1. Introduction
In the past, management of forests in Turkey was aimed at maximizing timber production regardless of sustainability of the multiple forest functions. However, forest stands did not receive any silvicultural treatment, all standing trees are above their economic rotation age, and almost half of the Turkish forests are considered to be degraded in terms of wood production [
1]. The majority of the species-rich and productive forests are dominated by eastern beech (
Fagus orientalis Lipsky) and mainly located in the mesic parts of the Black Sea Region [
2]. Eastern beech accounts for 24% of the forested area and 28% of the total stand volume in the region [
3,
4]. Topographic heterogeneity and a humid, temperate, oceanic climate, with some forests having highly diversified structure and texture, are the main characteristics of this region [
3,
5,
6].
Historical documents and the Ottoman archives indicate that most of the beech forests in this region are remnants of once species-rich forests [
7]. The long-lasting anthropogenic pressure of various civilizations on forests and management systems aiming at even-aged forests in the Black Sea Region has led to the pronounced dominancy of beech or to the creation of monospecific beech stands [
4,
7,
8]. Extensive fires in the 18th and 19th century also largely supported the creation of beech forests [
9,
10,
11]. Close-to-nature management system with maximum preference of natural regeneration is now considered to have gradually replaced the traditional shelterwood system in today’s beech forests, similarly to Europe [
4,
12,
13,
14,
15]. In near-natural beech forests, natural regeneration appears abundantly in small- or medium-sized canopy gaps created as a result of dieback or disturbance of single trees or their groups in mature stands [
16,
17,
18,
19].
The knowledge of gap size in the canopy closure of mature beech stands is important to understand the natural regeneration process [
20,
21]. For instance, Sefidi et al. [
17] stated that the majority of stand gaps (41%) were formed due to individual tree-falls in natural stands of eastern beech and nearly 60% of such gaps were smaller than 200 m
2 in size. Availability of light, temperature, moisture, and nutrients for tree species growing in recently formed gaps is determined by the size and position of gaps [
4,
22,
23] and microsite conditions [
19,
24]. Site conditions, parent stand structure, and parameters of the gap and its dynamics substantially influence species composition, quantity, and prosperity of natural regeneration [
18,
19,
23,
25,
26]. On the other site, the opening of gaps in the tree and shrub canopy provokes the loss of nemoral conditions and limits the development of several characteristic deciduous forest taxa that are most vulnerable to increased evapotranspiration [
27]. The knowledge of the structure and development of forest stands provides important information for their sustainable management in the times of global climate changes [
12,
13,
15,
28,
29,
30,
31,
32,
33]. Moreover, forest ecosystems are highly vulnerable to climate change (warming, prolonged droughts, climatic anomalies, and occurrence of fires) in temperate and Mediterranean zones and could suffer a decline in their habitat suitability if ongoing observed climate trends continue in the future [
34,
35]. Climate factors are the main driver of forest growth [
36,
37,
38,
39,
40,
41], such as in eastern beech [
42,
43,
44].
Large areas of semi-natural eastern beech forests have been converted to young poor stands mostly through natural regeneration [
4,
45]. Artificial regeneration can be also used for supplementary planting [
46] and for introduction of economically valuable tree species, e.g., wild cherry (
Prunus avium L.), sweet chestnut (
Castanea sativa Mill.), wild service tree (
Sorbus torminalis L.), and other valuable broadleaves to increase biodiversity [
47,
48,
49,
50]. Moreover, naturally growing species such as sweet chestnut, maple (
Acer spp.), wild cherry, ash (
Fraxinus spp.), mountain ash (
Sorbus spp.), elm (
Ulmus spp.), linden (
Tilia spp.), and goat-willow (
Salix spp.) are included in the natural mixture of the beech forests of the Black Sea Region [
8,
50].
There is a frequent occurrence of pontic rhododedron (
Rhododendron ponticum L.), an aggressive invasive shrubby species, in the understory of eastern beech forests. It occurs mainly on acidic soils in moist mountain areas up to the altitude of 2000 m a.s.l. in the Black Sea Region of Turkey [
7,
15]. This invasive shrub substantially eliminates the occurrence of natural regeneration of other species, subsequent growth of recruits, and stand diversity, causing site and production losses in the forest sector [
7,
15,
51]. This shrub grows up to 8 m in height, bearing a developed lignotuber from which many axes or branches arise [
52]. This shrub is aggressively invasive and can spread from seed and vegetative propagation. The future of the region’s forests is heavily threatened through invasion of this species [
4,
7,
15,
53]. Moreover, current distribution ranges of invasive pontic rhododendron can be modified in the future by ongoing global climate change [
54], which may lead to its expansion into new areas [
55].
Decisions on the methods of differentiated management of forests on ecological principles are complicated to ensure their ecological stability and biodiversity [
28,
56]. Models can be applied for support of a decision-making and planning process that helps finding the optimum solution to forest management [
57]. Forest management and silviculture treatments (including thinning, promoting optimal growing conditions, reducing hosts, soil preparation, and consistent monitoring) then play the main role to control invasive alien species [
58,
59] and protect, in our case, threatened beech forests from aggressive invasive pontic rhododendron [
60]. However, optimal management strategies should always be based on in-depth investigation and evaluation of species-specific site and stand conditions, especially stand structure and development of the natural and near-natural forests [
61]. Our study can contribute to the solution of this serious problem.
