Carbon Stock Estimations in a Mediterranean Riparian Forest: A Case Study Combining Field Data and UAV Imagery

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Riparian Typologies
2.2. Field Data Collection
2.2.1. Tree Inventory
2.2.2. Understory Biomass
2.2.3. Organic and Mineral Soil Layer
2.3. Allometric Models for Biomass Quantification
2.4. UAV Imagery Acquisition and Processing
2.5. Object-Based Classification and AGB Remote Estimation
2.5.1. Image Segmentation
2.5.2. Classification and Accuracy Assessment
3. Results
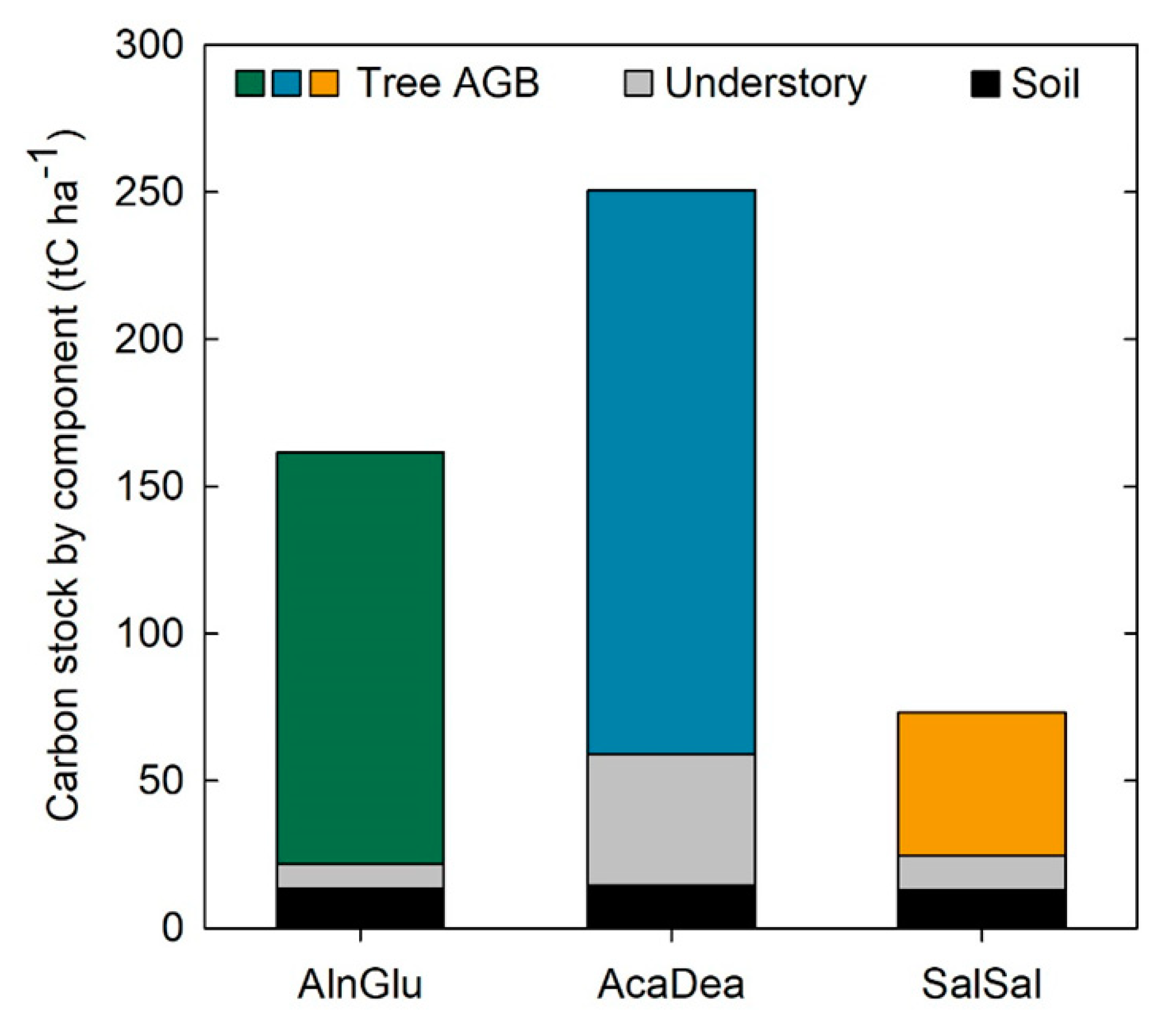
3.1. Carbon Stocks in Situ
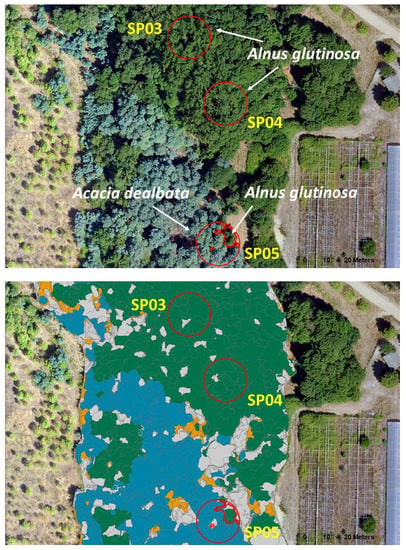
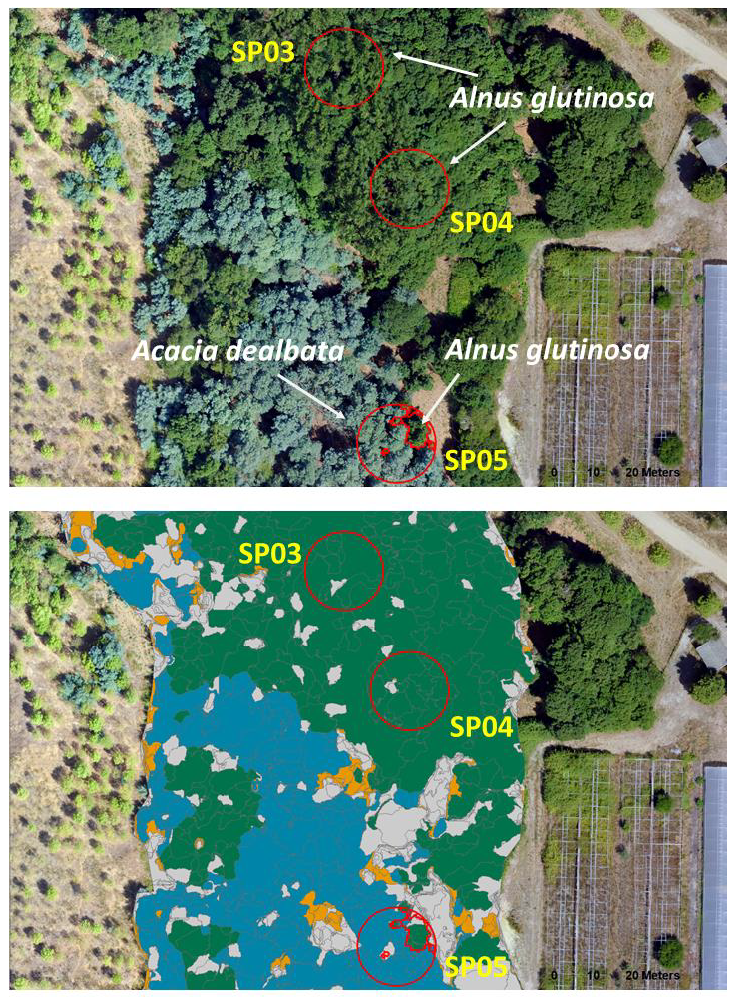
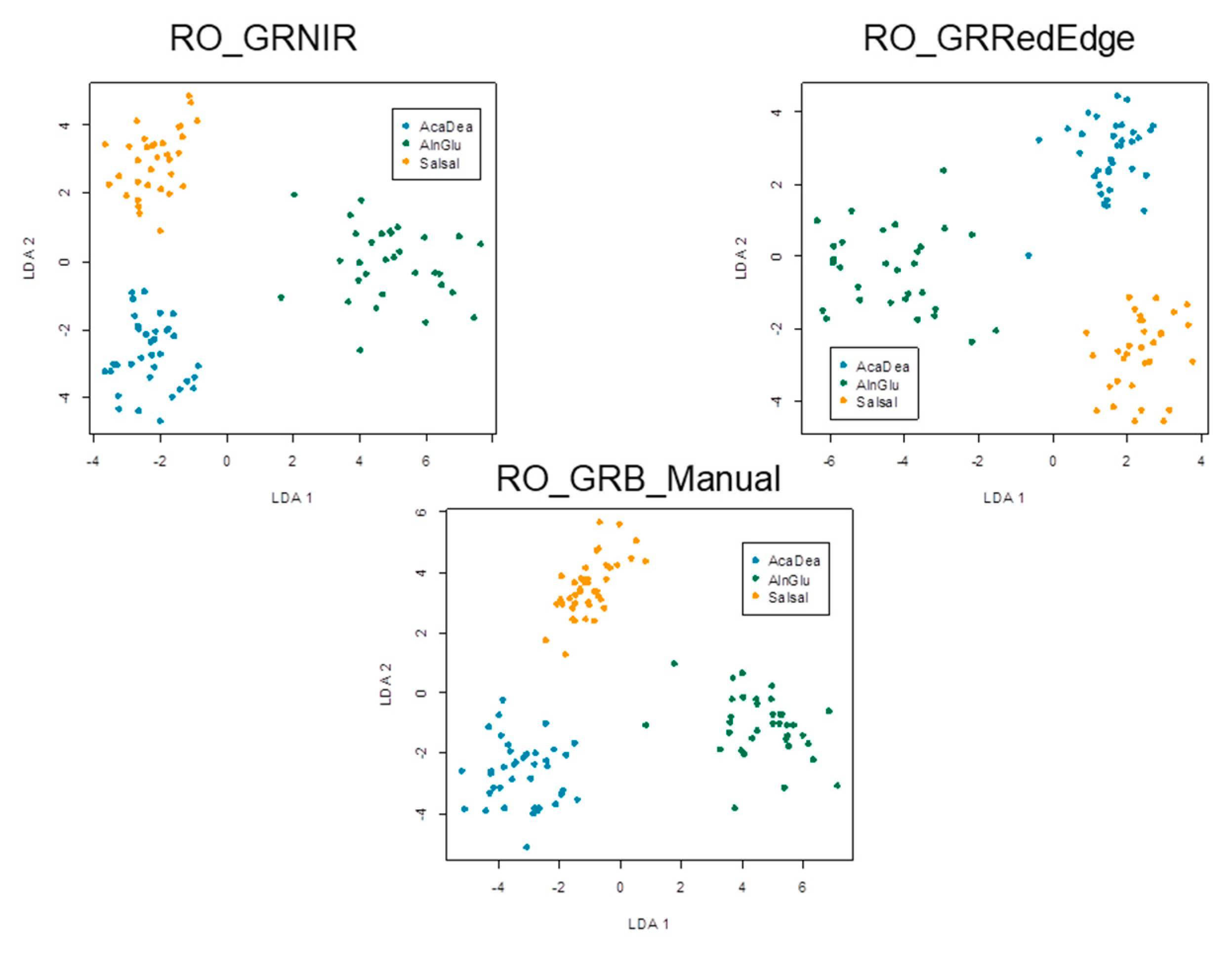
3.2. OBIA Species Classification and Tree AGB Maps
4. Discussion
4.1. Riparian Carbon Stocks
4.2. AGB Remote-Sensing Approach
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cierjacks, A.; Kleinschmit, B.; Babinsky, M.; Kleinschroth, F.; Markert, A.; Menzel, M.; Ziechmann, U.; Schiller, T.; Graf, M.; Lang, F. Carbon stocks of soil and vegetation on Danubian floodplain. J. Plant Nutr. Soil Sci. 2010, 173, 644–653. [Google Scholar] [CrossRef]
- Cartisano, R.; Mattioli, W.; Corona, P.; Mugnozza, G.S.; Sabatti, M.; Ferrari, B.; Cimini, D.; Giuliarelli, D. Assessing and mapping biomass potential productivity from poplar-dominated riparian forests: A case study. Biomass Bioenergy 2013, 54, 293–302. [Google Scholar] [CrossRef]
- Rieger, I.; Kowarik, I.; Cierjacks, A. Drivers of carbon sequestration by biomass compartment of riparian forests. Ecosphere 2015, 6, 185. [Google Scholar] [CrossRef]
- Sutfin, N.A.; Wohl, E.E.; Dwir, K.A. Banking carbon: A review of organic carbon storage and physical factors influencing retention in floodplains and riparian ecosystems. Earth Surf. Processes Landf. 2016, 41, 38–60. [Google Scholar] [CrossRef]
