Impact of Stand Density and Tree Social Status on Aboveground Biomass Allocation of Scots Pine Pinus sylvestris L.

 ,
,  ,
,  , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
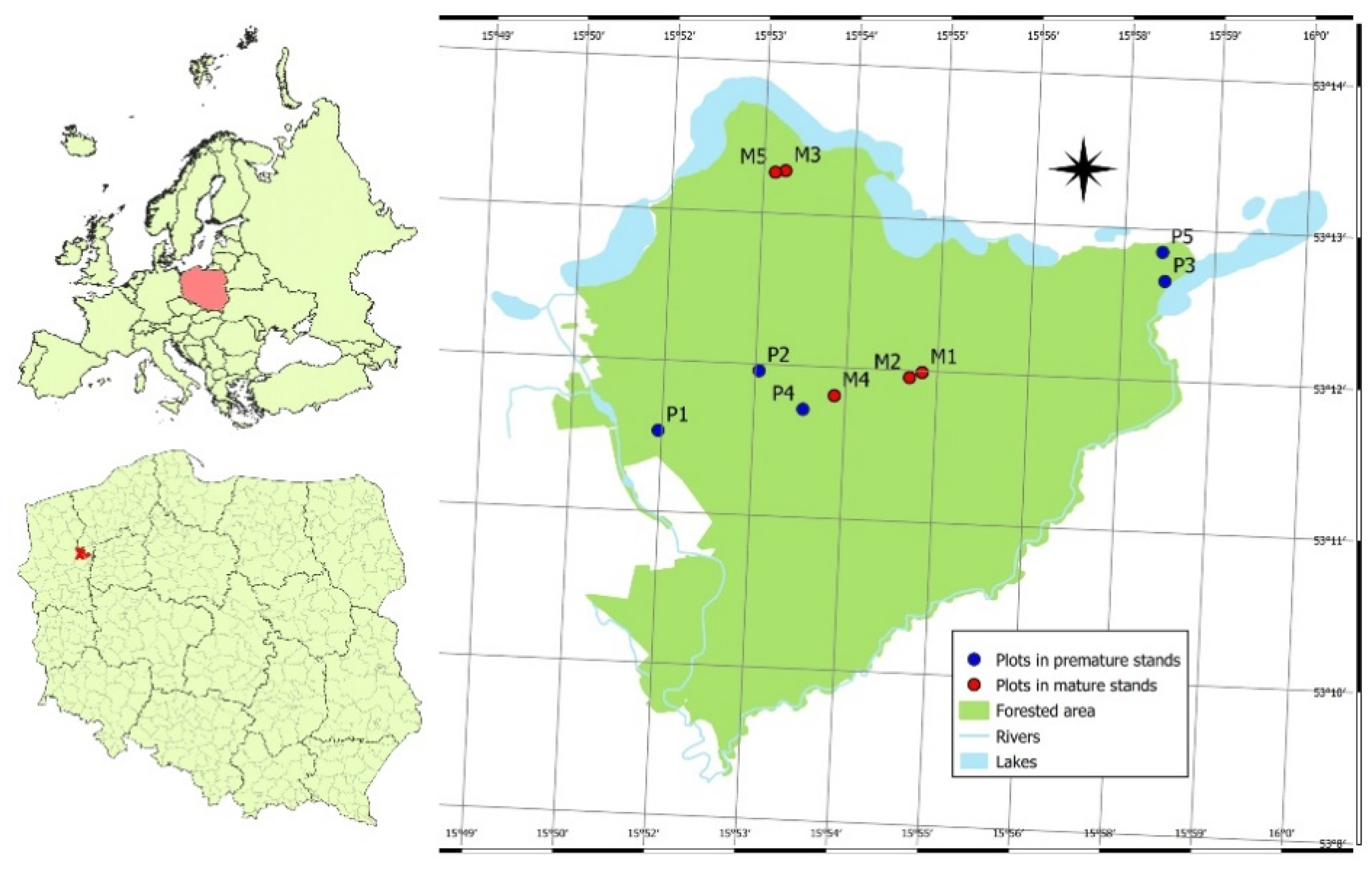
2.1. Study Area
2.2. Selection of Sample Trees
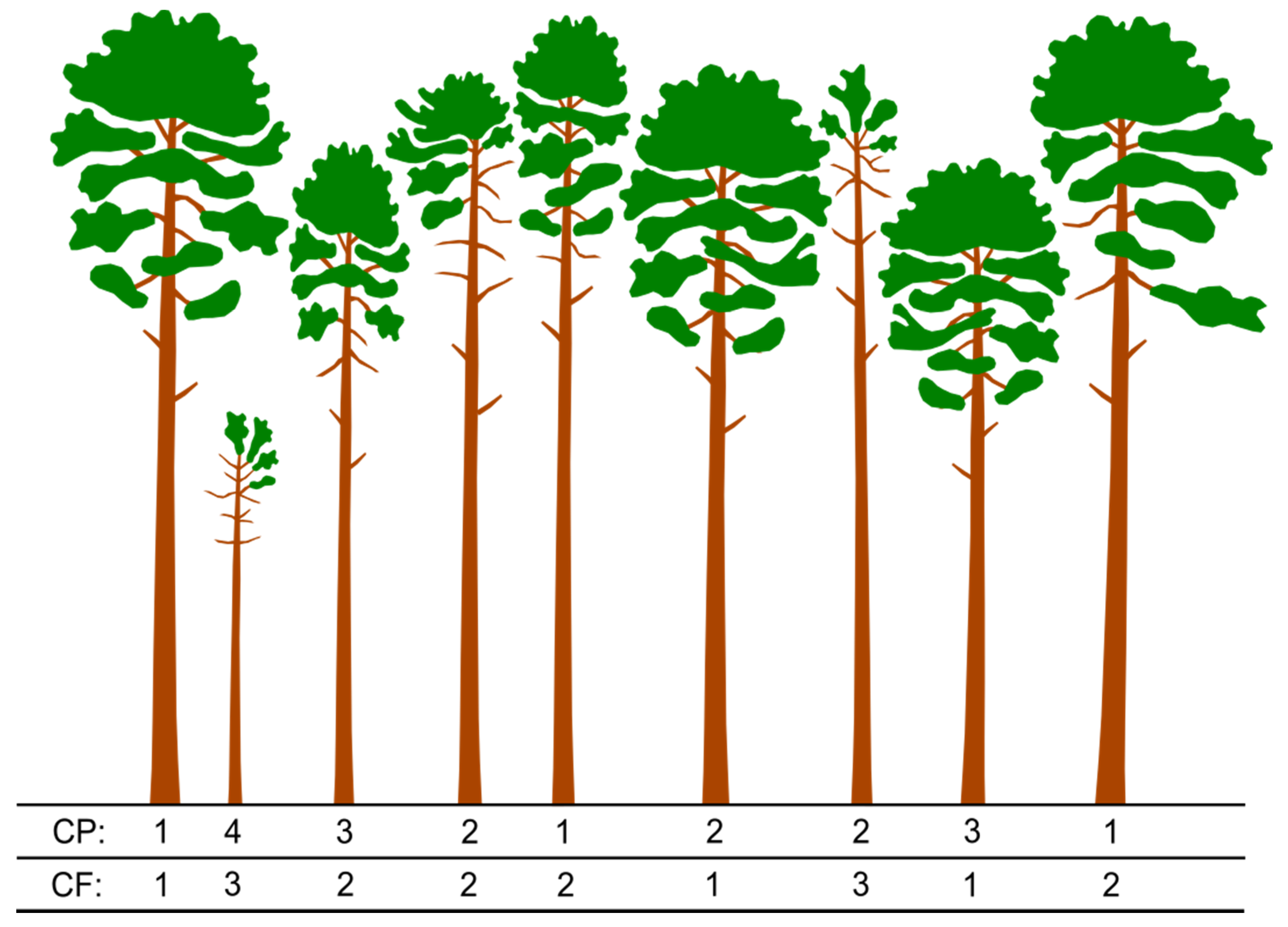
2.3. Tree Classification
2.4. Biomass Assessment
2.5. Statistical Analysis
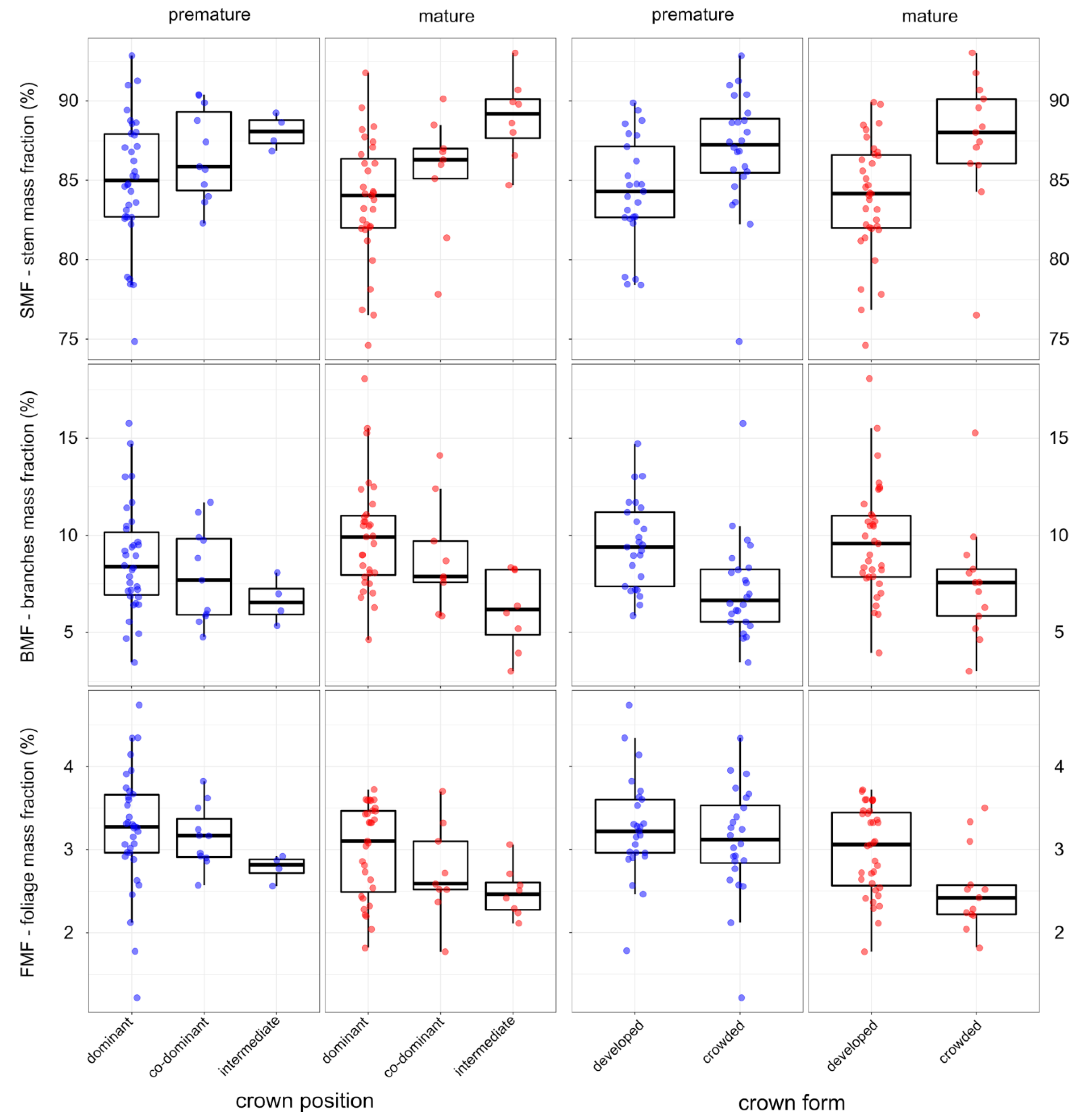
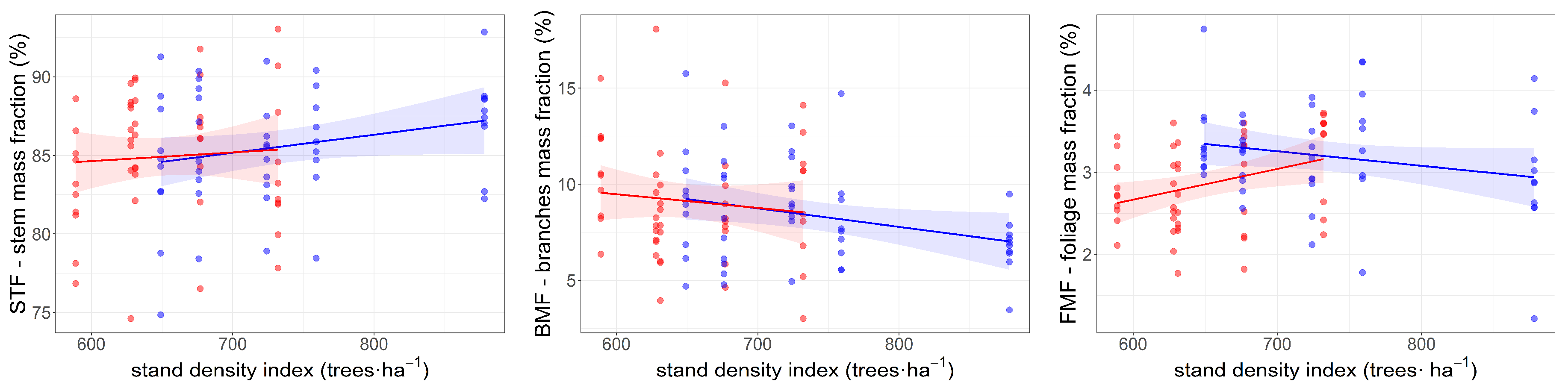
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Laiho, R.; Laine, J. Tree stand biomass and carbon content in an age sequence of drained pine mires in southern Finland. For. Ecol. Manag. 1997, 93, 161–169. [Google Scholar] [CrossRef]
- Tolunay, D. Carbon concentrations of tree components, forest floor and understorey in young Pinus sylvestris stands in north-western Turkey. Scand. J. For. Res. 2009, 24, 394–402. [Google Scholar] [CrossRef]
- Singh, V.; Tewari, A.; Kushwaha, S.P.S.; Dadhwal, V.K. Formulating allometric equations for estimating biomass and carbon stock in small diameter trees. For. Ecol. Manag. 2011, 261, 1945–1949. [Google Scholar] [CrossRef]
- Poorter, H.; Sack, L. Pitfalls and Possibilities in the Analysis of Biomass Allocation Patterns in Plants. Front. Plant Sci. 2012, 3. [Google Scholar] [CrossRef]
- Litton, C.; Raich, J.; Ryan, M. Carbon allocation in forest ecosystems. Glob. Chang. Biol. 2007, 13, 2089–2109. [Google Scholar] [CrossRef]
- Reich, P.A. Root–Shoot Relations: Optimality in Acclimation and Adaptation or the “Emperor’s New Clothes”? In Plant Roots, the Hidden Half, 3rd ed.; Waisel, Y., Eshel, A., Kafkafi, U., Eds.; Marcel Dekker Inc.: New York, NY, USA; Basel, Swterzland, 2002; pp. 205–220. [Google Scholar]
- Bartelink, H.H. A model of dry matter partitioning in trees. Tree Physiol. 1998, 18, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Thornley, J.H.M. A Balanced Quantitative Model for Root: Shoot Ratios in Vegetative Plants. Ann. Bot. 1972, 36, 431–441. [Google Scholar] [CrossRef]
- Cannell, M.G.R.; Dewar, R.C. Carbon allocation in trees: A review of concepts for modelling. Adv. Ecol. Res. 1994, 59–104. [Google Scholar] [CrossRef]
- Mäkelä, A.; Landsberg, J.; Ek, A.R.; Burk, T.E.; Ter-Mikaelian, M.; Agren, G.I.; Oliver, C.D.; Puttonen, P. Process-based models for forest ecosystem management: Current state of the art and challenges for practical implementation. Tree Physiol. 2000, 20, 289–298. [Google Scholar] [CrossRef]
- Bontemps, J.-D.; Bouriaud, O. Predictive approaches to forest site productivity: Recent trends, challenges and future perspectives. For. An Int. J. For. Res. 2013, 87, 109–128. [Google Scholar] [CrossRef]