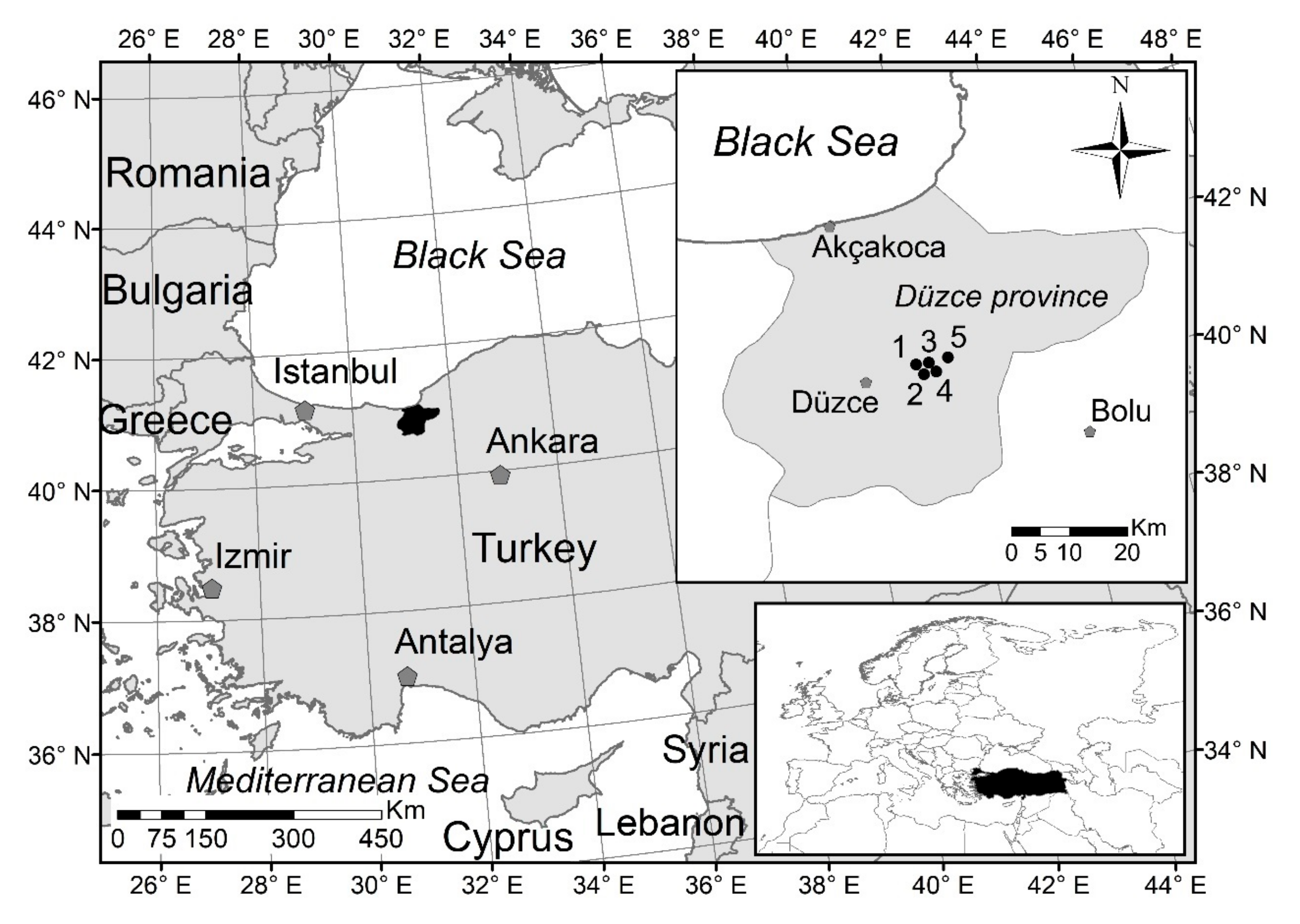
A general objective of our study was to evaluate effect of invasive pontic rhododendron on the structure of eastern beech stands in the north-west of Turkey in the Black Sea Region, with an emphasis on natural regeneration. Specific objectives were (1) to evaluate the parameters describing production, growth, diversity and structures of eastern beech stands, (2) to quantify and evaluate the effects of invasive pontic rhododendron and tree layer on the prosperity of natural regeneration in commercial and noncommercial forests, (3) to assess the effects of stand structure on the occurrence and expansion of pontic rhododendron, and (4) to quantify and evaluate the interactions among rhododendron, natural regeneration, stand production, and structural diversity.