- Dybala, K.E.; Steger, K.; Walsh, R.G.; Smart, D.R.; Gardali, T.; Seavy, N.E. Optimizing carbon storage and biodiversity co-benefits in reforested riparian zones. J. Appl. Ecol. 2018, 56, 1–11. [Google Scholar] [CrossRef]
- Woodall, C.W.; Liknes, G.C. Climatic regions as an indicator of forest coarse and fine woody debris carbon stocks in the United States. Carbon Balance Manag. 2008, 3, 5. [Google Scholar] [CrossRef]
- Dybala, K.E.; Matzek, V.; Gardali, T.; Seavy, N.E. Carbon sequestration in riparian forests: A global synthesis and meta-analysis. Glob. Chang. Biol. 2019, 25, 57–67. [Google Scholar] [CrossRef]
- Dwire, K.A.; Kauffman, J.B.; Brookshire, E.N.J.; Baham, J.E. Plant biomass and species composition along an environmental gradient in montane riparian meadows. Oecologia 2004, 139, 309–317. [Google Scholar] [CrossRef]
- Giese, L.A.B.; Aust, W.M.; Kolka, R.K.; Trettin, C.C. Biomass and carbon pools of disturbed riparian forests. For. Ecol. Manag. 2003, 180, 493–508. [Google Scholar] [CrossRef]
- Grabe, M.; Kleber, M.; Hartmann, K.J.; Jahn, R. Preparing a soil carbon inventory of Saxony-Anhalt, Central Germany using GIS and the state soil data base SABO_P. J. Plant Nutr. Soil Sci. 2003, 166, 642–648. [Google Scholar] [CrossRef]
- Cabezas, A.; Comín, F.A. Carbon and nitrogen accretion in the topsoil of the Middle Ebro River Floodplains (NE Spain): Implications for their ecological restoration. Ecol. Eng. 2010, 36, 640–652. [Google Scholar] [CrossRef]
- Suchenwirth, L.; Forster, M.; Cierjacks, A.; Lang, F.; Kleinschmit, B. Knowledge-based classification of remote sensing data for the estimation of below- and above-ground organic carbon stocks in riparian forests. Wetl. Ecol. Manag. 2012, 20, 151–163. [Google Scholar] [CrossRef]
