Structural Carbon Allocation and Wood Growth Reflect Climate Variation in Stands of Hybrid White Spruce in Central Interior British Columbia, Canada

Abstract
:1. Introduction
2. Materials and Methods
3. Results
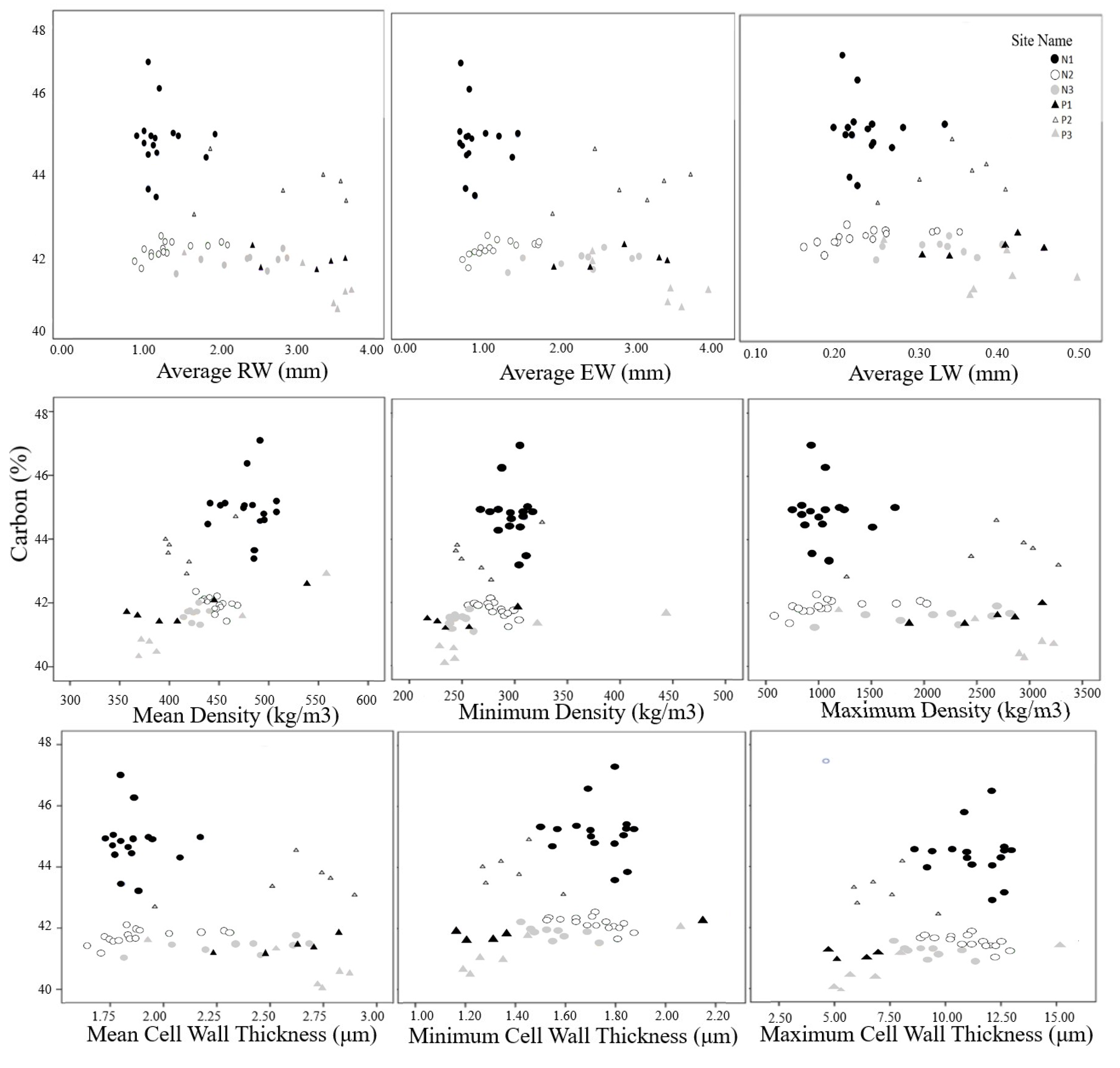
3.1. Percent Carbon vs. Radial Growth Variables
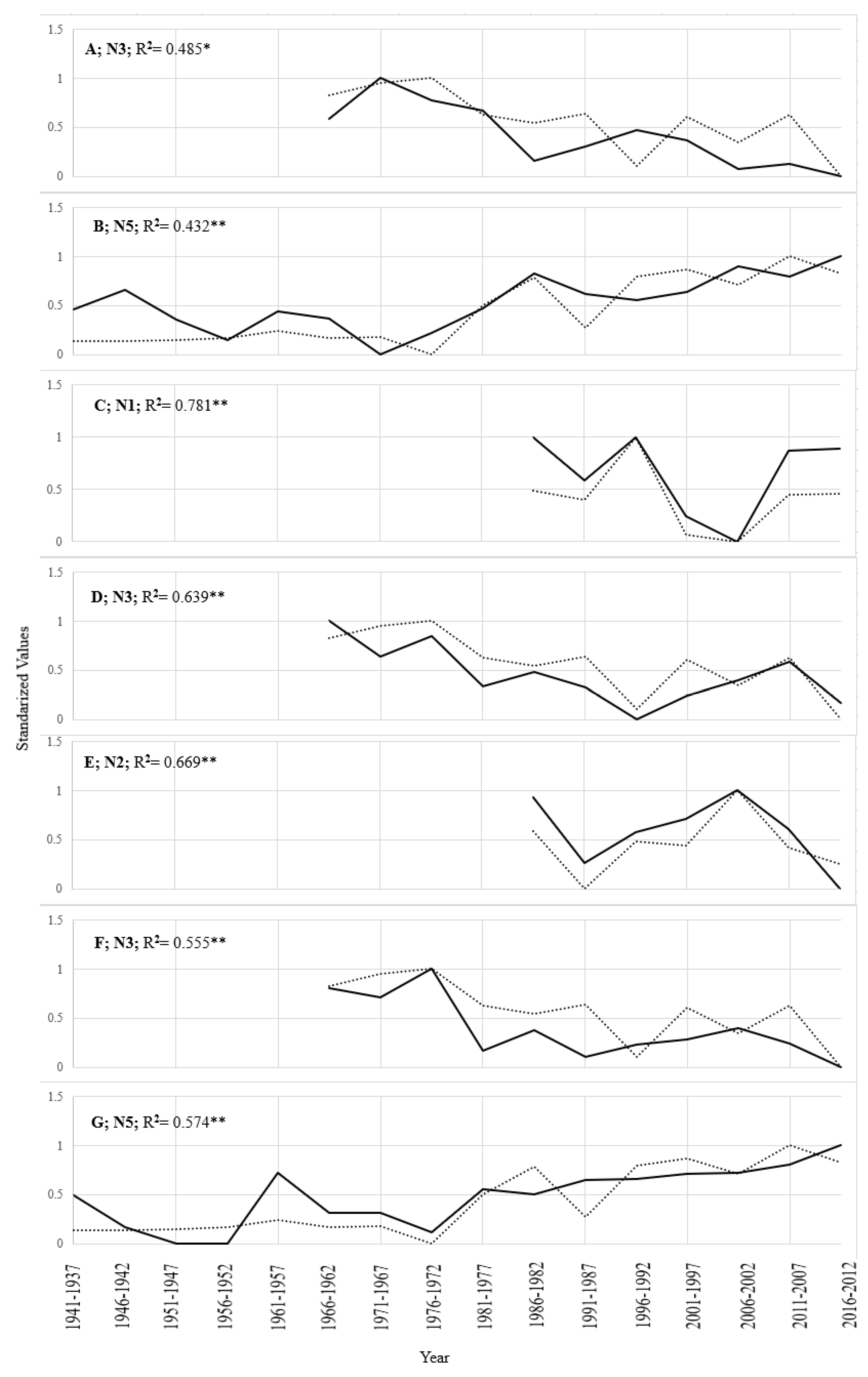
3.2. Percent Carbon vs. Climate
4. Discussion
4.1. Percent Carbon vs. Radial Growth Variables
4.2. Percent Carbon vs. Climate
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weber, J.C.; Sotelo Montes, C.; Abasse, T.; Sanquetta, C.R.; Silva, D.A.; Mayer, S.; Muniz, G.I.B.; Garcia, R.A. Variation in growth, wood density and carbon concentration in five tree and shrub species in Niger. New For. 2018, 49, 35–51. [Google Scholar]
- Elias, M.; Potvin, C. Assessing inter- and intra-specific variation in trunk carbon concentration for 32 neotropical tree species. Can. J. Res. 2003, 33, 1039–1045. [Google Scholar]
- Thomas, S.C.; Malczewski, G. Wood carbon content of tree species in eastern China: Interspecific variability and the importance of the volatile fraction. J. Environ. Manag. 2007, 85, 659–662. [Google Scholar]
- Lachenbruch, B.; Mcculloh, K.A. Tansley review Traits, properties, and performance: How mechanical functions in a cell, tissue, or whole. New Phytol. 2014, 204, 747–764. [Google Scholar]
- Zabek, L.M.; Prescott, C.E. Biomass equations and carbon content of aboveground leafless biomass of hybrid poplar in coastal British Columbia. For. Ecol. Manag. 2006, 223, 291–302. [Google Scholar]
- Castaño-Santamaría, J.; Bravo, F. Variation in carbon concentration and basic density along stems of sessile oak (Quercus petraea (Matt.) Liebl.) and Pyrenean oak (Quercus pyrenaica Willd.) in the Cantabrian Range (NW Spain). Ann. For. Sci. 2012, 69, 663–672. [Google Scholar]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Bigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar]
