Natural Regeneration Following Partial and Clear-Cut Harvesting in Mature Aspen-Jack Pine Stands in Eastern Canada


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Stand Characteristics and Silvicultural Treatments
2.3. Tree and Natural Regeneration Survey
2.4. On and Off Trail Regeneration Survey
2.5. Statistical Analyses
3. Results
3.1. Stand Basal Area Prior to and After Harvesting
3.2. Regeneration Abundance 7–19 Years after PC and 13–19 Years after CPRS
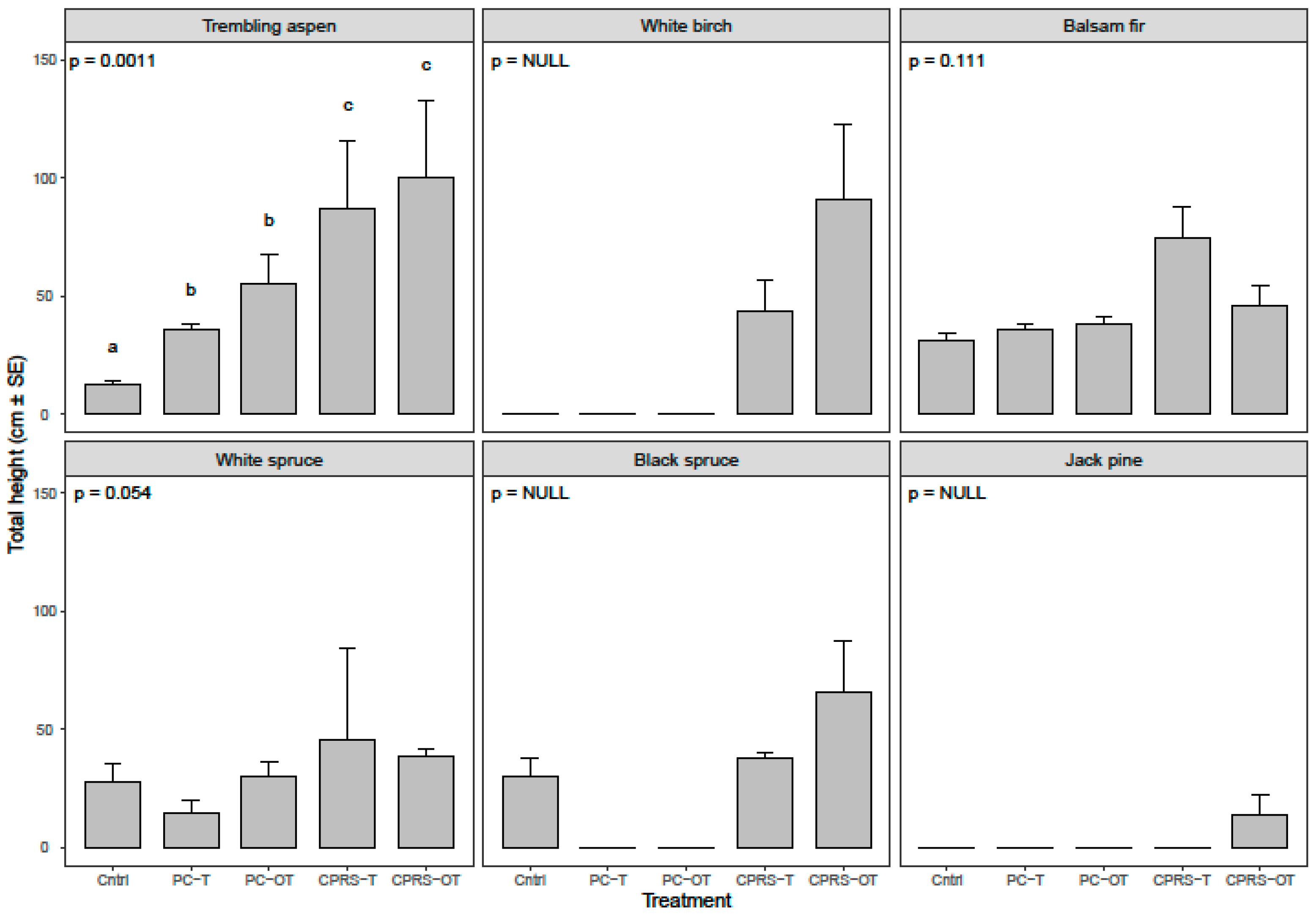
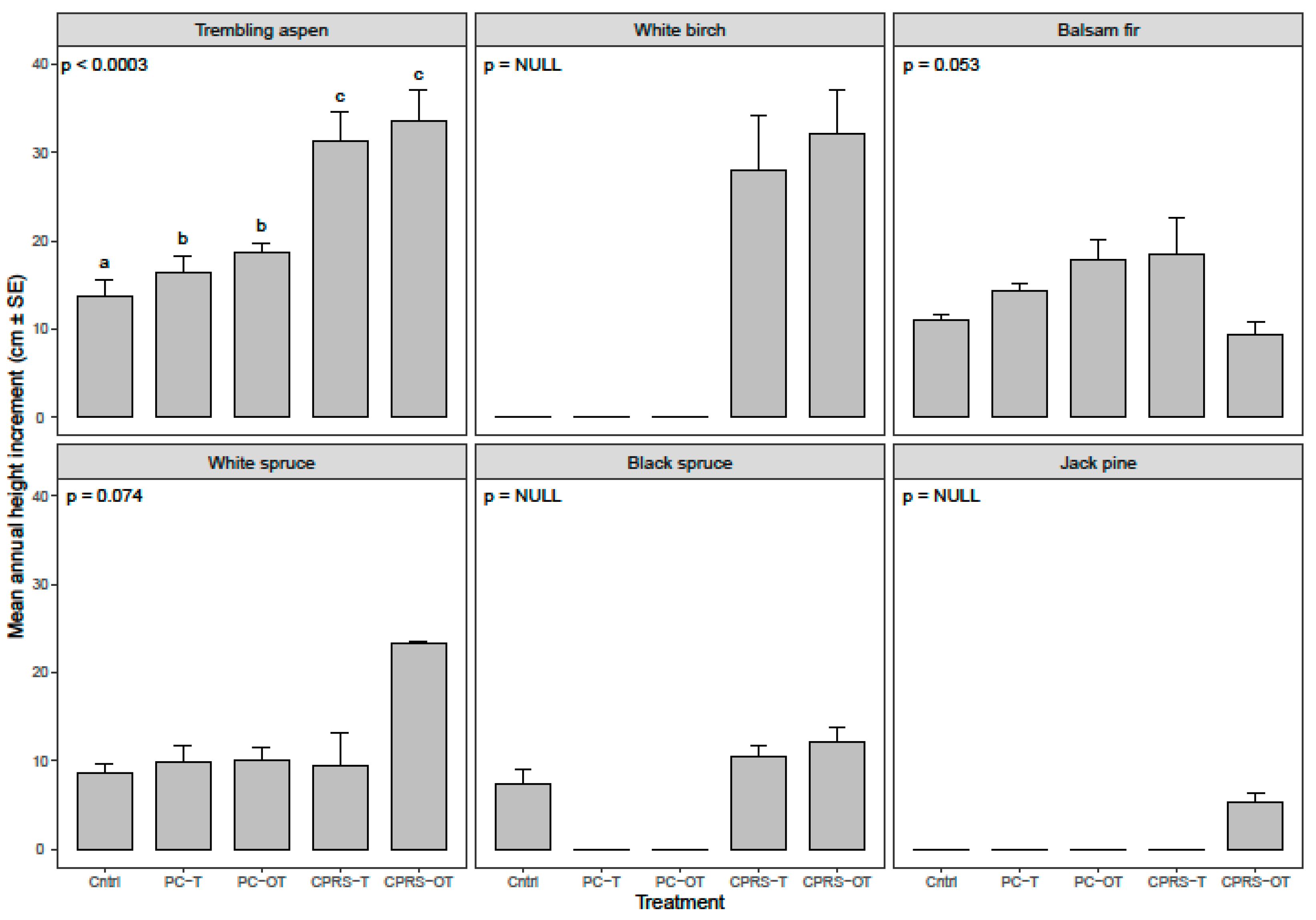
3.3. Regeneration Abundance and Growth on and off Machinery Trails
3.4. Influence of Basal Area and Ground Cover on Seedling Abundance
4. Discussion
4.1. Stand Merchantable Basal Area Proportion 7–19 Years after PC and 13–19 Years after CPRS
4.2. Effects of Partial Cutting on Regeneration Abundance of Conifers (H1) and Aspen (H2)
4.3. Effect of Position Relative to Machinery Trails on Density and Height of Conifers (H3, H4) and Trembling Aspen (H5, H6)
4.4. Influence of Basal Area and Germination Substrates on Seedling Abundance
5. Conclusions and Silvicultural Implications
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bergeron, Y.; Chen, H.Y.H.; Kenkel, N.C.; Leduc, A.L.; Macdonald, S.E. Boreal mixedwood stand dynamics: Ecological processes underlying multiple pathways. For. Chron. 2014, 90, 202–213. [Google Scholar] [CrossRef]
- Gauthier, S.; Leduc, A.; Harvey, B.; Bergeron, Y. Les perturbations naturelles et la diversité écosystémique. Nat. Can. 2001, 125, 10–17. [Google Scholar]
- Chen, H.Y.H.; Popadiouk, R.V. Dynamics of North American boreal mixedwoods. Environ. Rev. 2002, 10, 137–166. [Google Scholar] [CrossRef]
- Bergeron, Y.; Gauthier, S.; Flannigan, M.; Kafka, V. Fire regimes at the transition between mixedwood and coniferous boreal forest in northwestern Quebec. Ecology 2004, 85, 1916–1932. [Google Scholar] [CrossRef]
- Bergeron, Y.; Gauthier, S.; Kafka, V.; Lefort, P.; Lesieur, D. Natural fire frequency for the eastern Canadian boreal forest: Consequences for sustainable forestry. Can. J. For. Res. 2001, 31, 384–391. [Google Scholar] [CrossRef]
- Bergeron, Y.; Leduc, A.; Harvey, B.; Gauthier, S. Stratégies d’aménagement forestier qui s’inspirent de la dynamique des perturbations naturelles: Considérations à l’échelle du peuplement et de la forêt. For. Chron. 1999, 75, 55–61. [Google Scholar] [CrossRef]
- Bergeron, Y.; Dubue, M. Succession in the southern part of the Canadian boreal forest. Vegetatio 1988, 79, 51–63. [Google Scholar] [CrossRef]
- MacDonald, G.B. The case for boreal mixedwood management: An Ontario perspective. For. Chron. 1995, 71, 725–734. [Google Scholar] [CrossRef]
- Bergeron, Y.; Harvey, B. Basing silviculture on natural ecosystem dynamics: An approach applied to the southern boreal mixedwood forest of Quebec. For. Ecol. Manag. 1997, 92, 235–242. [Google Scholar] [CrossRef]
- Bergeron, Y. Species and stand dynamics in the mixed woods of Quebec’s southern boreal forest. Ecology 2000, 81, 1500–1516. [Google Scholar] [CrossRef]