- Filipi, A.M.; Güneralp, I.; Randal, J. Hyperspectral remote sensing of aboveground biomass on a river meander bend using multivariate adaptive regression splines and stochastic gradient boosting. Remote Sens. Lett. 2014, 5, 432–441. [Google Scholar] [CrossRef]
- Güneralp, I.; Filippi, A.M.; Randall, J. Estimation of floodplain aboveground biomass using multispectral remote sensing and nonparametric modeling. Int. J. Appl. Earth Obs. Geoinf. 2014, 33, 119–126. [Google Scholar] [CrossRef]
- Mendez-Estrella, R.; Romo-Leon, J.R.; Castellanos, A. Mapping changes in carbon storage and productivity services provided by riparian ecosystems of semi-arid environments in Northwestern Mexico. ISPRS Int. J. Geoinf. 2017, 6, 298. [Google Scholar] [CrossRef]
- Suchenwirth, L.; Stumer, W.; Schmidt, T.; Forster, M.; Kleinschmit, B. Large-scale mapping of carbon stocks in riparian forest with self-organizing maps and the k-neighbor algorithm. Forests 2014, 5, 1635–1652. [Google Scholar] [CrossRef]
- Husson, E.; Lindgren, F.; Ecke, F. Assessing biomass and metal contents in riparian vegetation along a pollution gradient using an unmanned aircraft system. Water Air Soil Pollut. 2014, 225, 1957. [Google Scholar] [CrossRef]
- Goetz, S.; Baccini, A.; Laporte, N.; Johns, T.; Walker, W.; Kellndorfer, J.; Houghton, R.; Sun, M. Mapping and monitoring carbon stocks with satellite observations: A comparison of methods. Carbon Balance Manag. 2009, 4, 2. [Google Scholar] [CrossRef]
- Jucker, T.; Caspersen, J.; Chavé, J.; Antin, C.; Barbier, N.; Bongers, F.; Dalponte, M.; van Ewijk, K.Y.; Forrester, D.I.; Haeni, M.; et al. Allometric equations for integrating remote sensing imagery into forest monitoring programmes. Glob. Chang. Biol. 2017, 23, 177–190. [Google Scholar] [CrossRef]