- Schwalm, C.R.; Williams, C.A.; Schaefer, K.; Arneth, A.; Bonal, D.; Buchmann, N.; Chen, J.; Law, B.L.; Lindroth, A.; Luyssaert, S.; et al. Assimilation exceeds respiration sensitivity to drought: A FLUXNET synthesis. Glob. Chang. Biol. 2010, 16, 657–670. [Google Scholar]
- Haughian, S.R.; Burton, P.J.; Taylor, S.W.; Curry, C. Expected effects of climate change on forest disturbance regimes in British Columbia. J. Ecosyst. Manag. 2012, 13, 1–24. [Google Scholar]
- Babst, F.; Alexander, M.R.; Szejner, P.; Bouriaud, O.; Klesse, S.; Roden, J.; Ciais, P.; Poulter, B.; Frank, D.; Moore, D.J.P.; et al. A tree-ring perspective on the terrestrial carbon cycle. Oecologia 2014, 176, 307–322. [Google Scholar]
- McDowell, N.G.; Allen, C.D. Darcy’s law predicts widespread forest mortality under climate warming. Nat. Clim. Chang. 2015, 5, 669–672. [Google Scholar]
- Lo, Y.H.; Blanco, J.A.; Seely, B.; Welham, C.; Kimmins, J.P. Relationships between climate and tree radial growth in interior British Columbia, Canada. For. Ecol. Manag. 2010, 259, 932–942. [Google Scholar]
- Fleming, S.W.; Whitfield, P.H. Spatiotemporal mapping of ENSO and PDO surface meteorological signals in British Columbia, Yukon, and southeast Alaska Spatiotemporal Mapping of ENSO and PDO Surface Meteorological Signals in British Columbia, Yukon, and Southeast Alaska. Can. Meteorol. Oceanogr. Soc. 2010, 48, 122–131. [Google Scholar]
- Jiang, X.; Huang, J.; Stadt, K.J.; Comeau, P.G.; Chen, H.Y.H. Spatial climate-dependent growth response of boreal mixedwood forest in western Canada. Glob. Planet. Chang. 2016, 139, 141–150. [Google Scholar]
- Millar, C.I.; Stephenson, N.L. Temperate forest health in an era of emerging megadisturbance. Science 2015, 349, 823–826. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press Inc.: New York, NY, USA, 1976. [Google Scholar]
- Bouriaud, O.; Bréda, N.; Dupouey, J.-L.; Granier, A. Is ring width a reliable proxy for stem-biomass increment? A case study in European beech. Can. J. For. Res. 2005, 35, 2920–2933. [Google Scholar]
- Grünwald, T.; Bernhofer, C. A decade of carbon, water and energy flux measurements of an old spruce forest at the Anchor Station Tharandt. Chem. Phys. Meteorol. 2007, 59, 387–396. [Google Scholar]