- Légaré, S.; Bergeron, Y.; Leduc, A.; Paré, D. Comparison of the understory vegetation in boreal forest types of southwest Quebec. Can. J. Bot. 2001, 79, 1019–1027. [Google Scholar] [CrossRef]
- Laquerre, S.; Leduc, A.; Harvey, B.D. Augmentation du couvert en peuplier faux-tremble dans les pessières noires du nord-ouest du Québec après coupe totale. Écoscience 2009, 16, 483–491. [Google Scholar] [CrossRef]
- Prévost, M.; Pothier, D. Partial cuts in a trembling aspen—conifer stand: Effects on microenvironmental conditions and regeneration dynamics. Can. J. For. Res. 2003, 33, 1–15. [Google Scholar] [CrossRef]
- Brais, S.; Harvey, B.D.; Bergeron, Y.; Messier, C.; Greene, D.; Belleau, A.; Paré, D. Testing forest ecosystem management in boreal mixedwoods of northwestern Quebec: Initial response of aspen stands to different levels of harvesting. Can. J. For. Res. 2004, 34, 431–446. [Google Scholar] [CrossRef]
- Brais, S.; Work, T.; Robert, É.; O’Connor, C.; Strukelj, M.; Bose, A.; Celentano, D.; Harvey, B. Ecosystem responses to partial harvesting in eastern boreal mixedwood stands. Forests 2013, 4, 364–385. [Google Scholar] [CrossRef]
- Bose, A.K.; Harvey, B.D.; Brais, S. Does partial harvesting promote old-growth attributes of boreal mixedwood trembling aspen (Populus tremuloides Michx.) stands? For. Ecol. Manag. 2015, 353, 173–186. [Google Scholar] [CrossRef]
- Laquerre, S.; Harvey, B.D.; Leduc, A. Spatial analysis of response of trembling aspen patches to clearcutting in black spruce-dominated stands. For. Chron. 2011, 87, 77–85. [Google Scholar] [CrossRef]
- Sims, R.A.; Kershaw, H.M.; Wickware, G.M. The Autecology of Major Tree Species in the North Central Region of Ontario; COFRDA Report 3302; Canada-Ontario Forest Resource Development Agreement, Canadian Forestry Service: Sault-Ste-Marie, ON, Canada, 1990. [Google Scholar]
- Harvey, B.; Brais, S. Effects of mechanized careful logging on natural regeneration and vegetation competition in the southeastern Canadian boreal forest. Can. J. For. Res. 2002, 32, 653–666. [Google Scholar] [CrossRef]
- Parent, S.; Ruel, J.-C. Chronologie de la croissance chez des semis de sapin baumier (Abies balsamea (L.) Mill.) après une coupe à blanc avec protection de la régénération. For. Chron. 2002, 78, 876–885. [Google Scholar] [CrossRef]
- Greene, D.F.; Zasada, J.C.; Sirois, L.; Kneeshaw, D.; Morin, H.; Charron, I.; Simard, M.-J. A review of the regeneration dynamics of North American boreal forest tree species. Can. J. For. Res. 1999, 29, 824–839. [Google Scholar] [CrossRef]
- Pothier, D.; Prévost, M. Photosynthetic light response and growth analysis of competitive regeneration after partial cutting in a boreal mixed stand. Trees-Struct. Funct. 2002, 16, 365–373. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. Silvics of North America: Volume 2. Hardwoods; Agriculture Handbook 654; United States Department of Agriculture (USDA), Forest Service: Washigton, DC, USA, 1990. [Google Scholar]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Conifer seedling recruitment in a southeastern Canadian boreal forest: The importance of substrate. J. Veg. Sci. 1998, 9, 575–582. [Google Scholar] [CrossRef]