4. Discussion
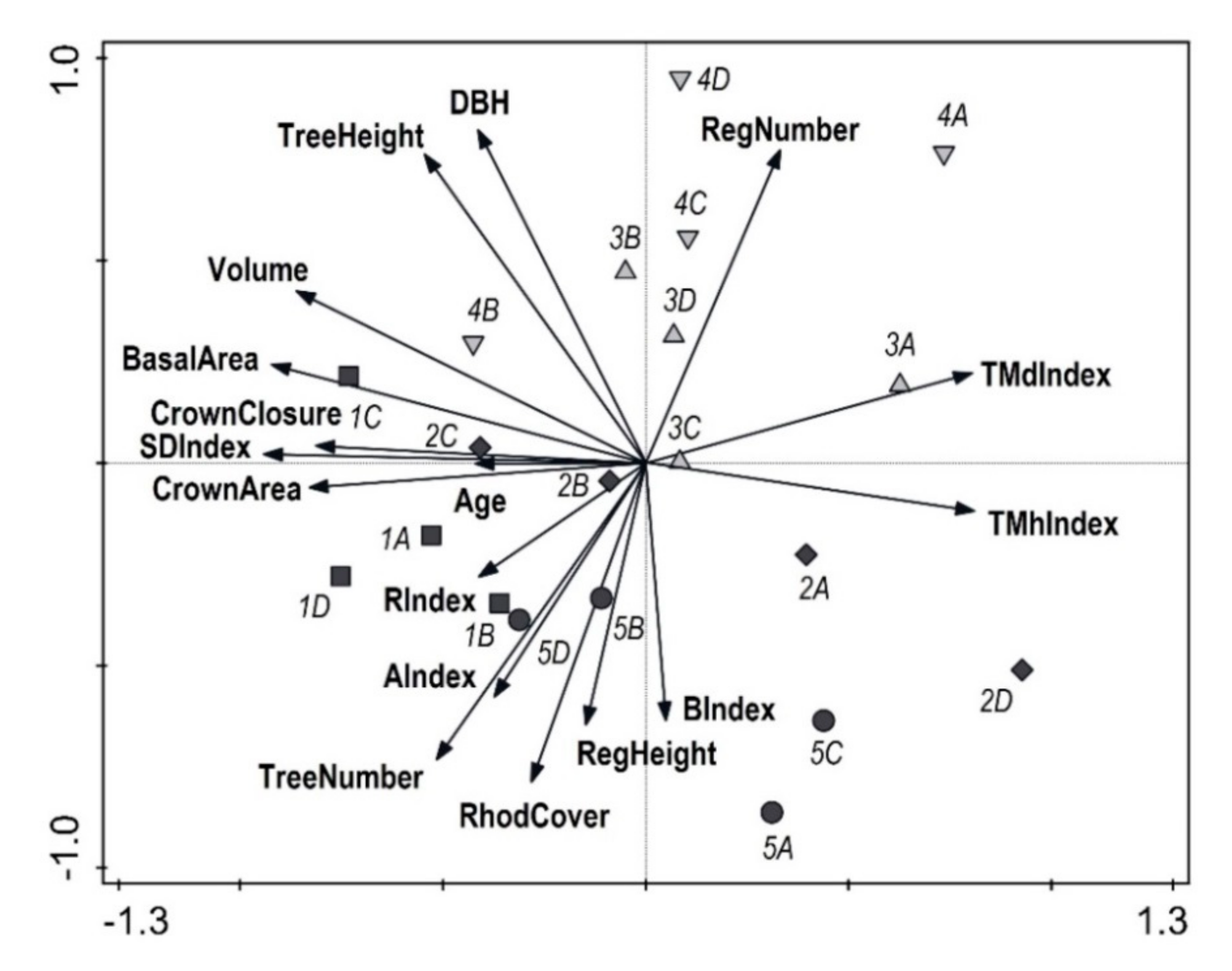
Eastern beech forest is one of the most important forest communities in the Black Sea Region [
82] and Turkey, where it occupies 1.7 million hectares [
14]. However, dynamics and structure of these valuable forest ecosystems are, in the long-term, significantly negatively affected by aggressive invasive pontic rhododendron [
83,
84]. A low species diversity was characteristic for the studied eastern beech stands in the BSR of Turkey in terms of species richness, heterogeneity, and evenness. Poor diversity is caused by strong competition of dense rhododendron cover and inappropriate management in its removal [
85,
86]. The horizontal structure of the tree layer was indistinctly aggregated or moderately regular while the random spatial pattern of trees across the research plots prevailed. Zenner et al. [
87] also documented the prevailing random distribution of trees in near-natural stands of eastern beech in the Caspian Iran. The initial spatial pattern of near-natural beech stands is usually aggregated or randomly irregular and it turns to a moderately regular pattern in favorable conditions [
88,
89]. This may be due to gradual growth, competition among trees, and expansion of crowns as beech crowns are able to promptly react to the changes in the normal environmental condition [
90].