- Liepiņš, J.; Lazdiņš, A.; Liepiņš, K. Equations for estimating above- and belowground biomass of Norway spruce, Scots pine, birch spp. and European aspen in Latvia. Scand. J. For. Res. 2018, 33, 58–70. [Google Scholar]
- Davis, S.C.; Hessl, A.E.; Scott, C.J.; Adams, M.B.; Thomas, R.B. Forest carbon sequestration changes in response to timber harvest. For. Ecol. Manag. 2009, 258, 2101–2109. [Google Scholar]
- Ter-Mikaelian, M.T.; Colombo, S.J.; Chen, J. Effect of age and disturbance on decadal changes in carbon stocks in managed forest landscapes in central Canada. Mitig. Adapt. Strateg. Glob. Chang. 2014, 19, 1063–1075. [Google Scholar]
- Meidinger, D.; Pojar, J. Ecosystems of British Columbia; BC Ministry of Forests: Victoria, BC, Canada, 1991. [Google Scholar]
- Delong, C.; Tanner, D.; Jull, M.J. A Field Guide for Site Identification and Interpretation for the Southwest Portion of the Prince George Forest Region; Province of Britsh Columbia Minsitry of Forests: Prince George, BC, Canada, 1992. [Google Scholar]
- Dale, M.R.T.; Fortin, M.J. Spatial Analysis: A Guide for Ecologists; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Jensen, W.B. The Origin of the Soxhlet Extractor. J. Chem. Educ. 2007, 84, 1913–1914. [Google Scholar]
- Grabner, M.; Wimmer, R.; Gierlinger, N.; Evans, R.; Downes, G. Heartwood extractives in larch and effects on x-ray densitometry. Can. J. For. Res. 2005, 35, 2781–2786. [Google Scholar]
- Evans, R.; Ilic, J. Rapid prediction of wood stiffness from microfibril angle and density. For. Prod. J. 2001, 51, 53. [Google Scholar]
- Yamaguchi, D.K. A simple method for cross-dating cores from living trees. Can. J. Res 1991, 21, 414–416. [Google Scholar]
- Wood, L.J.; Smith, D.J. Climate and glacier mass balance trends from AD 1780 to present in the Columbia mountians, British Columbia, Canada. Holocene 2012, 23, 739–748. [Google Scholar]
- Coates, D.K.; Haeussler, S.; Lindeburgh, S.; Pojar, R.; Stock, A.J. Ecology and Silviculture of Interior Spruce in British Columbia; Government of Canada: British Columbia, BC, Canada, 1994; pp. 1–182. [Google Scholar]