- Robert, E.; Brais, S.; Harvey, B.D.; Greene, D. Seedling establishment and survival on decaying logs in boreal mixedwood stands following a mast year. Can. J. For. Res. 2012, 42, 1446–1455. [Google Scholar] [CrossRef]
- Bose, A.K.; Harvey, B.D.; Brais, S.; Beaudet, M.; Leduc, A. Constraints to partial cutting in the boreal forest of Canada in the context of natural disturbance-based management: A review. Forestry 2014, 87, 11–28. [Google Scholar] [CrossRef]
- Asselin, H.; Fortin, M.-J.; Bergeron, Y. Spatial distribution of late-successional coniferous species regeneration following disturbance in southwestern Québec boreal forest. For. Ecol. Manag. 2001, 140, 29–37. [Google Scholar] [CrossRef]
- MacDonald, G.B.; Cherry, M.L.; Thompson, D.J. Effect of harvest intensity on development of natural regeneration and shrubs in an Ontario boreal mixedwood stand. For. Ecol. Manag. 2004, 189, 207–222. [Google Scholar] [CrossRef]
- Prévost, M.; Dumais, D.; Pothier, D. Growth and mortality following partial cutting in a trembling aspen–conifer stand: Results after 10 years. Can. J. For. Res. 2010, 40, 894–903. [Google Scholar] [CrossRef]
- Doucet, R. La Régénération préétablie dans les peuplements forestiers naturels au Québec. For. Chron. 1988, 64, 116–120. [Google Scholar] [CrossRef]
- Prévost, M.; DeBlois, J. Shelterwood cutting to release coniferous advance growth and limit aspen sucker development in a boreal mixedwood stand. For. Ecol. Manag. 2014, 323, 148–157. [Google Scholar] [CrossRef]
- Gradowski, T.; Lieffers, V.J.; Landhäusser, S.M.; Sidders, D.; Volney, J.; Spence, J.R. Regeneration of Populus nine years after variable retention harvest in boreal mixedwood forests. For. Ecol. Manag. 2010, 259, 383–389. [Google Scholar] [CrossRef]
- Saucier, J.P.; Robitaille, A.; Grondin, P.; Bergeron, J.F.; Gosselin, J. Les Régions Écologiques du Québec MÉRidional; Ministère des Ressources Naturelles et de la Faune, Direction des Inventaires Forestiers: Quebec, QC, Canada, 2011. [Google Scholar]
- Environment Canada. Available online: http://www.climate.weatheroffice.ec.gc.ca (accessed on 28 May 2019).
- Vincent, J.-S.; Hardy, L. Dynamique et paléogéographie de l’inlandsis laurentidien. gpq 1977, 31, 357–372. [Google Scholar] [CrossRef]
- Harvey, B.; Leduc, A. Plan Général D’aménagement-Forêt D’enseignement et de Recherche du lac Duparquet (1998–2023); Groupe de Recherche en Écologie Forestière (UQAM) and Unité de Recherche et de Développement forestier de l’Abitibi-Témiscamingue (UQAT): Quebec, QC, Canada, 1999; p. 153. [Google Scholar]
- Dansereau, P.-R.; Bergeron, Y. Fire history in the southern boreal forest of northwestern Quebec. Can. J. For. Res. 1993, 23, 25–32. [Google Scholar] [CrossRef]
- Morin, H.; Laprise, D.; Bergeron, Y. Chronology of spruce budworm outbreaks near Lake Duparquet, Abitibi region, Quebec. Can. J. For. Res. 1993, 23, 1497–1506. [Google Scholar] [CrossRef]
- Campbell, E.M.; David, A. MacLean; Yves Bergeron. The Severity of Budworm-Caused Growth Reductions in Balsam Fir/Spruce Stands Varies with the Hardwood Content of Surrounding Forest Landscapes. For. Sci. 2008, 54, 195–205. [Google Scholar] [CrossRef]