- Matzek, V.; Stella, J.; Ropion, P. Development of a carbon calculator tool for riparian forest restoration. Appl. Veg. Sci. 2018, 1–11. [Google Scholar] [CrossRef]
- Barrachina, M.; Cristóbal, J.; Tulla, A.F. Estimating above-ground biomass on mountain meadows and pastures through remote sensing. Int. J. Appl. Earth Obs. Geoinf. 2015, 38, 184–192. [Google Scholar] [CrossRef]
- Congalton, R.G.; Birch, K.; Jones, R.; Schriever, J. Evaluating remotely sensed techniques for mapping riparian vegetation. Comput. Electron. Agric. 2002, 37, 113–126. [Google Scholar] [CrossRef]
- Dufour, S.; Bernez, I.; Betbeder, J.; Corgne, S.; Hubert-Moy, L.; Nabucet, J.; Rapinel, S.; Sawtschuk, J.; Trollé, C. Monitoring restored riparian vegetation: How can recent developments in remote sensing sciences help? Knowl. Manag. Aquat. Ecosyst. 2013, 410, 10. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Aguiar, F.C.; Silva, J.M.N.; Ferreira, M.T.; Pereira, J.M.C. Optimal attributes for the object based detection of giant reed in riparian habitats: A comparative study between Airborne High Spatial Resolution and WorldView-2 imagery. Int. J. Appl. Earth Obs. Geoinf. 2014, 32, 79–91. [Google Scholar] [CrossRef]
- Rango, A.; Laliberte, A.; Herrick, J.E.; Winters, C.; Havstad, K.; Steele, C.; Browning, D.N. Unmanned aerial vehicle-based remote sensing for rangeland assessment, monitoring and management. J. Appl. Remote Sens. 2009, 3, 1–15. [Google Scholar] [CrossRef]
- Dunford, R.; Michel, K.; Gagnage, M.; Piégay, H.; Trémelo, M.L. Potential and constraints of Unmanned Aerial Vehicle technology for the characterization of Mediterranean riparian forest. Int. J. Remote Sens. 2009, 30, 4915–4935. [Google Scholar] [CrossRef]