- Jones, D.A.; Hara, K.L.O. The influence of preparation method on measured carbon fractions in tree tissues. Tree Physiol. 2017, 36, 1177–1189. [Google Scholar]
- Beck, P.S.A.; Andreu-Hayles, L.; D’Arrigo, R.; Anchukaitis, K.J.; Tucker, C.J.; Pinzón, J.E.; Goetz, S.J. A large-scale coherent signal of canopy status in maximum latewood density of tree rings at arctic treeline in North America. Glob. Planet. Chang. 2013, 100, 109–118. [Google Scholar]
- Navarro, M.; Moya, R.; Chazdon, R.; Ortiz, E.; Vilchez, B. Successional variation in carbon content and wood specific gravity of four tropical tree species. Bosque 2013, 34, 9–10. [Google Scholar]
- Clark, D.A.; Piper, S.C.; Keeling, C.D.; Clark, D.B. Tropical rain forest tree growth and atmospheric carbon dynamics linked to interannual temperature variation during 1984–2000. Proc. Natl. Acad. Sci. USA 2003, 100, 5852–5857. [Google Scholar]
- Somogyi, Z.; Cienciala, E.; Mäkipää, R.; Muukkonen, P.; Lehtonen, A.; Weiss, P. Indirect methods of large-scale forest biomass estimation. Eur. J. For. Res. 2007, 126, 197–207. [Google Scholar]
- Kearsley, E.; De Haulleville, T.; Hufkens, K.; Kidimbu, A.; Toirambe, B.; Baert, G.; Huygens, D.; Kebede, Y.; Defourny, P.; Bogaert, J.; et al. Conventional tree height-diameter relationships significantly overestimate aboveground carbon stocks in the Central Congo Basin. Nat. Commun. 2013, 4, 1–8. [Google Scholar]
- Ali, A.; Yan, E.R.; Chen, H.Y.H.; Chang, S.X.; Zhao, Y.T.; Yang, X.D.; Xu, M.S. Stand structural diversity rather than species diversity enhances aboveground carbon storage in secondary subtropical forests in Eastern China. Biogeosciences 2016, 13, 4627–4635. [Google Scholar]
- Zhang, Q.; Wang, C.; Wang, X.; Quan, X. Carbon concentration variability of 10 Chinese temperate tree species. For. Ecol. Manag. 2009, 258, 722–727. [Google Scholar]
- Martin, A.R.; Thomas, S.C. A reassessment of carbon content in tropical trees. PLoS ONE 2011, 6, e23533. [Google Scholar]
