumroese, R.K.; Terzaghi, M.; Onelli, E.; Scippa, G.S.; Chiatante, D. Seasonality of fine root dynamics and activity of root and shoot vascular cambium in a Quercus ilex L. forest (Italy). For. Ecol. Manag. 2019, 431, 26–34. [Google Scholar] [CrossRef]
- Yoshida, T.; Naito, S.; Nagumo, M.; Hyodo, N.; Inoue, T.; Umegane, H.; Yamazaki, H.; Miya, H.; Nakamura, F. Structural Complexity and Ecosystem Functions in a Natural Mixed Forest under a Single-Tree Selection Silviculture. Sustainability 2017, 9, 2093. [Google Scholar] [CrossRef]
- Alireza, A.; Abrari, V.K.; Mohammad, F.; Antonino, D.I. Influences of forest gaps on soil physico-chemical and biological properties in an oriental beech (Fagus orientalis L.) stand of Hyrcanian forest, north of Iran. iForest 2020, 13, 124–129. [Google Scholar]
- Leavitt, S.W.; Danzer, S.R. Method for Batch Processing Small Wood Samples to Holocellulose for Stable-Carbon Isotope Analysis. Anal. Chem. 1993, 65, 87–89. [Google Scholar] [CrossRef]
- Genet, M.; Stokes, A.; Salin, F.; Mickovski, S.B.; Fourcaud, T.; Dumail, J.F.; Van Beek, R. The influence of cellulose content on tensile strength in tree roots. Plant Soil 2005, 278, 1–9. [Google Scholar] [CrossRef]
- Doster, M.A.; Bostock, R.M. Quantification of Lignin Formation in Almond Bark in Response to Wounding and Infection by Phytophthora Species. Phytopathology 1988, 78, 473. [Google Scholar] [CrossRef]
- Trupiano, D.; Di Iorio, A.; Montagnoli, A.; Lasserre, B.; Rocco, M.; Grosso, A.; Scaloni, A.; Marra, M.; Chiatante, D.; Scippa, G.S. Involvement of lignin and hormones in the response of woody poplar taproots to mechanical stress. Physiol. Plant. 2012, 146, 39–52. [Google Scholar] [CrossRef]
- Cambi, M.; Certini, G.; Neri, F.; Marchi, E. The impact of heavy traffic on forest soils: A review. For. Ecol. Manag. 2015, 338, 124–138. [Google Scholar] [CrossRef]
- Gray, A.N.; Spies, T.A.; Easter, M.J. Microclimatic and soil moisture responses to gap formation in coastal Douglas-fir forests. Can. J. For. Res. 2002, 32, 332–343. [Google Scholar] [CrossRef]
- Terzaghi, M.; Di Iorio, A.; Montagnoli, A.; Baesso, B.; Scippa, G.S.; Chiatante, D. Forest canopy reduction stimulates xylem production and lowers carbon concentration in fine roots of European beech. For. Ecol. Manag. 2016, 379, 81–90. [Google Scholar] [CrossRef]
- Ni, X.; Berg, B.; Yang, W.; Li, H.; Liao, S.; Tan, B.; Yue, K.; Xu, Z.; Zhang, L.; Wu, F. Formation of forest gaps accelerates C, N and P release from foliar litter during 4 years of decomposition in an alpine forest. Biogeochemistry 2018, 139, 321–335. [Google Scholar] [CrossRef]
- Scharenbroch, B.C.; Bockheim, J.G. Impacts of forest gaps on soil properties and processes in old growth northern hardwood-hemlock forests. Plant Soil 2007, 294, 219–233. [Google Scholar] [CrossRef]
- Parsons, W.F.J.; Knight, D.H.; Miller, S.L. Root Gap Dynamics in Lodgepole Pine Forest: Nitrogen Transformations in Gaps of Different Size. Ecol. Appl. 1994, 4, 354–362. [Google Scholar] [CrossRef]
- Thomas, F.M.; Molitor, F.; Werner, W. Lignin and cellulose concentrations in roots of Douglas fir and European beech of different diameter classes and soil depths. Trees—Struct. Funct. 2014, 28, 309–315. [Google Scholar] [CrossRef]
- Zhang, C.B.; Chen, L.H.; Jiang, J. Why fine tree roots are stronger than thicker roots: The role of cellulose and lignin in relation to slope stability. Geomorphology 2014, 206, 196–202. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, Y.; Wang, Y.; Mao, Z.; Langendoen, E.J. How does root biodegradation after plant felling change root reinforcement to soil? Plant Soil 2020, 446, 211–227. [Google Scholar] [CrossRef]
- McCormack, M.L.; Iversen, C.M. Physical and Functional Constraints on Viable Belowground Acquisition Strategies. Front. Plant Sci. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Kong, D.; Wang, J.; Wu, H.; Valverde-Barrantes, O.J.; Wang, R.; Zeng, H.; Kardol, P.; Zhang, H.; Feng, Y. Nonlinearity of root trait relationships and the root economics spectrum. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Kramer-Walter, K.R.; Bellingham, P.J.; Millar, T.R.; Smissen, R.D.; Richardson, S.J.; Laughlin, D.C. Root traits are multidimensional: Specific root length is independent from root tissue density and the plant economic spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- Fayle, D.C.F. Distribution of radial growth during the development of red pine root systems. Can. J. For. Res. 1975, 5, 608–625. [Google Scholar] [CrossRef]
- Vincent, M.; Krause, C.; Zhang, S.Y. Radial growth response of black spruce roots and stems to commercial thinning in the boreal forest. Forestry 2009, 82, 557–571. [Google Scholar] [CrossRef]