- Soil Classification Working Group. The Canadian System of Soil Classification, 3rd ed.; Publ. 1646: Ottawa, ON, Canada, 1998; ISBN o-660-17404-9. [Google Scholar]
- SAS. JMP version 10; SAS Institute Inc.: Cary, NC, Canada, 2019. [Google Scholar]
- Vincent, A.B. Growth habits of mountain maple in the Ontario clay belt. For. Chron. 1965, 41, 330–344. [Google Scholar] [CrossRef]
- Messier, C.; Parent, S.; Bergeron, Y. Effects of overstory and understory vegetation on the understory light environment in mixed boreal forests. J. Veg. Sci. 1998, 9, 511–520. [Google Scholar] [CrossRef]
- Pothier, D.; Doucet, R.; Boily, J. The effect of advance regeneration height on future yield of black spruce stands. Can. J. For. Res. 1995, 25, 536–544. [Google Scholar] [CrossRef]
- Man, R.; Kayahara, G.J.; Rice, J.A.; MacDonald, G.B. Eleven-year responses of a boreal mixedwood stand to partial harvesting: Light, vegetation, and regeneration dynamics. For. Ecol. Manag. 2008, 255, 697–706. [Google Scholar] [CrossRef]
- Bose, A.K.; Brais, S.; Harvey, B.D. Trembling aspen (Populus tremuloides Michx.) volume growth in the boreal mixedwood: Effect of partial harvesting, tree social status, and neighborhood competition. For. Ecol. Manag. 2014, 327, 209–220. [Google Scholar] [CrossRef]
- Mitchell, S.J. The windthrow triangle: A relative windthrow hazard assessment procedure for forest managers. For. Chron. 1995, 71, 446–450. [Google Scholar] [CrossRef]
- Bouchard, M.; Kneeshaw, D.; Bergeron, Y. Mortality and stand renewal patterns following the last spruce budworm outbreak in mixed forests of western Quebec. For. Ecol. Manag. 2005, 204, 297–313. [Google Scholar] [CrossRef]
- Moulinier, J.; Lorenzetti, F.; Bergeron, Y. Effects of a forest tent caterpillar outbreak on the dynamics of mixedwood boreal forests of eastern Canada. Écoscience 2013, 20, 182–193. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Messier, C.; Burton, P.J.; Ruel, J.C.; Grover, B.E. Naturebased Silviculture for Sustaining a Variety of Boreal Forest Values. Chapter 13; Burton, P.J., Messier, C., Smith, D.W., Adamowicz, W.L., Eds.; Towards Sustainable Management of the Boreal Forest, NRC Research Press: Ottawa, ON, Canada, 2003; pp. 480–530. [Google Scholar]
- Rowe, J.S. Critique of Some Vegetational Concepts as Applied to Forests of Northwestern Alberta. Can. J. Bot. 1961, 39, 1007–1017. [Google Scholar] [CrossRef]
- Caners, R.T.; Kenkel, N.C. Forest stand structure and dynamics at Riding Mountain National Park, Manitoba, Canada. Community Ecol. 2003, 4, 185–204. [Google Scholar] [CrossRef]
- Cumming, S.G.; Schmiegelow, F.K.A.; Burton, P.J. Gap dynamics in boreal aspen stands: Is the forest older than we think? Ecol. Appl. 2000, 10, 744–759. [Google Scholar] [CrossRef]
- Perala, D.A. Prescribed Burning in an Aspen-Mixed Hardwood Forest. Can. J. For. Res. 1974, 4, 222–228. [Google Scholar] [CrossRef]
- Ilisson, T.; Chen, H.Y.H. The direct regeneration hypothesis in northern forests. J. Veg. Sci. 2009, 20, 735–744. [Google Scholar] [CrossRef]