- Bao, F.C.; Jiang, Z.H.; Jiang, X.M.; Lu, X.; Luo, X.Q.; Zhang, S.Y. Differences in wood properties between juvenile wood and mature wood in 10 species grown in China. Wood Sci. Technol. 2001, 35, 363–375. [Google Scholar]
- Zobel, B.J.; van Buijtenen, J.P. Wood Variation: Its Causes and Controls; Springer: Berlin/Heidelberg, Germany, 1989. [Google Scholar]
- Bert, D.; Danjon, F. Carbon concentration variations in the roots, stem and crown of mature Pinus pinaster (Ait.). For. Ecol. Manag. 2006, 222, 279–295. [Google Scholar]
- Lamlom, S.H.; Savidge, R.A. A reassessment of carbon content in wood: Variation within and between 41 North American species. Biomass Bioenergy 2003, 25, 381–388. [Google Scholar]
- Gower, S.T.; Krankina, I.O.; Olson, R.J.; Apps, M.; Linder, S.; Wangi, C. Net primary production and carbon allocation patterns of boreal forest ecosystems. Ecol. Appl. 2001, 11, 1395–1411. [Google Scholar]
- Wang, C.; Bond-Lamberty, B.; Gower, S.T. Carbon distribution of a well- and poorly-drained black spruce fire chronosequence. Glob. Chang. Biol. 2003, 9, 1066–1079. [Google Scholar]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Morin, H.; Saracino, A.; Motta, R.; Borghetti, M. Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length. New Phytol. 2006, 170, 301–310. [Google Scholar]
- Guehl, J.M. Étude comparée des potentialités hivernales d’assimilation carbonée de trois conifères de la zone tempérée (Pseudotsuga menziesii Mirb., Abies alba Mill. et Picea excelsa Link.). Ann. Sci. 1985, 42, 23–38. [Google Scholar]
- McMahon, M.S.; Parker, G.G.; Miller, D.R. Evidence for a recent increase in forest growth is questionable. Proc. Natl. Acad. Sci. USA 2010, 107, 3611–3615. [Google Scholar]
- Hember, R.A.; Kurz, W.A.; Metsaranta, J.M.; Black, T.; Guy, R.D.; Coops, N.C. Accelerating regrowth of temperate-maritime forests due to environmental change. Glob. Chang. Biol. 2012, 18, 2026–2040. [Google Scholar]
- Jarvis, P.; Linder, S. Constraints to growth of boreal forests. Nature 2000, 405, 904–905. [Google Scholar]