- Husson, E.; Hagner, O.; Ecke, F. Unmanned aircraft systems help to map aquatic vegetation. Appl. Veg. Sci. 2014, 567–577. [Google Scholar] [CrossRef]
- Michez, A.; Piégay, H.; Lisein, J.; Claessens, H.; Lejeune, P. Classification of riparian forest species and health condition using multi-temporal and hyperspatial imagery from unmanned aerial system. Environ. Monit. Assess. 2016, 188, 146. [Google Scholar] [CrossRef]
- Blaschke, T. Object based image analysis for remote sensing. J. Photogramm. Remote Sens. 2010, 65, 2–16. [Google Scholar] [CrossRef]
- Peña-Barragán, J.M.; Ngugi, M.K.; Plant, R.E.; Six, J. Object-based crop identification using multiple vegetation indices, textural features and crop phenology. Remote Sens. Environ. 2011, 115, 1301–1316. [Google Scholar] [CrossRef]
- Ninyerola, M.; Pons, X.; Roure, J.M. Atlas Climático Digital de la Península Ibérica. Metodología y Aplicaciones en Bioclimatología y Geobotánica; Universidad Autónoma de Barcelona: Bellaterra, Spain, 2005; ISBN 932860-8-7. [Google Scholar]
- Ravindranath, N.H.; Ostwald, M. Carbon Inventory Methods Handbook for Greenhouse Gas Inventory, Carbon Mitigation and Roundwood Production Projects; Advances in Global Change Research Book Series; Springer Netherlands: Dordrecht, The Netherlands, 2008; Volume 29. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Good Practice Guidance for Land Use, Land-Use Change and Forestry; Penman, J., Gytarsky, M., Hiraishi, T., Krug, T., Kruger, D., Pipatti, R., Buendia, L., Miwa, K., Ngara, T., Tanabe, K., et al., Eds.; Institute for Global Environmental Strategies: Hayama, Japan, 2003; Available online: http://www.ipcc-nggip.iges.or.jp (accessed on 21 November 2019).