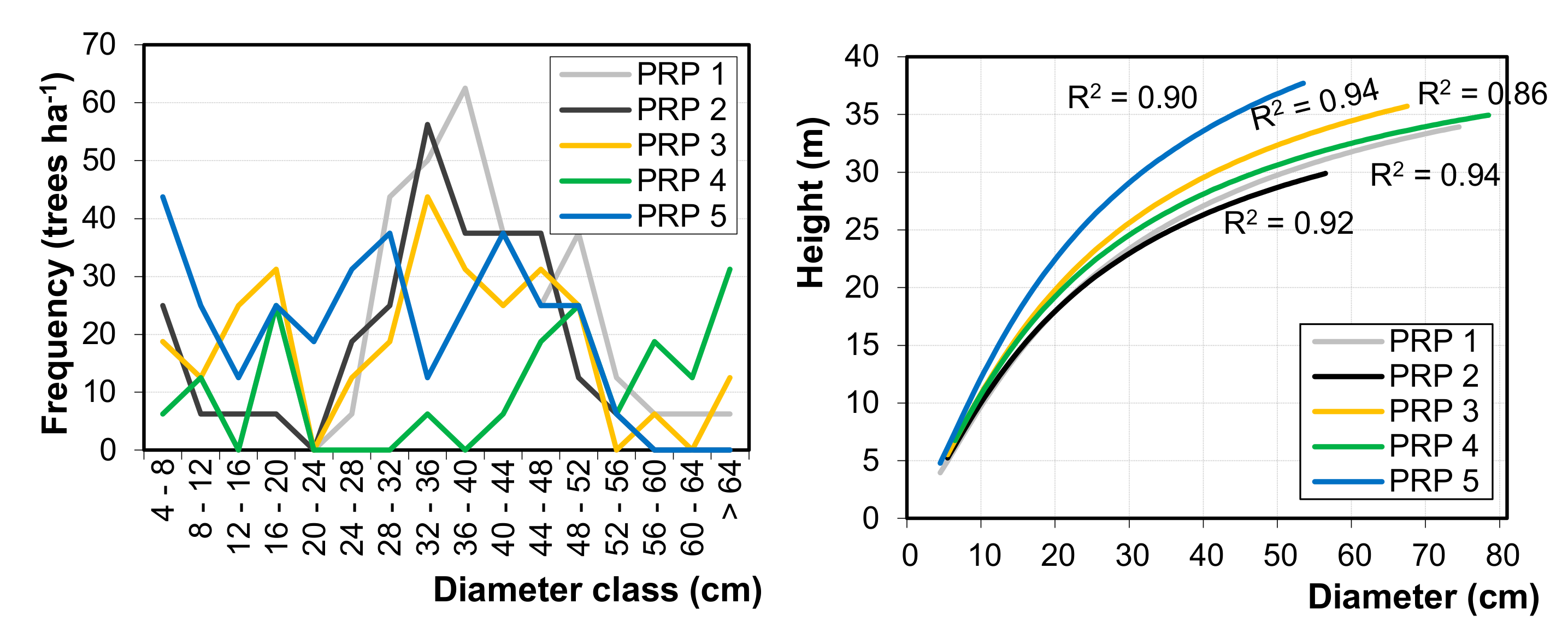
The vertical structure of investigated stands varied from medium to very high. Diameter structure was also medium-diversified. Frequency distribution of diameter of the tree layer was mostly bell-shaped, which documents the stage of optimum [
54]. Similarly, other studies show Gaussian distribution of diameters of eastern beech in the mature stands. Atalay [
3] and Eşen et al. [
4] also reported the relatively highest forest biodiversity in the oriental beech stands in the BSR within Turkish forests. A high biodiversity of near-natural forest stands with oriental beech was also reported in Iran [
87,
91]. These studies show that forests create the structural mosaic with alternation of particular developmental stages and phases. Sagheb-Talebi and Schütz [
92] found irregular and diversified structure with a high proportion of large trees in the natural beech forests of the Caspian Iran.
The number of trees on the studied PRPs varies from 175 to 381 trees ha
−1. These numbers are roughly consistent with the data from the Elborz Mountain range along the southern Caspian Sea, i.e., 219–443 trees ha
−1 [
87]. In comparable stands of eastern beech in Iran, Akhavan et al. [
93] reported 302–454 trees ha
−1 and Sefidi et al. [
17] reported 198 trees ha
−1 on average. The highest stand volume was found on PRP 1, reaching 589 m
3 ha
−1, and the lowest stand volume was on PRP 2, i.e., 331 m
3 ha
−1. In eastern beech stands of identical growth conditions, Hassani and Amani [
94] reported higher volumes (i.e., 562–678 m
3 ha
−1) and Sefidi et al. [
17] documented 386 m
3 ha
−1. Basal area of the studied stands was in the range of 28.8–46.1 m
2 ha
−1, and similarly Zenner et al. [
87] found stand basal area varying from 33.7 to 44.1 m
2 ha
−1.