- Knudson, D.V.; Lindsey, C. Type I and Type II errors in correlations of various sample sizes. Compr. Psychol. 2014, 3, 2165–2228. [Google Scholar]
- Zhang, Q.; Alfaro, R.I.; Hebda, R.J. Dendroecological studies of tree growth, climate and spruce beetle outbreaks in central British Columbia, Canada. For. Ecol. Manag. 1999, 121, 215–225. [Google Scholar]
- Oberhuber, W.; Gruber, A.; Kofler, W.; Swidrak, I. Radial stem growth in response to microclimate and soil moisture in a drought-prone mixed coniferous forest at an inner Alpine site. Eur. J. For. Res. 2014, 133, 467–479. [Google Scholar]
- Peterson, D.W.; Peterson, D.L.; Ettl, G.L. Growth responses of subalpine fir to climate variability in the pacific northwest. Can. J. For. Res. 2002, 32, 1503–1517. [Google Scholar]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar]
- Martin-Benito, D.; Beeckman, H.; Cañellas, I. Influence of drought on tree rings and tracheid features of Pinus nigra and Pinus sylvestris in a mesic Mediterranean forest. Eur. J. For. Res. 2013, 132, 33–45. [Google Scholar]
- Peñuelas, J.; Ogaya, R.; Boada, M.; Jump, A.S. Migration, invasion and decline: Changes in recruitment and forest structure in a warming-linked shift of European beech forest in Catalonia (NE Spain). Ecography 2007, 30, 829–837. [Google Scholar]
- Lazarus, B.E.; Castanha, C.; Germino, M.J.; Kueppers, L.M.; Moyes, A.B. Growth strategies and threshold responses to water deficit modulate effects of warming on tree seedlings from forest to alpine. J. Ecol. 2018, 106, 571–585. [Google Scholar]
- Li, T.; Ren, B.; Wang, D.; Liu, G. Spatial variation in the storages and age-related dynamics of forest carbon sequestration in different climate zones-evidence from black locust plantations on the loess plateau of China. PLoS ONE 2015, 10, e0121862. [Google Scholar]
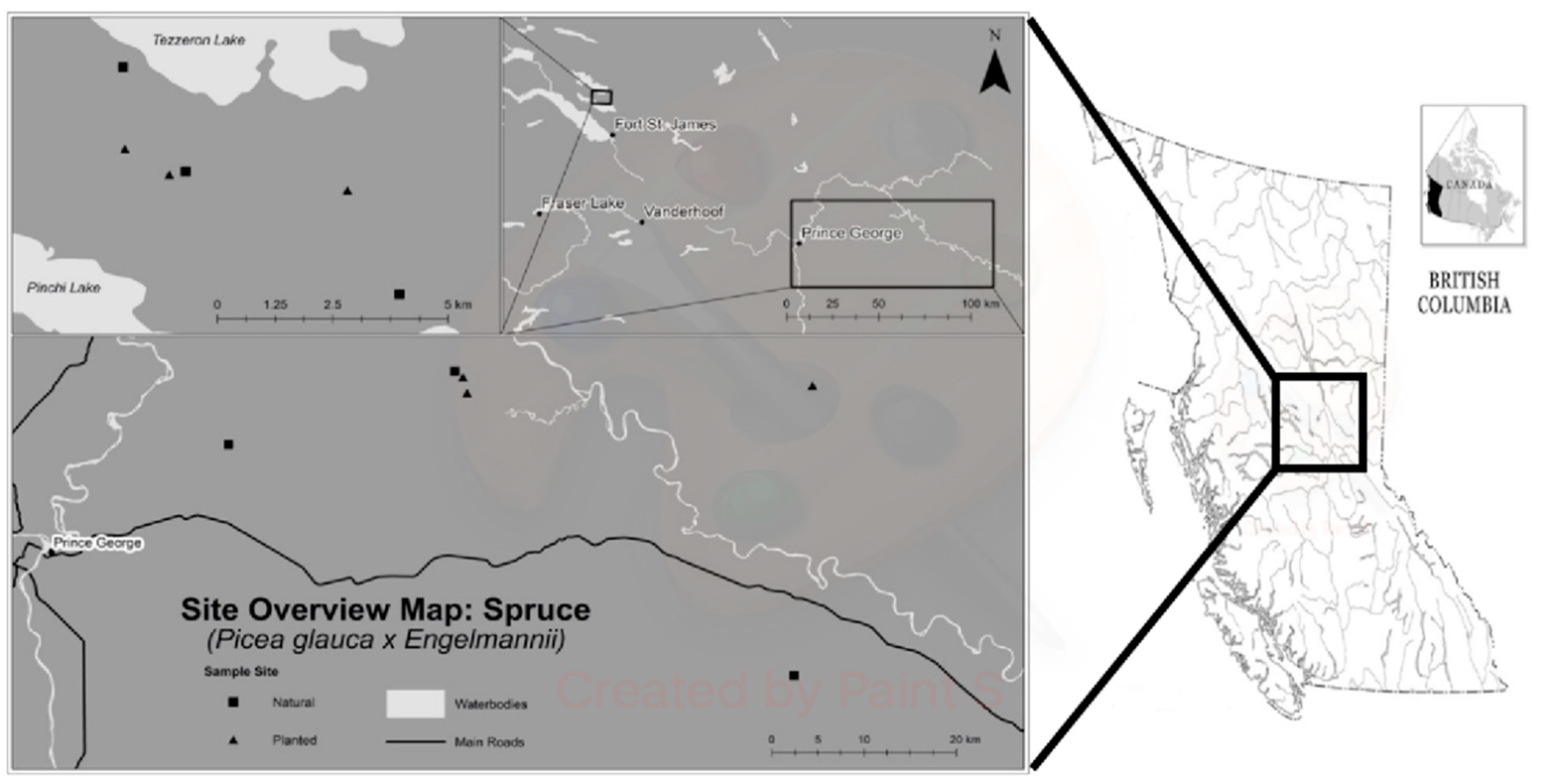



{kind=link}
{kind=link}
{kind=link}
| Geographic Information | Tree Characteristics | |||||||
|---|---|---|---|---|---|---|---|---|
| Site | Latitude | Longitude | Elevation (m) | Slope (%) | Mean Age (Years) | Number of 5 mm Tree Cores | Mean DBH (cm) | |