- Hember, R.A.; Kurz, W.A. Low tree-growth elasticity of forest biomass indicated by an individual-based model. Forests 2018, 9, 21. [Google Scholar] [CrossRef]
- Ruiz-Benito, P.; Vacchiano, G.; Lines, E.R.; Reyer, C.P.O.; Ratcliffe, S.; Morin, X.; Hartig, F.; Mäkelä, A.; Yousefpour, R.; Chaves, J.E.; et al. Available and missing data to model impact of climate change on European forests. Ecol. Model. 2020, 416, 108870. [Google Scholar] [CrossRef]
- Jelonek, T.; Tomczak, A.; Karaszewski, Z.; Jakubowski, M.; Arasimowicz-Jelonek, M.; Grzywiński, W.; Kopaczyk, J.; Klimek, K. The biomechanical formation of trees. Drewno 2019, 62, 5–22. [Google Scholar] [CrossRef]
- Trouvé, R.; Bontemps, J.-D.; Seynave, I.; Collet, C.; Lebourgeois, F. Stand density, tree social status and water stress influence allocation in height and diameter growth of Quercus petraea (Liebl.). Tree Physiol. 2015, 35, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Grote, R.; Gessler, A.; Hommel, R.; Poschenrieder, W.; Priesack, E. Importance of tree height and social position for drought-related stress on tree growth and mortality. Trees 2016, 30, 1467–1482. [Google Scholar] [CrossRef]
- Tavankar, F.; Picchio, R.; Lo Monaco, A.; Nikooy, M.; Venanzi, R.; Bonyad, A.E. Wound healing rate in oriental beech trees following logging damage. Drewno 2019, 62, 5–22. [Google Scholar] [CrossRef]
- Perry, D.A. The competition process in forest stands. In Attributes of Trees As Crop Plants; Cannell, M.G.R., Jackson, J.E., Eds.; Institute of Terrestial Ecology: Abbots Ripton, Hunts, UK, 1985; pp. 481–506. [Google Scholar]
- Hemery, G.E.; Savill, P.S.; Pryor, S.N. Applications of the crown diameter–stem diameter relationship for different species of broadleaved trees. For. Ecol. Manag. 2005, 215, 285–294. [Google Scholar] [CrossRef]
- Lundqvist, L.; Elfving, B. Influence of biomechanics and growing space on tree growth in young Pinus sylvestris stands. For. Ecol. Manag. 2010, 260, 2143–2147. [Google Scholar] [CrossRef]
- Kaźmierczak, K. Selected measures of the growth space of a single tree in maturing pine stand. Sylwan 2009, 153, 298–303. [Google Scholar] [CrossRef]
- Turski, M.; Jaszczak, R.; Deus, R. Selected features of tree crowns and their relationship with the dbh and height in pine tree-stands of different age classes. Sylwan 2012, 156, 369–378. [Google Scholar] [CrossRef]
- Bielinis, E.; Kwiatkowski, J.; Boiko, S. Identification of Pinus sylvestris clones with the highest and lowest allelopathic potentials. Balt. For. 2019, 25, 52–58. [Google Scholar] [CrossRef]