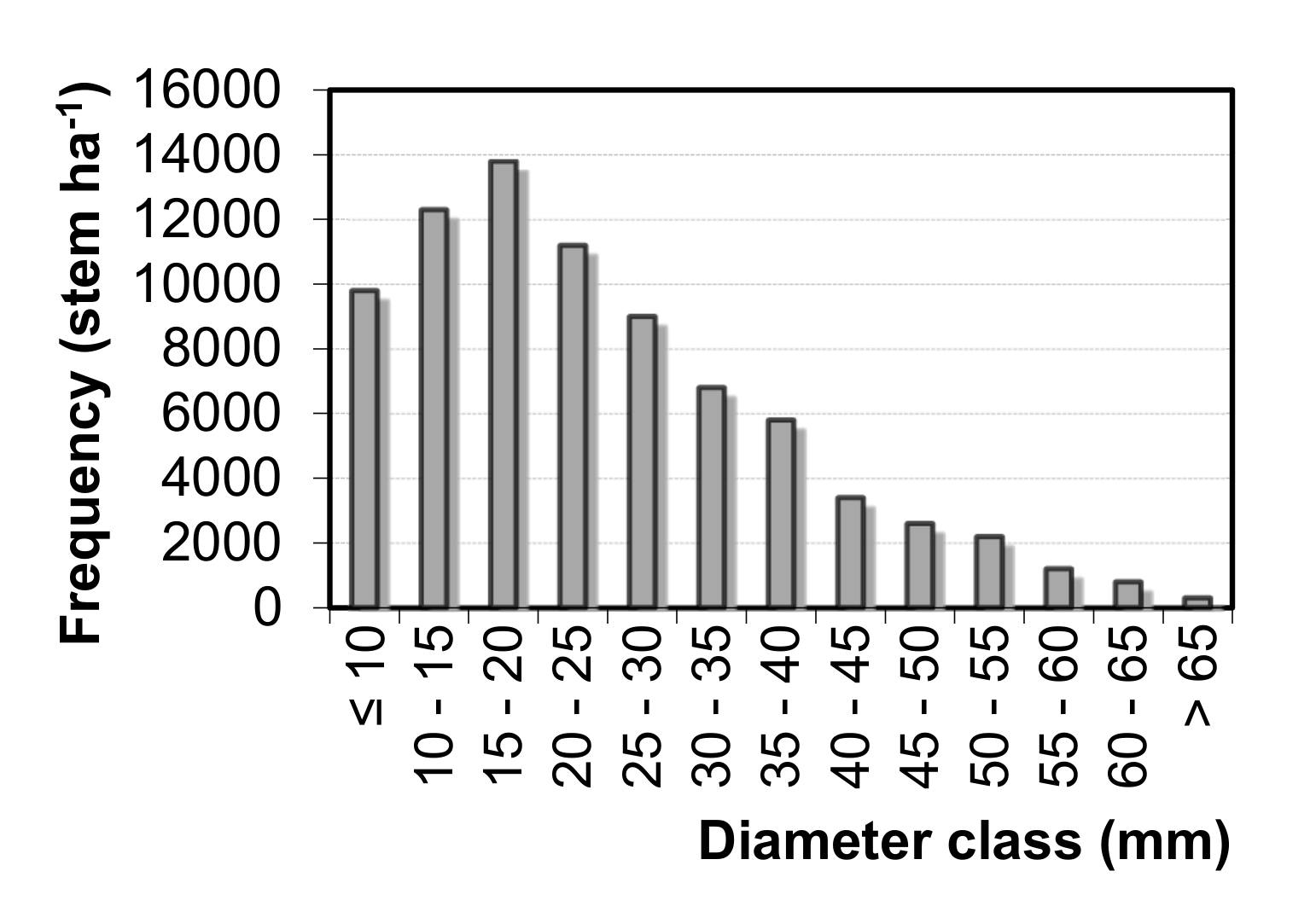
Natural regeneration of beech forests is strongly limited by dense undergrowth of aggressive invasive pontic rhododendron, which is very abundant in eastern beech stands in the entire western BSR [
7,
95,
96,
97]. The number of monopodial woody stems varied from 72,178 to 86,884 stems ha
−1 on our PRP, and this number was between the values reported by Eşen et al. [
7], which were 48,000 and 91,000 stems ha
−1 for
R. ponticum and
R. flavum, respectively. High rhododendron production was also observed from neighboring districts, where biomass storage capacity of pontic rhododendron reached up to 40,035 kg ha
−1 with preference of sunny exposure and altitude to 500 m a. s. l. [
98]. From this reason, pontic rhododendron can be (except positive effect of clearing on tree regeneration) used for fuel and in wood industry, especially for manufacture of density fiberboard panels [
99,
100]. Our study also reported dense rhododendron cover (81%–97%) and low tree regeneration (113–619 recruits ha
−1). Only two research plots (PRP 3 and 4), where invasive rhododendron was cleared 3 and 6 years ago, respectively, contained a high number of tree regeneration (63,987–105,075 recruits ha
−1). Comparable amount of beech natural regeneration (24,500 to 123,400 seedlings ha