- Kurkowski, T.A.; Mann, D.H.; Rupp, T.S.; Verbyla, D.L. Relative importance of different secondary successional pathways in an Alaskan boreal forest. Can. J. For. Res. 2008, 38, 1911–1923. [Google Scholar] [CrossRef]
- Bose, A.; Nelson, A.S.; Olson, M. Growth and mortality response of forest regeneration to partial harvesting varies by species shade tolerance. Can. J. For. Res. 2020, 0, 0. [Google Scholar] [CrossRef]
- Bourgeois, L.; Messier, C.; Brais, S. Mountain maple and balsam fir early response to partial and clear-cut harvesting under aspen stands of northern Quebec. Can. J. For. Res. 2004, 34, 2049–2059. [Google Scholar] [CrossRef]
- Beaudet, M.; Harvey, B.D.; Messier, C.; Coates, K.D.; Poulin, J.; Kneeshaw, D.D.; Brais, S.; Bergeron, Y. Managing understory light conditions in boreal mixedwoods through variation in the intensity and spatial pattern of harvest: A modelling approach. For. Ecol. Manag. 2011, 261, 84–94. [Google Scholar] [CrossRef]
- Frey, B.R.; Lieffers, V.J.; Landhäusser, S.M.; Comeau, P.G.; Greenway, K.J. An analysis of sucker regeneration of trembling aspen. Can. J. For. Res. 2003, 33, 1169–1179. [Google Scholar] [CrossRef]
- Frelich, L.E.; Reich, P.B. Spatial Patterns and Succession in a Minnesota Southern-Boreal Forest. Ecol. Monogr. 1995, 65, 325–346. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Mulak, T.L.; Lieffers, V.J. The effect of roots and litter of Calamagrostis canadensis on root sucker regeneration of Populus tremuloides. Forestry 2007, 80, 481–488. [Google Scholar] [CrossRef]
- Schneider, R.; Zhang, S.Y.; Swift, D.E.; Bégin, J.; Lussier, J.-M. Predicting selected wood properties of jack pine following commercial thinning. Can. J. For. Res. 2008, 38, 2030–2043. [Google Scholar] [CrossRef]
- Ashton, M.S.; Kelty, M.J. The Practice of Silviculture. In Applied Forest Ecology, 10th ed.; Wiley: Hoboken, NJ, USA, 2017; ISBN 9781119270959. [Google Scholar]
- Bretfeld, M.; Doerner, J.P.; Franklin, S.B. Radial growth response and vegetative sprouting of aspen following release from competition due to insect-induced conifer mortality. For. Ecol. Manag. 2015, 347, 96–106. [Google Scholar] [CrossRef]
- Solarik, K.A.; Volney, W.J.A.; Lieffers, V.J.; Spence, J.R.; Hamann, A. Factors affecting white spruce and aspen survival after partial harvest. J. Appl. Ecol. 2012, 49, 145–154. [Google Scholar] [CrossRef]
- Kneeshaw, D.; Bergeron, Y.; de Grandpré, L. Early response of Abies balsamea seedlings to artificially created openings. J. Veg. Sci. 1998, 9, 543–550. [Google Scholar] [CrossRef]
- Kneeshaw, D.D.; Bergeron, Y. Ecological factors affecting the abondance of advance regeneration in Quebec’s southwestern boreal forest. Can. J. For. Res. 1996, 26, 888–898. [Google Scholar] [CrossRef]
- Thomas, P.A.; Wein, R.W. Water availability and the comparative emergence of four conifer species. Can. J. Bot. 1985, 63, 1740–1746. [Google Scholar] [CrossRef]
- Ostry, M.E.; United States Forest Service. A Guide to Insect, Disease, and Animal Pests of Poplars; US Department of Agriculture, Forest Service: USA, 1989. Available online: https://naldc.nal.usda.gov/download/CAT89930507/PDF (accessed on 5 July 2020).