- FAO. Measuring and modelling soil carbon stocks and stock changes in livestock production systems: Guidelines for assessment (Version 1). In Livestock Environmental Assessment and Performance (LEAP) Partnership; FAO: Rome, Italy, 2019; p. 170, Licence: CC BY-NC-SA 3.0 IGO. [Google Scholar]
- Smith, J.E.; Heath, L.S.; Hoover, C.M. Carbon factors and models for forest carbon estimates for the 2005-2011 National Greenhouse Gas Inventories of the United States. For. Ecol. Manag. 2013, 307, 7–19. [Google Scholar] [CrossRef]
- Guerra-Hernández, J.; González-Ferreiro, E.; Sarmento, A.; Silva, J.; Nunes, A.; Correia, A.; Fontes, L.; Tomé, M.; Díaz-Varela, R. Using high resolution UAV imagery to estimate tree variables in Pinus pinea plantation in Portugal. For. Syst. 2016, 25, 1–5 eSC09. [Google Scholar] [CrossRef]
- Lu, B.; He, Y. Species classification using Unmanned Aerial Vehicle (UAV)-acquired high spatial resolution imagery in a heterogeneous grassland. ISPRS J. Photogramm. Remote Sens. 2017, 128, 73–85. [Google Scholar] [CrossRef]
- Hakala, T.; Markelin, L.; Honkavaara, E.; Scott, B.; Theocharous, T.; Nevalainen, O.; Näsi, R.; Suomalainen, J.; Viljanen, N.; Greenwell, C.; et al. Direct reflectance measurements from drones: Sensor absolute radiometric calibration and system tests for forest reflectance characterization. Sensors 2018, 18, 1417. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Phinn, S.; Johansen, K.; Robson, A. Assessing radiometric correction approaches for Multi-Spectral UAS imagery for horticultural applications. Remote Sens. 2018, 10, 1684. [Google Scholar] [CrossRef]
- Ke, Y.; Quackenbush, L.J.; Im, J. Synergistic use of QuickBird multispectral imagery and LIDAR data for object-based forest species classification. Remote Sens. Environ. 2010, 114, 1141–1154. [Google Scholar] [CrossRef]
- Comaniciu, D.; Meer, P. Mean shift: A robust approach toward feature space analysis. IEEE Trans. Pattern Anal. Mach. Intell. 2002, 24, 603–619. [Google Scholar] [CrossRef]
- Michel, J.; Youssefi, D.; Grizonnet, M. Stable mean-shift algorithm and its application to the segmentation of arbitrarily large remote sensing images. IEEE Trans. Geosci. Remote Sens. 2015, 53, 952–964. [Google Scholar] [CrossRef]
- Benz, U.C.; Hofmann, P.; Willhauck, G.; Lingenfelder, I.; Heynen, M. Multiresolution, object-oriented fuzzy analysis of remote sensing data for GIS-ready information. ISPRS J. Photogramm. Remote Sens. 2004, 58, 239–258. [Google Scholar] [CrossRef]
- Laliberté, A.S.; Rango, A.; Havstad, K.M.; Paris, J.F.; Beck, R.F.; McNeely, R.; Gonzalez, A.L. Object-oriented image analysis for mapping shrub encroachment from 1937 to 2003 in southern New Mexico. Remote Sens. Environ. 2004, 93, 198–210. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a green channel in remote sensing of global vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Merzlyak, M.N. Remote sensing of chlorophyll concentration in higher plant leaves. Adv. Space Res. 1998, 22, 689–692. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. Plant Pathol. J. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Dash, J.; Huang, W.; Peng, D.; Qin, Q.; Mortimer, H.; Casa, R.; Pignatti, S.; Laneve, G.; Pascucci, S.; et al. Vegetation indices. Combining the red and red-edge spectral information for leaf area index retrieval. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2018, 1–12, 1482–1493. [Google Scholar] [CrossRef]