| FSJ | N1 | 54 38′50.80 | 124 23′35.1 | 768 | <2 | 101 | 40 | 27 |
| N2 | 54 39′46.60 | 124 24′36.6 | 833 | <5 | 119 | 35 | 39 | |
| N3 | 54 36′58.21 | 124 19′05.5 | 727 | 0 | 52 | 34 | 31 | |
| P1 | 54 38′47.50 | 124 23′68.2 | 801 | 0 | 28 | 40 | 18 | |
| P2 | 54 38′48.00 | 124 24′34.5 | 866 | 20 | 31 | 40 | 22 | |
| P3 | 54 38′14.17 | 124 20′05.5 | 802 | 0 | 25 | 36 | 19 | |
| PG | N4 | 54 04′58.90 | 122 01′32.3 | 730 | 0 | 93 | 40 | 41 |
| N5 | 54 46′33.90 | 121 29′14.6 | 1113 | 0 | 145 | 40 | 48 | |
| N6 | 54 01′01.00 | 122 24′54.5 | 707 | 0 | 154 | 40 | 32 | |
| P4 | 54 04′05.90 | 121 26′48.8 | 843 | <5 | 30 | 40 | 26 | |
| P5 | 54 05′19.90 | 122 01′31.8 | 713 | <10 | 30 | 40 | 28 | |
| P6 | 54 04′01.10 | 122 01′09.7 | 689 | 0 | 33 | 40 | 23 | |
| Stand Level | Regional Level | ||||
|---|---|---|---|---|---|
| % C (SD) * | % C (SD) * | % C (SD) | |||
| N1 | 45.34 a (0.87) | P1 | 42.21 a (0.45) | N | 42.23 a (0.24) |
| N2 | 42.38 b (0.22) | P2 | 44.11 b (0.62) | P | 41.89 a (0.73) |
| N3 | 42.06 b (0.59) | P3 | 41.57 a (0.96) | ||
| FSJ | PG | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 | P2 | P3 | N1trunc | N2 | N2trunc | N3 | P4 | P5 | P6 | N5 | N6 | |
| Jan | - | - | - | - | - | - | −0.70 * | - | - | - | 0.66 * | - |
| Mar | - | - | - | - | - | - | −0.64 * | - | - | - | - | - |
| Apr | - | - | - | - | - | - | −0.74 * | - | - | - | - | - |
| Jul | - | - | - | - | - | - | - | −0.82 * | - | −0.87 * | 0.64 ** | - |
| Jul | - | - | - | - | - | −0.82 * | −0.75 ** | - | - | −0.82 * | 0.76 ** | - |
| Aug | - | - | - | - | - | - | −0.66 * | - | - | - | - | - |
| Aug | - | - | - | - | - | −0.78 * | - | - | - | - | 0.64 ** | - |
| Sept | - | - | - | - | - | - | - | −0.87 * | - | - | - | - |
| Sept | 0.98 ** | - | 0.86 ** | - | - | - | - | - | - | - | - | - |
| Nov | - | - | - | - | - | 0.77 * | - | - | - | - | - | - |
| Dec | - | - | - | - | - | - | - | - | - | - | - | - |
| Spring | - | - | - | - | - | - | −0.8 * | - | - | - | - | |
| Summer | - | - | - | - | - | - | −0.68 * | - | - | - | - | - |
| Winter | - | - | - | - | - | - | −0.75 ** | - | - | - | - | - |