- Kojola, S.; Ahtikoski, A.; Hökkä, H.; Penttilä, T. Profitability of alternative management regimes in Scots pine stands on drained peatlands. Eur. J. For. Res. 2012, 131, 413–426. [Google Scholar] [CrossRef]
- Giuggiola, A.; Bugmann, H.; Zingg, A.; Dobbertin, M.; Rigling, A. Reduction of stand density increases drought resistance in xeric Scots pine forests. For. Ecol. Manag. 2013, 310, 827–835. [Google Scholar] [CrossRef]
- Tahvonen, O.; Pihlainen, S.; Niinimäki, S. On the economics of optimal timber production in boreal Scots pine stands. Can. J. For. Res. 2013, 43, 719–730. [Google Scholar] [CrossRef]
- Curtis, R.O.; Marshall, D.D.; Bell, J.F. LOGS: A pioneering example of silvicultural research in coast Douglas-fir. J. For. 1997, 95, 19–25. [Google Scholar]
- Kuliešis, A.; Saladis, J.; Kuliešis, A.A. Development and productivity of young Scots pine stands by regulating density. Balt. For. 2010, 16, 235–246. [Google Scholar]
- Nilsson, U.; Agestam, E.; Ekö, P.-M.; Elfving, B.; Fahlvik, N.; Johansson, U.; Karlsson, K.; Lundmark, T.; Wallentin, C. Thinning of Scots pine and Norway spruce monocultures in Sweden. Studia For. Suec. 2010, 219, 1–46. [Google Scholar]
- Valinger, E.; Elfving, B.; Mörling, T. Twelve-year growth response of Scots pine to thinning and nitrogen fertilisation. For. Ecol. Manag. 2000, 134, 45–53. [Google Scholar] [CrossRef]
- Mäkinen, H.; Isomäki, A. Thinning intensity and long-term changes in increment and stem form of Scots pine trees. For. Ecol. Manag. 2004, 203, 21–34. [Google Scholar] [CrossRef]
- Mäkinen, H.; Hynynen, J.; Isomäki, A. Intensive management of Scots pine stands in southern Finland: First empirical results and simulated further development. For. Ecol. Manag. 2005, 215, 37–50. [Google Scholar] [CrossRef]
- Zhang, J.; Oliver, W.W.; Ritchie, M.W. Effect of stand densities on stand dynamics in white fir (Abies concolor) forests in northeast California, USA. For. Ecol. Manag. 2007, 244, 50–59. [Google Scholar] [CrossRef]
- Del Río, M.; Calama, R.; Cañellas, I.; Roig, S.; Montero, G. Thinning intensity and growth response in SW-European Scots pine stands. Ann. For. Sci. 2008, 65. [Google Scholar] [CrossRef]
- Gizachew, B.; Brunner, A. Density-growth relationships in thinned and unthinned Norway spruce and Scots pine stands in Norway. Scand. J. For. Res. 2011, 26, 543–554. [Google Scholar] [CrossRef]
- Routa, J.; Kellomäki, S.; Strandman, H. Effects of forest management on total biomass production and CO2 emissions from use of energy biomass of Norway spruce and Scots pine. Bioenergy Res. 2012, 5, 733–747. [Google Scholar] [CrossRef]