- Cerdeira, J.; Duarte Silva, P.; Cadima, J.; Minhoto, M. Subselect: Selecting Variable Subsets. R Package Version 0.14. Available online: https://CRAN.R-project.org/package=subselect (accessed on 21 October 2019).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Kuhn, M.; Wing, J.; Weston, S.; Williams, A.; Keefer, C.; Engelhardt, A.; Cooper, T.; Mayer, Z.; Kenkel, B.; The R Core Team; et al. Caret: Classification and Regression Training. R Package Version 6.0–84. 2019. Available online: https://CRAN.R-project.org/package=caret (accessed on 21 November 2019).
- McGarigal, K.; Marks, B.J. FRAGSTATS. Spatial Pattern Analysis Program for Quantifying Landscape Structure. Version 2.0; Forest Science Department, Oregon State University: Corvallis, OR, USA, 1994. [Google Scholar]
- Correia, A.C.; Tomé, M.; Pacheco, C.A.; Faias, S.; Dias, A.C.; Freire, J.; Carvalho, P.O.; Pereira, J.S. Biomass allometry and carbon factors for a Mediterranean pine (Pinus pinea L.) in Portugal. For. Syst. 2010, 19, 418–433. [Google Scholar] [CrossRef]
- Correia, A.C.; Costa e Silva, F.; Dubbert, M.; Piayda, A.; Pereira, J.S. Severe dry winter affects plant phenology and carbon balance of a cork oak woodland understorey. Acta Oecol. 2016, 26, 1–12. [Google Scholar] [CrossRef]
- Soares, P.; Tomé, M.; Pereira, J.S.A. Produtividade do eucaliptal. In Impactes Ambientais do Eucaliptal em Portugal; Alves, A.A.M., Pereira, J.S., Silva, e.J.M.N., Eds.; ISAPress: Lisboa, Portugal, 2007; ISBN 978-972-8669-25-6. [Google Scholar]
- Mitra, S.; Wassmann, R.; Vlek, P.L. An appraisal of global wetland area and its organic carbon stock. Curr. Sci. 2005, 88, 25–35. [Google Scholar]
- Ruffing, C.; Dwire, K.A.; Daniels, M.D. Carbon pools in stream riparian corridors; legacy of disturbance along mountain streams of southeastern Wyoming. Earth Surf. Processes Landf. 2016, 41, 208–223. [Google Scholar] [CrossRef]
- Norton, J.B.; Jungst, L.J.; Norton, U.; Olsen, H.R.; Tate, K.W.; Horwath, W.R. Soil carbon and nitrogen storage in upper montane riparian meadows. Ecosystems 2011, 14, 1217–1231. [Google Scholar] [CrossRef]
- Aguiar, F.C.; Ferreira, M.T.; Albuquerque, A.; Moreira, I. Alien and endemic flora on reference and non-reference sites from Mediterranean type-streams of Portugal. Aquat. Conserv. 2007, 17, 335–347. [Google Scholar] [CrossRef]
- Lu, D. Aboveground biomass estimation using Landsat TM data in the Brazilian Amazon. Int. J. Remote Sens. 2005, 26, 2509–2525. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Description | Calculation | Reference |
|---|---|---|---|
| Mean (AVGBAND_x) | Basic spectral statistics | Average value of the pixels comprising the object in each band (Green, Red, RedEdge, NIR) | Barrachina et al., 2015 [21] |