−1) was observed in the Sökü district in Western part of BSR [
83]. Three to four individuals per m
2 may be considered as a sufficient number of tree regeneration for the prosperous development of stands [
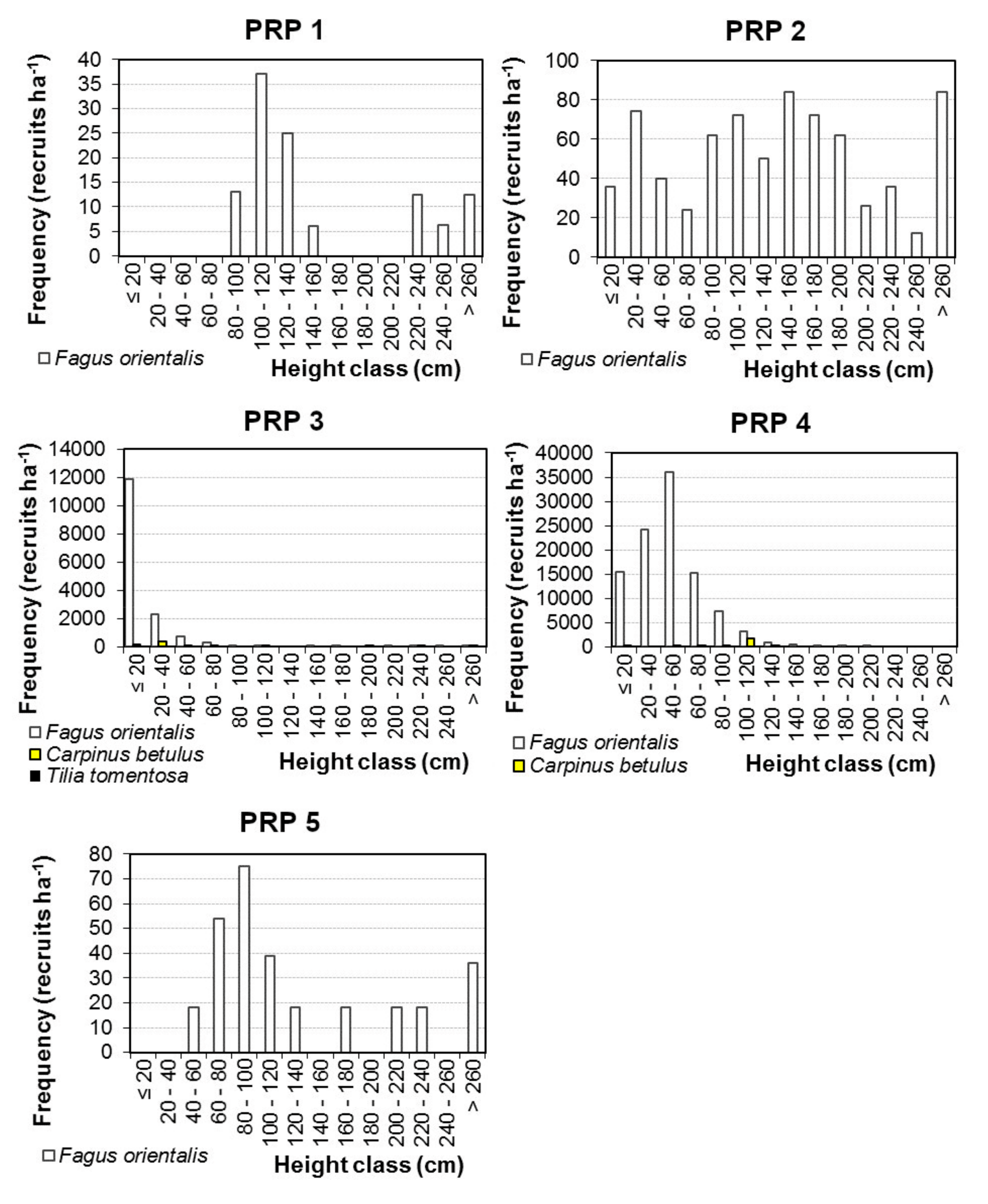
101]. From this perspective, only tree regeneration on PRP 3 and 4 with 0–3% cover percentage of rhododendron may be considered as sufficient. On the average, eastern beech accounted for 97% of natural regeneration (recruits) on the studied research plots. Because site preparation was done in seed mast year of beech, the seed status of other tree species was not taken into consideration. As a result, regeneration was purely dominated by beech seedlings, but other tree species consisted of admixture of mature stands. However, rhododendron had an opposite influence on the mean height of recruits. Due to competitiveness of this woody shrub, the height of recruits was significantly taller on the research plots with a high cover percentage of rhododendron (87.2–221.5 cm) compared to the recruit heights (23.2–43.3 cm) on PRPs where rhododendron was removed. While the influence of rhododendron cover (light competition) on regeneration was strong, influence of overstory canopy closure on recruit number and height was not proved.