- Thorpe, H.C.; Thomas, S.C.; Caspersen, J.P. Tree mortality following partial harvests is determined by skidding proximity. Ecol. Appl. 2008, 18, 1652–1663. [Google Scholar] [CrossRef]
- Bose, A.K.; Harvey, B.D.; Brais, S. Sapling recruitment and mortality dynamics following partial harvesting in aspen-dominated mixedwoods in eastern Canada. For. Ecol. Manag. 2014, 329, 37–48. [Google Scholar] [CrossRef]
- Stone, D.M.; Elioff, J.D. Soil disturbance and aspen regeneration on clay soils: Three case histories. For. Chron. 2000, 76, 747–752. [Google Scholar] [CrossRef][Green Version]
- Grigal, D.F. Effects of extensive forest management on soil productivity. For. Ecol. Manag. 2000, 138, 167–185. [Google Scholar] [CrossRef]
- Brais, S. Persistence of Soil Compaction and Effects on Seedling Growth in Northwestern Quebec. Soil Sci. Soc. Am. J. 2001, 65, 1263–1271. [Google Scholar] [CrossRef]
- Puettmann, K.J.; D’Amato, A.W.; Arikian, M.; Zasada, J.C. Spatial impacts of soil disturbance and residual overstory on density and growth of regenerating aspen. For. Ecol. Manag. 2008, 256, 2110–2120. [Google Scholar] [CrossRef]
- Stone, D.M.; Kabzems, R. Aspen development on similar soils in Minnesota and British Columbia after compaction and forest floor removal. For. Chron. 2002, 78, 886–891. [Google Scholar] [CrossRef]
- Mundell, T.L.; Landhäusser, S.M.; Lieffers, V.J. Root carbohydrates and aspen regeneration in relation to season of harvest and machine traffic. For. Ecol. Manag. 2008, 255, 68–74. [Google Scholar] [CrossRef]
- Powers, R.F. On the Sustainable Productivity of Planted Forests. New Forests 1999, 17, 263–306. [Google Scholar] [CrossRef]
- Man, R.; Rice, J.A.; MacDonald, G.B. Five-year light, vegetation, and regeneration dynamics of boreal mixedwoods following silvicultural treatments to establish productive aspen–spruce mixtures in northeastern Ontario. Can. J. For. Res. 2010, 40, 1529–1541. [Google Scholar] [CrossRef]
- Startsev, A.D.; McNabb, D.H. Skidder traffic effects on water retention, pore-size distribution, and van Genuchten parameters of boreal forest soils. Soil Sci. Soc. Am. J. 2001, 65, 224–231. [Google Scholar] [CrossRef]
- Komatsu, H.; Katayama, A.; Hirose, S.; Kume, A.; Higashi, N.; Ogawa, S.; Otsuki, K. Reduction in soil water availability and tree transpiration in a forest with pedestrian trampling. Agric. For. Meteorol. 2007, 146, 107–114. [Google Scholar] [CrossRef]
- Steneker, G.A. Factors affecting the suckering of trembling aspen. For. Chron. 1974, 50, 32–34. [Google Scholar] [CrossRef]
- Fraver, S.; Wagner, R.G.; Day, M. Dynamics of coarse woody debris following gap harvesting in the Acadian forest of central Maine, U.S.A. Can. J. For. Res. 2002, 32, 2094–2105. [Google Scholar] [CrossRef]
- Alban, D.H.; Host, G.E.; Elioff, J.D.; Shadis, D.A. Soil and Vegetation Response to Soil Compaction and Forest Floor Removal after Aspen Harvesting; Research Paper NC-315; U.S. Dept. of Agriculture, Forest Service, North Central Forest Experiment Station: St. Paul, MN, USA, 1994. [Google Scholar]
- Greene, D.F.; Kneeshaw, D.D.; Messier, C.; Lieffers, V.; Cormier, D.; Doucet, R.; Coates, K.D.; Groot, A.; Grover, G.; Calogeropoulos, C. Modelling silvicultural alternatives for conifer regeneration in boreal mixedwood stands (aspen/white spruce/balsam fir). For. Chron. 2002, 78, 281–295. [Google Scholar] [CrossRef]
- Huffman, R.D.; Fajvan, M.A.; Wood, P.B. Effects of residual overstory on aspen development in Minnesota. Can. J. For. Res. 1999, 29, 284–289. [Google Scholar] [CrossRef]