- Bembenek, M.; Karaszewski, Z.; Kondracki, K.; Łacka, A.; Mederski, P.S.; Skorupski, M.; Strzeliński, P.; Sułkowski, S.; Węgiel, A. Value of merchantable timber in Scots pine stands of different densities. Drewno 2014, 57, 133–142. [Google Scholar] [CrossRef]
- Eriksson, E. Thinning operations and their impact on biomass production in stands of Norway spruce and Scots pine. Biomass Bioenergy 2006, 30, 848–854. [Google Scholar] [CrossRef]
- Węgiel, A.; Bembenek, M.; Łacka, A.; Mederski, P.S. Relationship between stand density and value of timber assortments: A case study for Scots pine stands in north-western Poland. New Zealand J. For. Sci. 2018, 48, 1–9. [Google Scholar] [CrossRef]
- Lehtonen, A.; Mäkipää, R.; Heikkinen, J.; Sievänen, R.; Liski, J. Biomass expansion factors (BEFs) for Scots pine, Norway spruce and birch according to stand age for boreal forests. For. Ecol. Manag. 2004, 188, 211–224. [Google Scholar] [CrossRef]
- Jelonek, T.; Pazdrowski, W.; Tomczak, A.; Szaban, J. The effect of social position of tree in the stand and site on wood quality of Scots pine (Pinus sylvestris L.). Electron. J. Pol. Agric. Univ. 2008, 11, 10. [Google Scholar]
- Bajrakatri, A.; Pimenta, R.; Pinto, T.; Miranda, I.; Knapic, S.; Nunes, L.; Pereira, H. Stem quality of Quercus cerris trees from Kosovo for the sawmilling industry. Drewno 2018, 61, 57–69. [Google Scholar] [CrossRef]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Funct. Plant Biol. 2000, 27, 1191. [Google Scholar] [CrossRef]
- Nilsson, U.; Albrektson, A. Productivity of needles and allocation of growth in young Scots pine trees of different competitive status. For. Ecol. Manag. 1993, 62, 173–187. [Google Scholar] [CrossRef]
- Mäkinen, H. Effect of intertree competition on biomass production of Pinus sylvestris (L.) half-sib families. For. Ecol. Manag. 1996, 86, 105–112. [Google Scholar] [CrossRef]
- Vanninen, P.; Mäkelä, A. Needle and stem wood production in Scots pine (Pinus sylvestris) trees of different age, size and competitive status. Tree Physiol. 2000, 20, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Vanninen, P. Allocation of above-ground growth in Pinus sylvestris–impacts of tree size and competition. Silva Fenn. 2004, 38. [Google Scholar] [CrossRef]
- Ochał, W.; Grabczyński, S.; Orzeł, S.; Wertz, B.; Socha, J. Aboveground biomass allocation in Scots pines of different biosocial positions in the stand. Sylwan 2013, 157, 737–746. [Google Scholar] [CrossRef]