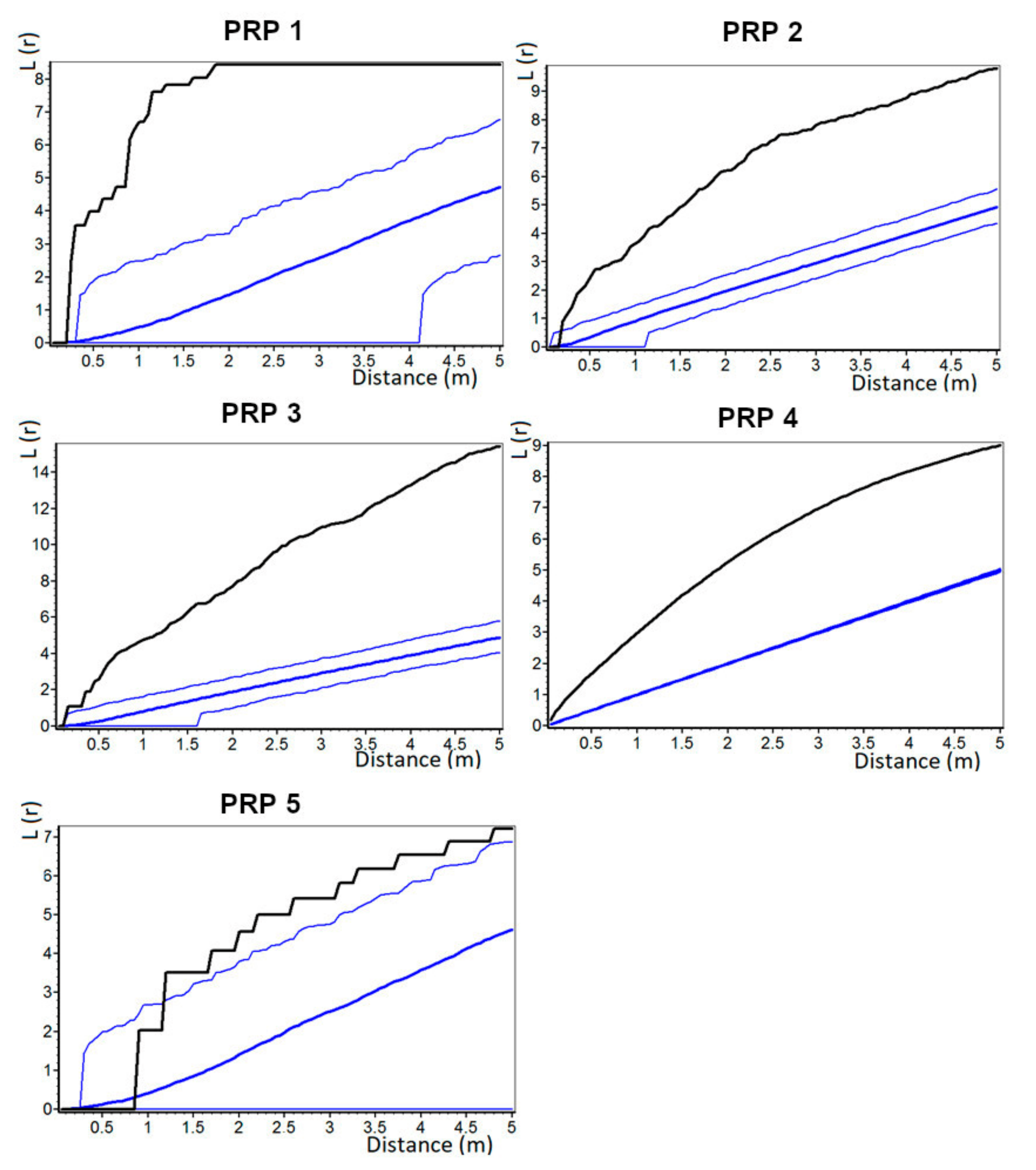
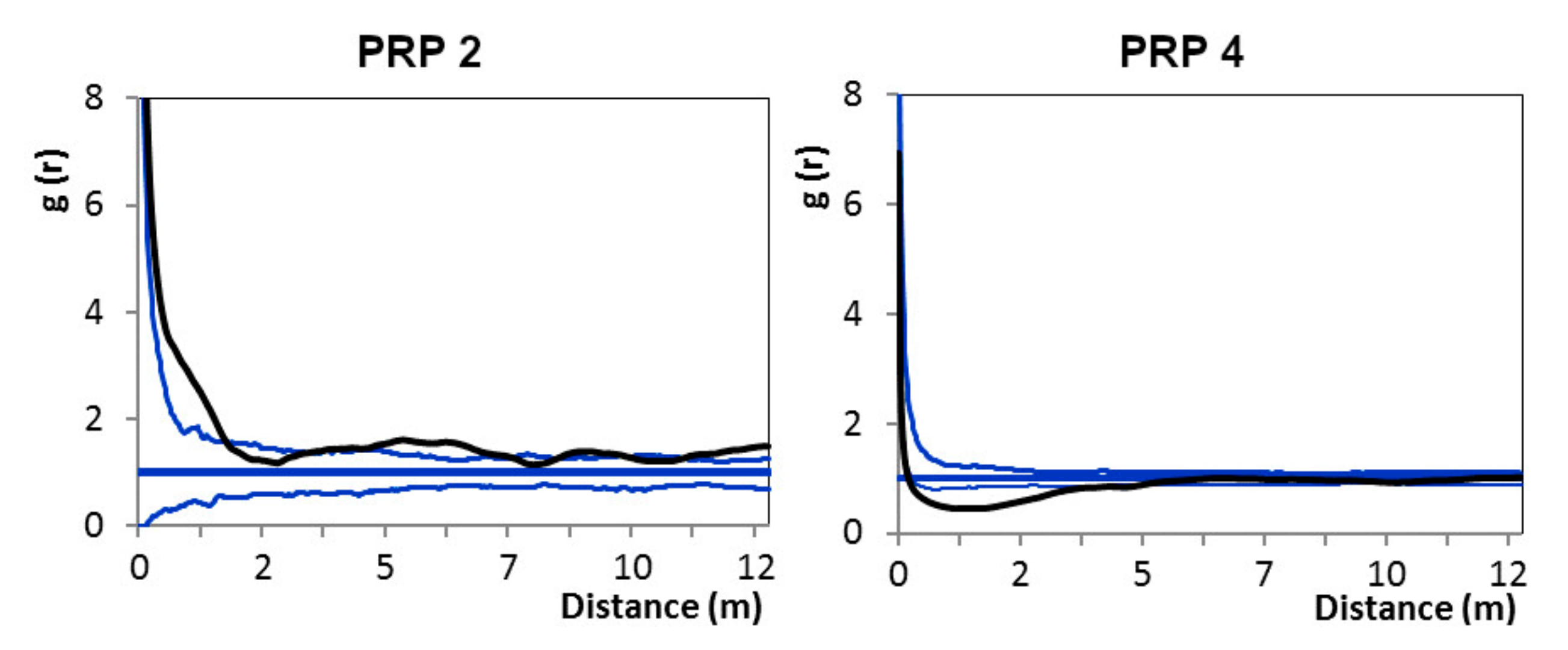
The horizontal structure of natural regeneration was distinctly aggregated on all studied PRPs. A similar pattern was also found in comparable oriental beech stands [
93]. The situation is identical in European beech (
Fagus sylvatica L.) [
19,
102]. A tendency towards aggregation of seedlings and in the lowerstory is a result of prosperous growth of natural regeneration in canopy gaps that originated after dead trees [
23,
103]. As for the spatial pattern (contrary to recruit number and height), parent trees had a significant negative influence on the individual recruits within a 4 m radius from tree stem on the PRPs, where rhododendron cover with generative regeneration prevailed. Vacek et al. [
19] reported that distribution of parent trees negatively affected the placement of natural regeneration of European beech within a 2.5 m radius. This negative spatial interaction between natural regeneration and tree layer in small spacing may be caused by insufficient light conditions for growth of seedlings closer to mature tree. However, trees had positive influence on natural regeneration to the distance of 1.5 m on the research plots with dense rhododendron cover. This is caused by rich root sprouting of eastern beech [
17]. Natural regeneration, especially from the seed without support from the parent plant, has no chance to survive under the dense rhododendron cover compared to the sprouts or suckers [
7]. The species diversity of natural regeneration was low on the study site; however, it was higher on PRP 3 and 4, where rhododendron was removed. On other plots, no other tree species (except beech) occurred. Removal of rhododendron may give an opportunity to the other tree species to establish regeneration. However, during the rhododendron clearance, the other tree species are usually removed from the beech forests, and frequent use of bulldozers had a negative effect on the seed bank during the soil removal [