- Landhäusser, S.M.; Lieffers, V.J. Growth of Populus tremuloides in association with Calamagrostis canadensis. Can. J. For. Res. 1998, 28, 396–401. [Google Scholar] [CrossRef]
- Raymond, P.; Ruel, J.-C.; Pineau, M. Effet d’une coupe d’ensemencement et du milieu de germination sur la régénération des sapinières boréales riches de seconde venue du Québec. For. Chron. 2000, 76, 643–652. [Google Scholar] [CrossRef]
- Taylor, A.R.; Chen, H.Y.H. Multiple successional pathways of boreal forest stands in central Canada. Ecography 2011, 34, 208–219. [Google Scholar] [CrossRef]
- Narukawa, Y.; Iida, S.; Tanouchi, H.; Abe, S.; Yamamoto, S.-I. State of fallen logs and the occurrence of conifer seedlings and saplings in boreal and subalpine old-growth forests in Japan. Ecol. Res. 2003, 18, 267–277. [Google Scholar] [CrossRef]
- Harmon, M.E.; Franklin, J.F. Tree Seedlings on Logs in Picea-Tsuga Forests of Oregon and Washington. Ecology 1989, 70, 48–59. [Google Scholar] [CrossRef]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Substrate and litterfall effects on conifer seedling survivorship in southern boreal stands of Canada. Can. J. For. Res. 2003, 33, 672–681. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Plant litter. Decomposition, Humus Formation, Carbon Sequestration, 3rd ed.; Springer: Heidelberg, Germany, 2014; ISBN 9783642388200. [Google Scholar]
- Lavertu, D.; Mauffette, Y.; Bergeron, Y. Suckering success of aspen (Populus tremuloides Michx.) in relation to stand age and soil disturbance. J. Veg. Sci. 1994, 5, 561–568. [Google Scholar] [CrossRef]
- Côté, S.; Bélanger, L. Variations de la régénération préétablie dans les sapinières boréales en fonction de leurs caractéristiques écologiques. Can. J. For. Res. 1991, 21, 1779–1795. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A. The Role of Physical Damage in the Seedling Mortality Regime of a Neotropical Rain Forest. Oikos 1989, 55, 225. [Google Scholar] [CrossRef]
- Metcalfe, D.J.; Grubb, P.J. The responses to shade of seedlings of very small-seeded tree and shrub species from tropical rain forest in Singapore. Funct. Ecol. 1997, 11, 215–221. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatment | Harvest Year | Sampling Interval (y) | Stand Basal Area (m2 ha−1) | |
|---|---|---|---|---|
| Pre-Harvest | Post-Harvest | |||
| Cntrl | 40.8 | 38.0 | ||
| PC | 1998–2010 | 7–19 | 45.9 | 23.2 |
| CPRS | 1998–2004 | 13–19 | 46.8 | 4.0 |
| Species | Treatment | |||
|---|---|---|---|---|
| Control | PC | CPRS | p-Value | |
| Prior to treatment | ||||
| Trembling aspen | 31.3 ± 2.1 | 30.4 ± 2.0 | 31.7 ± 1.7 | 0.911 |
| White birch | 1.2 ± 2.2 | 1.8 ± 2.0 | 1.1 ± 2.0 | 0.438 |
| Balsam fir | 2.6 ± 2.3 | 2.9 ± 2.2 | 3.6 ± 4.2 | 0.598 |
| White spruce | 1.6 ± 1.2 | 2.7 ± 3.3 | 2.4 ± 2.8 | 0.685 |
| Black spruce | 1.2 ± 1.2 | 2.5 ± 3.4 | 1.6 ± 1.2 | 0.085 |
| Jack pine | 2.7 ± 1.4 | 5.2 ± 4.2 | 6.1± 4.2 | 0.735 |
| White cedar | 0.2 ± 1.4 | 0.3 ± 1.5 | 0.4 ± 1.2 | 0.454 |
| Total | 40.8 ± 2.3 | 45.9 ± 3.7 | 46.8 ± 2.0 | 0.264 |
| After treatment | ||||
| Trembling aspen | 24.4a ± 1.6 | 13.8b ± 2.2 | 3.1c ± 2.4 | <0.001 |
| White birch | 1.5a ± 2.0 | 1.3a ± 1.6 | 0.2b ± 0.6 | 0.008 |
| Balsam fir | 6.2a ± 4.1 | 2.4a ± 1.7 | 0.6b ± 1.5 | <0.001 |
| White spruce | 2.3a ± 2.6 | 3.0a ± 2.6 | 0.1b ± 0.3 | 0.030 |
| Black spruce | 1.6 ± 3.3 | 0.2 ± 0.7 | 0.0 ± 0.0 | 0.076 |
| Jack pine | 1.6 ± 4.2 | 2.4 ± 4.0 | 0.0 ± 0.0 | 0.085 |
| White cedar | 0.4 ± 1.4 | 0.1 ± 0.5 | 0.0 ± 0.0 | 0.551 |
| Total | 38.0a ± 2.1 | 23.2b ± 1.4 | 4.0c ± 0.5 | <0.001 |
| Species | Treatment | |||