- Talbot, J.M.; Treseder, K.K. Interactions among lignin, cellulose, and nitrogen drive litter chemistry-decay relationships. Ecology 2012, 93, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Walela, C.; Daniel, H.; Wilson, B.; Lockwood, P.; Cowie, A.; Harden, S. The initial lignin: Nitrogen ratio of litter from above and below ground sources strongly and negatively influenced decay rates of slowly decomposing litter carbon pools. Soil Biol. Biochem. 2014, 77, 268–275. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W. The decomposition of fine and coarse roots: Their global patterns and controlling factors. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef]
- Lützow, M.V.; Kögel-Knabner, I.; Ekschmitt, K.; Matzner, E.; Guggenberger, G.; Marschner, B.; Flessa, H. Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions—A review. Eur. J. Soil Sci. 2006, 57, 426–445. [Google Scholar] [CrossRef]
- Valverde-Barrantes, O.J.; Smemo, K.A.; Blackwood, C.B. Fine root morphology is phylogenetically structured, but nitrogen is related to the plant economics spectrum in temperate trees. Funct. Ecol. 2015, 29, 796–807. [Google Scholar] [CrossRef]
- Comas, L.H.; Eissenstat, D.M.; Lakso, A.N. Assessing root death and root system dynamics in a study of grape canopy pruning. New Phytol. 2000, 147, 171–178. [Google Scholar] [CrossRef]
- Lee, M.H.; Comas, L.H.; Callahan, H.S. Experimentally reduced root-microbe interactions reveal limited plasticity in functional root traits in Acer and Quercus. Ann. Bot. 2014, 113, 513–521. [Google Scholar] [CrossRef]
- Comas, L.H.; Eissenstat, D.M. Patterns in root trait variation among 25 co-existing North American forest species. New Phytol. 2009, 182, 919–928. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fine Root Trait | Diameter (mm) | Core Position | Orientation | Gap Size (c) | |||
|---|---|---|---|---|---|---|---|
| (df = 1) | (df = 3) | (df = 1) | |||||
| F | p | F | p | F | p | ||
| Length (m m−2) | 0.5 | 0.125 | 0.724 | 0.343 | 0.794 | 0.087 | 0.768 |
| 0.5–1 | 0.054 | 0.817 | 0.031 | 0.993 | 0.152 | 0.697 | |
| 1–2 | 0.274 | 0.601 | 0.163 | 0.921 | 0.510 | 0.476 | |
| Dry mass (g m−2) | 0.5 | 0.009 | 0.924 | 0.329 | 0.805 | 0.007 | 0.935 |
| 0.5–1 | 0.067 | 0.796 | 0.338 | 0.798 | 0.030 | 0.862 | |
| 1–2 | 0.843 | 0.360 | 0.207 | 0.891 | 0.253 | 0.616 | |
| RTD (g cm−3) | 0.5 | 0.405 | 0.525 | 0.387 | 0.762 | 0.137 | 0.712 |
| 0.5–1 | 0.096 | 0.756 | 1.220 | 0.303 | 1.387 | 0.240 | |
| 1–2 | 1.082 | 0.299 | 0.822 | 0.483 | 0.844 | 0.359 | |
| SRL (m g−1) | 0.5 | 0.687 | 0.408 | 0.434 | 0.729 | 0.804 | 0.371 |
| 0.5–1 | 0.001 | 0.998 | 0.671 | 0.571 | 0.659 | 0.418 | |
| 1–2 | 3.296 | 0.071 | 1.230 | 0.299 | 0.135 | 0.714 | |
| Chemical Trait | Diam Class (df = 2) | Gap Size (c) (df = 1) | ||
|---|---|---|---|---|
| F | p | F | p | |
| C | 2.598 | 0.096 | 1.596 | 0.219 |
| N | 43.73 | <0.001 | 25.28 | <0.001 |
| C:N | 20.89 | <0.001 | 3.05 | 0.087 |
| Cellulose | 0.374 | 0.692 | 0.024 | 0.879 |
| Lignin | 1.747 | 0.197 | 9.371 | 0.006 |
| Cellulose:Lignin | 2.299 | 0.123 | 13.442 | 0.001 |
| Lignin: N | 3.209 | 0.059 | 14.54 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kondori, A.A.; Vajari, K.A.; Feizian, M.; Montagnoli, A.; Di Iorio, A. Gap Size in Hyrcanian Forest Affects the Lignin and N Concentrations of the Oriental Beech (Fagus orientalis Lipsky) Fine Roots but Does Not Change Their Morphological Traits in the Medium Term. Forests 2021, 12, 137. https://doi.org/10.3390/f12020137
Kondori AA, Vajari KA, Feizian M, Montagnoli A, Di Iorio A. Gap Size in Hyrcanian Forest Affects the Lignin and N Concentrations of the Oriental Beech (Fagus orientalis Lipsky) Fine Roots but Does Not Change Their Morphological Traits in the Medium Term. Forests. 2021; 12(2):137. https://doi.org/10.3390/f12020137
Chicago/Turabian StyleKondori, Alireza Amoli, Kambiz Abrari Vajari, Mohammad Feizian, Antonio Montagnoli, and Antonino Di Iorio. 2021. "Gap Size in Hyrcanian Forest Affects the Lignin and N Concentrations of the Oriental Beech (Fagus orientalis Lipsky) Fine Roots but Does Not Change Their Morphological Traits in the Medium Term" Forests 12, no. 2: 137. https://doi.org/10.3390/f12020137
APA StyleKondori, A. A., Vajari, K. A., Feizian, M., Montagnoli, A., & Di Iorio, A. (2021). Gap Size in Hyrcanian Forest Affects the Lignin and N Concentrations of the Oriental Beech (Fagus orientalis Lipsky) Fine Roots but Does Not Change Their Morphological Traits in the Medium Term. Forests, 12(2), 137. https://doi.org/10.3390/f12020137