| Feb | - | - | 0.84 * | - | - | - | - | - | - | - | - | - |
| Mar | - | - | - | −0.85 * | - | - | - | - | - | - | - | - |
| May | - | 0.77 * | - | - | - | - | - | - | - | - | - | - |
| Jun | - | - | - | - | - | - | - | - | - | - | - | 0.79 * |
| Sept | - | - | - | - | 0.58 ** | 0.87 ** | - | - | - | - | - | - |
| Nov | - | - | - | - | - | - | - | −0.78 * | - | - | - | - |
| Spring | - | - | - | - | - | - | - | - | −0.86 * | - | - | - |
| Winter | - | - | 0.97 ** | 0.9 ** | - | - | - | - | - | - | - | - |
| Month | Site | Carbon (R2) | Pearson’s R (Xmea v Xmod) | p-Value |
|---|---|---|---|---|
| Jan temp | N3 | 0.485 ** | 0.696 | 0.017 |
| Jan temp | N5 | 0.432 ** | 0.657 | 0.006 |
| Mar precip | N1trunc | 0.718 ** | 0.848 | 0.016 |
| Jul temp | N2trunc | 0.669 * | 0.818 | 0.025 |
| Jul temp | N3 | 0.555 ** | 0.745 | 0.009 |
| Aug temp | N5 | 0.574 ** | 0.758 | 0.001 |
| Previous Aug temp | N3 | 0.439 * | 0.663 | 0.026 |
| Spring temp | N3 | 0.639 ** | 0.800 | 0.003 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanusic, A.; Wood, L.J.; Lewis, K. Structural Carbon Allocation and Wood Growth Reflect Climate Variation in Stands of Hybrid White Spruce in Central Interior British Columbia, Canada. Forests 2020, 11, 879. https://doi.org/10.3390/f11080879
Ivanusic A, Wood LJ, Lewis K. Structural Carbon Allocation and Wood Growth Reflect Climate Variation in Stands of Hybrid White Spruce in Central Interior British Columbia, Canada. Forests. 2020; 11(8):879. https://doi.org/10.3390/f11080879
Chicago/Turabian StyleIvanusic, Anastasia, Lisa J. Wood, and Kathy Lewis. 2020. "Structural Carbon Allocation and Wood Growth Reflect Climate Variation in Stands of Hybrid White Spruce in Central Interior British Columbia, Canada" Forests 11, no. 8: 879. https://doi.org/10.3390/f11080879
APA StyleIvanusic, A., Wood, L. J., & Lewis, K. (2020). Structural Carbon Allocation and Wood Growth Reflect Climate Variation in Stands of Hybrid White Spruce in Central Interior British Columbia, Canada. Forests, 11(8), 879. https://doi.org/10.3390/f11080879