- Ziemblińska, K.; Urbaniak, M.; Chojnicki, B.H.; Black, T.A.; Niu, S.; Olejnik, J. Net ecosystem productivity and its environmental controls in a mature Scots pine stand in north-western Poland. Agric. For. Meteorol. 2016, 228–229, 60–72. [Google Scholar] [CrossRef]
- Reineke, L.H. Perfecting a stand-density index for even-aged forests. J. Agric. Res. Wash. 1933, 46, 627–638. [Google Scholar]
- Siipilehto, J. A comparison of two parameter prediction methods for stand structure in Finland. Silva Fenn. 2000, 34, 331–349. [Google Scholar] [CrossRef]
- Schädelin, W. Die Durchforstung als Auslese- und Veredelungsbetrieb Höchster Wertleistung; Verlag Paul Haupt, Bern-Leipzig: Bern, Switzerland, 1936. [Google Scholar]
- Raptis, D.; Kazana, V.; Kazaklis, A.; Stamatiou, C. A crown width-diameter model for natural even-aged black pine forest management. Forests 2018, 9, 610. [Google Scholar] [CrossRef]
- Sharma, R.P.; Bílek, L.; Vacek, Z.; Vacek, S. Modelling crown width–diameter relationship for Scots pine in the central Europe. Trees 2017, 31, 1875–1889. [Google Scholar] [CrossRef]
- Sharma, R.P.; Vacek, Z.; Vacek, S. Individual tree crown width models for Norway spruce and European beech in Czech Republic. For. Ecol. Manag. 2016, 366, 208–220. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 5 June 2020).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Lüdecke, D. sjPlot: Data Visualization for Statistics in Social Science. R Package Version 2.8.4. Available online: https://CRAN.R-project.org/package=sjPlot (accessed on 5 June 2020).
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Statistics for Biology and Health; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Zeide, B. Optimal stand density: A solution. Can. J. For. Res. 2004, 34, 846–854. [Google Scholar] [CrossRef]
- MacDonald, E.; Hubert, J. A review of the effects of silviculture on timber quality of Sitka spruce. For. Inst. For. Great Br. 2002, 75, 107–138. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef] [PubMed]
- Bartelink, H.H. Allometric relationships on biomass and needle area of Douglas-fir. For. Ecol. Manag. 1996, 86, 193–203. [Google Scholar] [CrossRef]
- Vanninen, P.; Mäkelä, A. Carbon budget for Scots pine trees: Effects of size, competition and site fertility on growth allocation and production. Tree Physiol. 2005, 25, 17–30. [Google Scholar] [CrossRef]
- Naidu, S.L.; DeLucia, E.H.; Thomas, R.B. Contrasting patterns of biomass allocation in dominant and suppressed loblolly pine. Can. J. For. Res. 1998, 28, 1116–1124. [Google Scholar] [CrossRef]