86].
The mineral soil exposure during site preparation is often recommended for sufficient natural regeneration because the litter layer does not provide a favorable seedbed for beech seeds [
96,
104]. In this context, Agestam et al. [
105] stated that the suitable preparation of site conditions, and especially of the soil surface with the exposure of mineral contents, increased the germination rate of beech seeds compared to sites without soil preparation in southern Sweden. One year after the bulldozer removal of rhododendron undergrowth along with thickness of 5–10 cm soil layer in beech forests of the BSR, Yildiz et al. [
95] and Sarginci [
106] reported 300,000–700,000 beech seedlings ha
−1. Such a method of soil preparation is more suitable from short-time economic aspects than manual cutting or clearance of rhododendron. At the same time, it leads to the accelerated mineralization causing nutrient loss, decrease in microbial activity, and soil compaction [
95,
101,
107]. Soil compaction can also hinder the root growth, decrease tolerance to drought, and limit the uptake of immobile soil nutrients [
108]. For these reasons, and especially from the perspective of the sustainable forest management, manual cutting and clearance of rhododendron undergrowth seems better than its removal by a bulldozer with great disturbance of the soil surface. In addition, during site preparation, scraping the forest floor by a rake-equipped bulldozer significantly disturbed the seed banks. A similar opinion was published by [
101], who supported manual clearance of rhododendron and its burning because of their efficiency for the long-term productivity of oriental beech stands in Turkey.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}