| Normalized Difference Vegetation Index (NDVI) | Greenness level. High values for green vegetation | Barrachina et al., 2015 [21] | |
| Green Normalized Difference Vegetation Index (GNDVI) | Greenness level. More sensitive to variations of chlorophyll contents than NDVI | Gitelson et al., 1996; 1998 [45,46] | |
| Red Edge Normalized Difference Vegetation Index (RedEdgeNDVI) | Greenness level. Less prone to saturation | Xie et al., 2018 [48] | |
| Green Chlorophyll Index (GCI) | Greenness level. More sensitive to variations of chlorophyll contents and a higher signal-to-noise ratio. | Xie et al., 2018 Gitelson et al., 2003 [47] | |
| Red Edge Chlorophyll Index (RedEdgeCI) | Greenness level. Less prone to saturation. | Xie et al., 2018 [48] | |
| Modified Simple Ration (MSR) | Greenness level. Proposed to suppress saturation issues, | Xie et al., 2018 [48] | |
| Red Edge Modified Simple Ration (RedEdgeMSR) | Xie et al., 2018 [48] |
| Reference (RO_GRNIR) | Reference (RO_GRRedEdge) | Reference (RO_RBG_Manual) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Prediction | AcaDea | AlnGlu | SalSal | AcaDea | AlnGlu | SalSal | AcaDea | AlnGlu | SalSal |
| AcaDea | 34.4 | 0.4 | 0 | 34.5 | 1.4 | 0 | 32.7 | 0.4 | 0 |
| AlnGlu | 0 | 30.9 | 0 | 0 | 30.9 | 0 | 0 | 30.7 | 0 |
| SalSal | 0 | 0.1 | 34.2 | 0 | 0.3 | 33.2 | 0 | 0.6 | 35.6 |
| Accuracy (average) | 99.5 | 98.3 | 99.0 | ||||||
| 95% | 99% | 99.9% | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Nump (N) | MPS (m2) | CA (%) | Nump (N) | MPS (m2) | CA (%) | Nump (N) | MPS (m2) | CA (%) | |
| AcaDea | 5961 | 4.2 ± 0.3 | 22.3 | 7823 | 3.9 ± 0.2 | 27.3 | 9645 | 3.7 ± 0.2 | 32.1 |
| AlnGlu | 5509 | 4.3 ± 0.4 | 20.9 | 7380 | 4.0 ± 0.3 | 26.9 | 8727 | 3.8 ± 0.3 | 29.8 |
| SalSal | 3694 | 3.0 ± 0.2 | 9.9 | 5249 | 2.6 ± 0.2 | 12.1 | 6888 | 2.3 ± 0.1 | 14.2 |
| Unclass | 27187 | 1.9 ± 0.1 | 46.9 | 21899 | 1.7 ± 0.1 | 33.7 | 17091 | 1.5 ± 0.0 | 23.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, M.R.; Aguiar, F.C.; Martins, M.J.; Rico, N.; Ferreira, M.T.; Correia, A.C. Carbon Stock Estimations in a Mediterranean Riparian Forest: A Case Study Combining Field Data and UAV Imagery. Forests 2020, 11, 376. https://doi.org/10.3390/f11040376
Fernandes MR, Aguiar FC, Martins MJ, Rico N, Ferreira MT, Correia AC. Carbon Stock Estimations in a Mediterranean Riparian Forest: A Case Study Combining Field Data and UAV Imagery. Forests. 2020; 11(4):376. https://doi.org/10.3390/f11040376
Chicago/Turabian StyleFernandes, Maria Rosário, Francisca C. Aguiar, Maria João Martins, Nuno Rico, Maria Teresa Ferreira, and Alexandra C. Correia. 2020. "Carbon Stock Estimations in a Mediterranean Riparian Forest: A Case Study Combining Field Data and UAV Imagery" Forests 11, no. 4: 376. https://doi.org/10.3390/f11040376
APA StyleFernandes, M. R., Aguiar, F. C., Martins, M. J., Rico, N., Ferreira, M. T., & Correia, A. C. (2020). Carbon Stock Estimations in a Mediterranean Riparian Forest: A Case Study Combining Field Data and UAV Imagery. Forests, 11(4), 376. https://doi.org/10.3390/f11040376