- Kellomäki, S. Effect of the within-stand light conditions on the share of stem, branch and needle growth in a twenty-year-old Scots pine stand. Silva Fenn. 1981, 15, 130–139. [Google Scholar] [CrossRef][Green Version]
- Ratajczak, E.; Szostak, A.; Bidzińska, G.; Leszczyszyn, E. Market in wood by-products in Poland and their flows in the wood sector. Drewno 2018, 61, 5–20. [Google Scholar] [CrossRef]
- Bembenek, M.; Giefing, D.E.; Jelonek, T.; Karaszewski, Z.; Kruszyk, R.; Tomczak, A.; Woszczyk, M.; Mederski, P.S. Carbon content in juvenile and mature wood of Scots pine (Pinus sylvestris L.). Balt. For. 2015, 21, 279–284. [Google Scholar]
- Wegiel, A.; Polowy, K. Aboveground carbon content and storage in mature Scots pine stands of different densities. Forests 2020, 11, 240. [Google Scholar] [CrossRef]
- Hasenauer, H.; Monserud, R.A. A crown ratio model for Austrian forests. For. Ecol. Manag. 1996, 84, 49–60. [Google Scholar] [CrossRef]
- Biging, G.S.; Dobbertin, M. Evaluation of competition indices in individual tree growth models. For. Sci. 1995, 41, 360–377. [Google Scholar] [CrossRef]
- Stadt, K.J.; Huston, C.; Coates, K.D.; Feng, Z.; Dale, M.R.T.; Lieffers, V.J. Evaluation of competition and light estimation indices for predicting diameter growth in mature boreal mixed forests. Ann. For. Sci. 2007, 64, 477–490. [Google Scholar] [CrossRef]
- Monserud, R.A.; Sterba, H. Modeling individual tree mortality for Austrian forest species. For. Ecol. Manag. 1999, 113, 109–123. [Google Scholar] [CrossRef]
- Oker-Blom, P.; Pukkala, T.; Kuuluvainen, T. Relationship between radiation interception and photosynthesis in forest canopies: Effect of stand structure and latitude. Ecol. Model. 1989, 49, 73–87. [Google Scholar] [CrossRef]
- Tahvanainen, T.; Forss, E. Individual tree models for the crown biomass distribution of Scots pine, Norway spruce and birch in Finland. For. Ecol. Manag. 2008, 255, 455–467. [Google Scholar] [CrossRef]
- Yang, B.; Xue, W.; Yu, S.; Zhou, J.; Zhang, W. Effects of stand age on biomass allocation and allometry of Quercus acutissima in the Central Loess Plateau of China. Forests 2019, 10, 41. [Google Scholar] [CrossRef]
- Picchio, R.; Venanzi, R.; Latterini, F.; Marchi, E.; Laschi, A.; Lo Monaco, A. Corsican pine (Pinus laricio Poiret) stand management: Medium and long lasting effects of thinning on biomass growth. Forests 2018, 9, 257. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Plot | Age (Years) | SDI (Trees·ha−1) | Mean DBH (cm) | Mean Height (m) | Basal Area (m2·ha−1) | Standing Stock (m3·ha−1) |
|---|---|---|---|---|---|---|
| P1 | 59 | 724 | 23.4 | 21.6 | 31.2 | 312 |
| P2 | 57 | 759 | 20.6 | 21.4 | 31.0 | 310 |