|---|---|---|---|---|
| Control | PC | CPRS | p-Value | |
| Seedlings | ||||
| Trembling aspen | 1417b ± 321 | 5188a ± 884 | 1667b ± 561 | 0.024 |
| White birch | 42 ± 42 | 104 ± 61 | 21 ± 21 | 0.436 |
| Balsam fir | 12,042a ± 2135 | 13,604a ± 1719 | 3583b ± 964 | <0.001 |
| White spruce | 500b ± 132 | 1979a ± 596 | 417b ± 165 | 0.048 |
| Black spruce | 479 ± 209 | 146 ± 71 | 312 ± 163 | 0.678 |
| Jack pine | 0 ± 0 | 0 ± 0 | 83 ± 83 | |
| White cedar | 0 ± 0 | 63 ± 35 | 0 ± 0 | |
| Total | 14,473a ± 13,268 | 21,084b ± 19,282 | 6083c ± 5160 | <0.001 |
| Saplings | ||||
| Trembling aspen | 173c ± 56 | 1486b ± 320 | 10,193a ± 111 | <0.001 |
| White birch | 60 ± 20 | 233 ± 72 | 533 ± 112 | 0.225 |
| Balsam fir | 1093 ± 175 | 846 ± 121 | 1666 ± 711 | 0.316 |
| White spruce | 60 ± 21 | 103 ± 24 | 76 ± 32 | 0.745 |
| Black spruce | 53 ± 28 | 10 ± 7 | 80 ± 47 | 0.468 |
| Jack pine | 0 ± 0 | 0 ± 0 | 53 ± 53 | |
| White cedar | 0 ± 0 | 7 ± 7 | 0 ± 0 | |
| Total | 1439b ± 832 | 2685b ± 1154 | 12,601a ± 10,269 | <0.001 |
| Species | Treatment | |||||
|---|---|---|---|---|---|---|
| Control | PC-T | PC-OT | CPRS-T | CPRS-OT | p-Value | |
| Trembling aspen | 2931b ± 635 | 6667a ± 1280 | 2073b ± 735 | 366c ± 316 | 64c ± 73 | <0.001 |
| White birch | 0 ± 0 | 0 ± 0 | 0 ± 0 | 61 ± 71 | 50 ± 65 | 0.697 |
| Balsam fir | 9083a ± 1028 | 1710b ± 359 | 3170b ± 316 | 119c ± 84 | 313c ± 190 | <0.001 |
| White spruce | 500 ± 446 | 329 ± 195 | 366 ± 202 | 113 ± 110 | 188 ± 148 | 0.429 |
| Black spruce | 125 ± 107 | 0 ± 0 | 0 ± 0 | 120 ± 120 | 188 ± 148 | 0.530 |
| Jack pine | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 63 ± 62 | |
| White cedar | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | |
| Total | 12,639a ± 255 | 8706b ± 224 | 5609b ± 230 | 666c ± 220 | 866c ± 219 | <0.001 |
| Well-Decomposed Woody Debris | Deciduous Litter | Coniferous Litter | Basal Area | ||||
|---|---|---|---|---|---|---|---|
| Trembling Aspen | Balsam Fir | ||||||
| Pre-Harvest | Post-Harvest | Pre-Harvest | Post-Harvest | ||||
| Aspen seedling abundance | 0.249 (0.018) | 0.150 (0.158) | −0.219 (0.038) | 0.364 (<0.001) | 0.039 (0.717) | 0.312 (0.003) | 0.136 (0.200) |
| Fir seedling abundance | 0.210 (0.047) | −0.413 (< 0.001) | 0.445 (<0.001) | 0.327 (0.002) | −0.212 (0.045) | 0.007 (0.949) | 0.150 (0.157) |
| Total hardwood abundance | 0.252 (0.016) | 0.145 (0.174) | −0.216 (0.041) | 0.363 (<0.001) | 0.039 (0.714) | 0.314 (<0.001) | 0.135 (0.714) |
| Total conifer abundance | 0.215 (0.042) | −0.215 (0.042) | 0.215 (<0.001) | 0.036 (0.001) | −0.207 (0.051) | −0.003 (0.978) | 0.160 (0.131) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maleki, K.; Nguema Allogo, F.; Lafleur, B. Natural Regeneration Following Partial and Clear-Cut Harvesting in Mature Aspen-Jack Pine Stands in Eastern Canada. Forests 2020, 11, 741. https://doi.org/10.3390/f11070741
Maleki K, Nguema Allogo F, Lafleur B. Natural Regeneration Following Partial and Clear-Cut Harvesting in Mature Aspen-Jack Pine Stands in Eastern Canada. Forests. 2020; 11(7):741. https://doi.org/10.3390/f11070741
Chicago/Turabian StyleMaleki, Kobra, Freddy Nguema Allogo, and Benoit Lafleur. 2020. "Natural Regeneration Following Partial and Clear-Cut Harvesting in Mature Aspen-Jack Pine Stands in Eastern Canada" Forests 11, no. 7: 741. https://doi.org/10.3390/f11070741
APA StyleMaleki, K., Nguema Allogo, F., & Lafleur, B. (2020). Natural Regeneration Following Partial and Clear-Cut Harvesting in Mature Aspen-Jack Pine Stands in Eastern Canada. Forests, 11(7), 741. https://doi.org/10.3390/f11070741