| P3 | 55 | 878 | 19.2 | 20.5 | 33.8 | 325 |
| P4 | 53 | 676 | 16.6 | 18.3 | 30.6 | 265 |
| P5 | 55 | 649 | 18.5 | 20.0 | 38.5 | 364 |
| M1 | 82 | 628 | 28.2 | 22.9 | 30.5 | 319 |
| M2 | 82 | 677 | 25.7 | 20.8 | 31.5 | 301 |
| M3 | 82 | 589 | 23.6 | 19.6 | 30.1 | 275 |
| M4 | 82 | 631 | 23.9 | 20.1 | 35.7 | 334 |
| M5 | 82 | 732 | 21.8 | 19.3 | 31.5 | 286 |
| Stand’s Stage of Development | Tree’s Social Status | N | DBH (cm) | Height (m) | Crown Length (m) | Volume (m3) | |
|---|---|---|---|---|---|---|---|
| premature | crown position | dominant | 35 | 21.7 ± 0.7 | 20.8 ± 0.3 | 7.0 ± 0.3 | 0.390 ± 0.029 |
| co-dominant | 11 | 16.5 ± 0.9 | 19.2 ± 0.5 | 5.7 ± 0.3 | 0.219 ± 0.033 | ||
| intermediate | 4 | 14.6 ± 1.7 | 16.3 ± 1.0 | 4.1 ± 0.6 | 0.134 ± 0.035 | ||
| crown form | developed | 25 | 22.2 ± 0.8 | 20.7 ± 0.3 | 7.0 ± 0.3 | 0.406 ± 0.033 | |
| crowded | 24 | 17.6 ± 0.9 | 19.4 ± 0.4 | 5.8 ± 0.4 | 0.253 ± 0.033 | ||
| dying | 1 | 22.0 | 20.6 | 9.9 | 0.395 | ||
| mature | crown position | dominant | 32 | 26.7 ± 0.9 | 21.6 ± 0.3 | 7.6 ± 0.4 | 0.589 ± 0.042 |
| co-dominant | 9 | 21.5 ± 1.0 | 20.5 ± 0.6 | 6.4 ± 0.4 | 0.387 ± 0.051 | ||
| intermediate | 8 | 21.7 ± 1.5 | 20.2 ± 0.9 | 5.8 ± 0.8 | 0.339 ± 0.060 | ||
| overtopped | 1 | 16.5 | 18.7 | 5.7 | 0.182 | ||
| crown form | developed | 35 | 26.2 ± 0.8 | 21.8 ± 0.2 | 7.7 ± 0.3 | 0.574 ± 0.038 | |
| crowded | 14 | 20.5 ± 1.1 | 19.3 ± 0.4 | 5.4 ± 0.5 | 0.303 ± 0.040 | ||
| dying | 1 | 33.7 | 21.3 | 7.2 | 0.886 | ||
| Stand’s Stage of Development | Tree’s Social Status | Stem | Living Branches | Foliage | Dead Branches | Cones | Total | |
|---|---|---|---|---|---|---|---|---|
| pre-mature | crown position | dominant | 165.9 ± 11.9 | 19.8 ± 2.5 | 6.7 ± 0.6 | 5.8 ± 0.9 | 0.8 ± 0.2 | 199.0 ± 15.7 |
| co-dominant | 96.7 ± 13.9 | 9.4 ± 2.1 | 3.6 ± 0.5 | 2.3 ± 0.4 | 0.2 ± 0.1 | 112.1 ± 16.5 | ||
| intermediate | 55.8 ± 13.1 | 4.0 ± 0.8 | 1.7 ± 0.4 | 1.6 ± 0.4 | 0 | 63.2 ± 14.6 | ||
| crown form | developed | 171.8 ± 13.8 | 20.6 ± 2.4 | 6.9 ± 0.7 | 5.6 ± 0.9 | 0.9 ± 0.3 | 205.8 ± 17.5 | |
| crowded | 109.4 ± 13.5 | 10.8 ± 2.9 | 4.1 ± 0.7 | 3.7 ± 1.0 | 0.4 ± 0.1 | 128.4 ± 17.9 | ||
| dying | 171.4 | 36.4 | 10.9 | 6.4 | 0.2 | 225.2 | ||
| mature | crown position | dominant | 242.9 ± 16.8 | 31.8 ± 3.5 | 8.9 ± 0.7 | 10.3 ± 1.3 | 0.7 ± 0.2 | 294.6 ± 21.7 |
| co-dominant | 160.9 ± 20.6 | 17.4 ± 3.1 | 5.1 ± 0.7 | 5.1 ± 0.8 | 0.6 ± 0.3 | 189.1 ± 24.4 | ||
| intermediate | 139.7 ± 23.8 | 10.5 ± 2.6 | 4.0 ± 0.8 | 3.6 ± 0.8 | 0.5 ± 0.2 | 158.3 ± 27.8 | ||
| overtopped | 77.2 | 6.4 | 2.1 | 2.3 | 0 | 88.0 | ||
| crown form | developed | 237.5 ± 14.9 | 29.5 ± 3.0 | 8.5 ± 0.6 | 9.3 ± 1.0 | 0.9 ± 0.2 | 285.7 ± 19.1 | |
| crowded | 125.6 ± 15.9 | 12.4 ± 3.3 | 3.8 ± 0.7 | 3.8 ± 1.0 | 0.1 ± 0 | 145.7 ± 20.3 | ||
| dying | 345.0 | 59.6 | 12.3 | 26.5 | 0.7 | 444.1 | ||
| SMF | BMF | FMF | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Fixed Effects | β | CI | t | β | CI | t | β | CI | t |
| (Intercept) | 80.36 | 74.03 ÷ 86.69 | 24.88 * | 16.03 | 10.28 ÷ 21.78 | 5.46 * | 3.59 | 2.18 ÷ 5.00 | 5.00 * |
| co-dominant CP | 1.12 | −0.32 ÷ 2.55 | 1.53 | −0.94 | −2.25 ÷ 0.36 | −1.42 | −0.16 | −0.46 ÷ 0.13 | −1.09 |
| intermediate CP | 3.39 | 1.59 ÷ 5.19 | 3.69 * | −2.94 | −4.57 ÷ −1.30 | −3.52 * | −0.44 | −0.81 ÷ −0.07 | −2.34 * |
| crowded CF | 1.94 | 0.70 ÷ 3.18 | 3.06 * | −1.69 | −2.82 ÷ −0.56 | −2.94 * | −0.25 | −0.51 ÷ −0.01 | −1.91 * |
| mature stands | 0.45 | −0.89 ÷ 1.80 | 0.66 | −0.09 | −1.31 ÷ 1.13 | −0.14 | −0.37 | −0.67 ÷ −0.07 | −2.42 * |
| SDI | 0.0084 | −0.0001 ÷ 0.0170 | 1.93 | −0.0083 | −0.0161 ÷ −0.0005 | −2.08 * | −0.0001 | −0.0021 ÷ 0.0018 | −0.15 |
| Random Effects | |||||||||
| σ2 | 7.96 | 6.57 | 0.33 | ||||||
| τ00, plot | 0.13 | 0.15 | 0.01 | ||||||
| ICC plot | 0.02 | 0.02 | 0.02 | ||||||
| N plot | 10 | 10 | 10 | ||||||
| observations | 97 | 97 | 97 | ||||||
| marginal-R2/conditional-R2 | 0.275/0.288 | 0.273/0.292 | 0.176/0.192 | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wertz, B.; Bembenek, M.; Karaszewski, Z.; Ochał, W.; Skorupski, M.; Strzeliński, P.; Węgiel, A.; Mederski, P.S. Impact of Stand Density and Tree Social Status on Aboveground Biomass Allocation of Scots Pine Pinus sylvestris L. Forests 2020, 11, 765. https://doi.org/10.3390/f11070765
Wertz B, Bembenek M, Karaszewski Z, Ochał W, Skorupski M, Strzeliński P, Węgiel A, Mederski PS. Impact of Stand Density and Tree Social Status on Aboveground Biomass Allocation of Scots Pine Pinus sylvestris L. Forests. 2020; 11(7):765. https://doi.org/10.3390/f11070765
Chicago/Turabian StyleWertz, Bogdan, Mariusz Bembenek, Zbigniew Karaszewski, Wojciech Ochał, Maciej Skorupski, Paweł Strzeliński, Andrzej Węgiel, and Piotr S. Mederski. 2020. "Impact of Stand Density and Tree Social Status on Aboveground Biomass Allocation of Scots Pine Pinus sylvestris L." Forests 11, no. 7: 765. https://doi.org/10.3390/f11070765
APA StyleWertz, B., Bembenek, M., Karaszewski, Z., Ochał, W., Skorupski, M., Strzeliński, P., Węgiel, A., & Mederski, P. S. (2020). Impact of Stand Density and Tree Social Status on Aboveground Biomass Allocation of Scots Pine Pinus sylvestris L. Forests, 11(7), 765. https://doi.org/10.3390/f11070765