Micro-Hotspots for Conservation: An Umbrella Tree Species for the Unique Socotran Reptile Fauna

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites and Sampling
2.2. Data Analyses
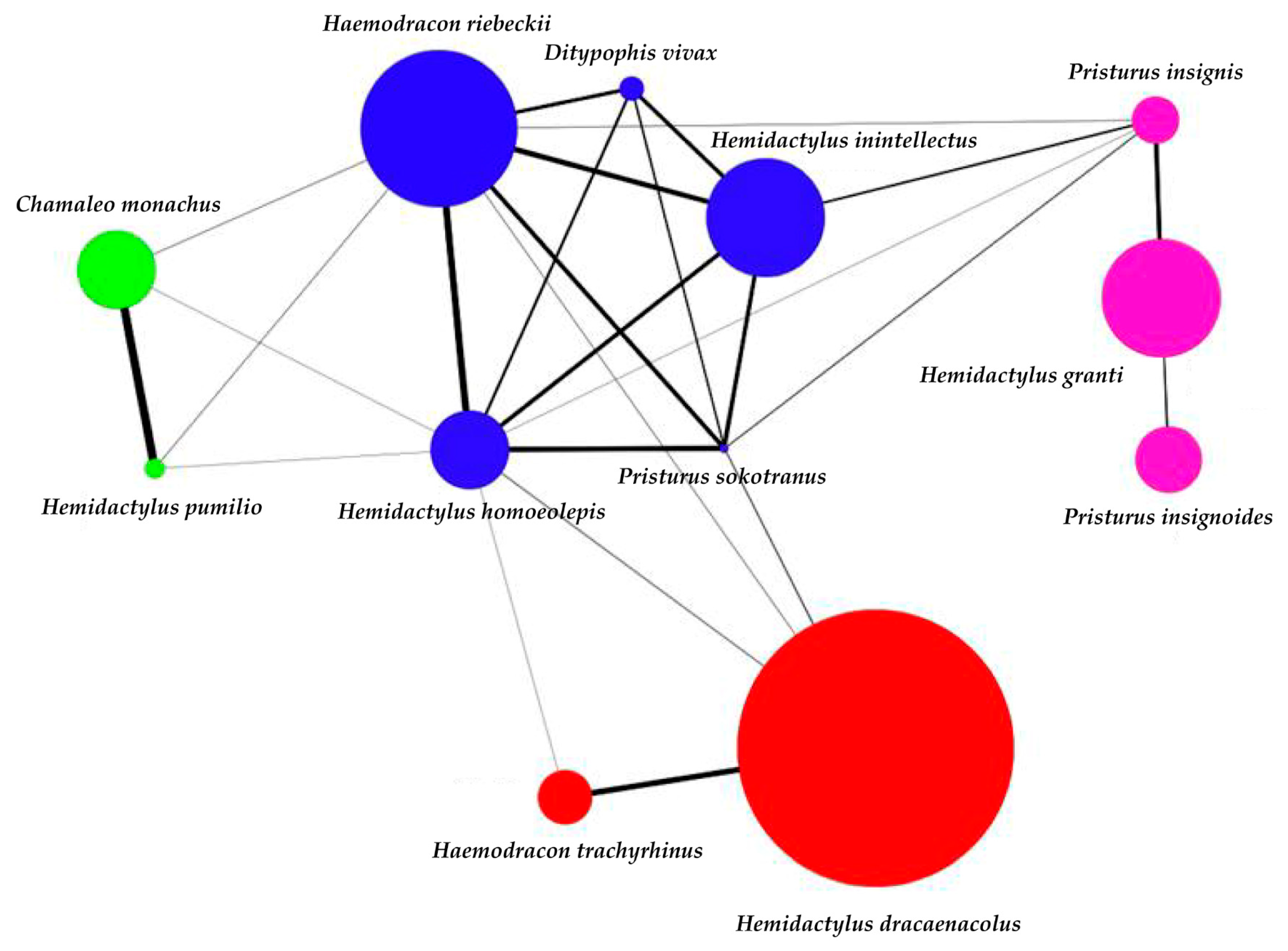
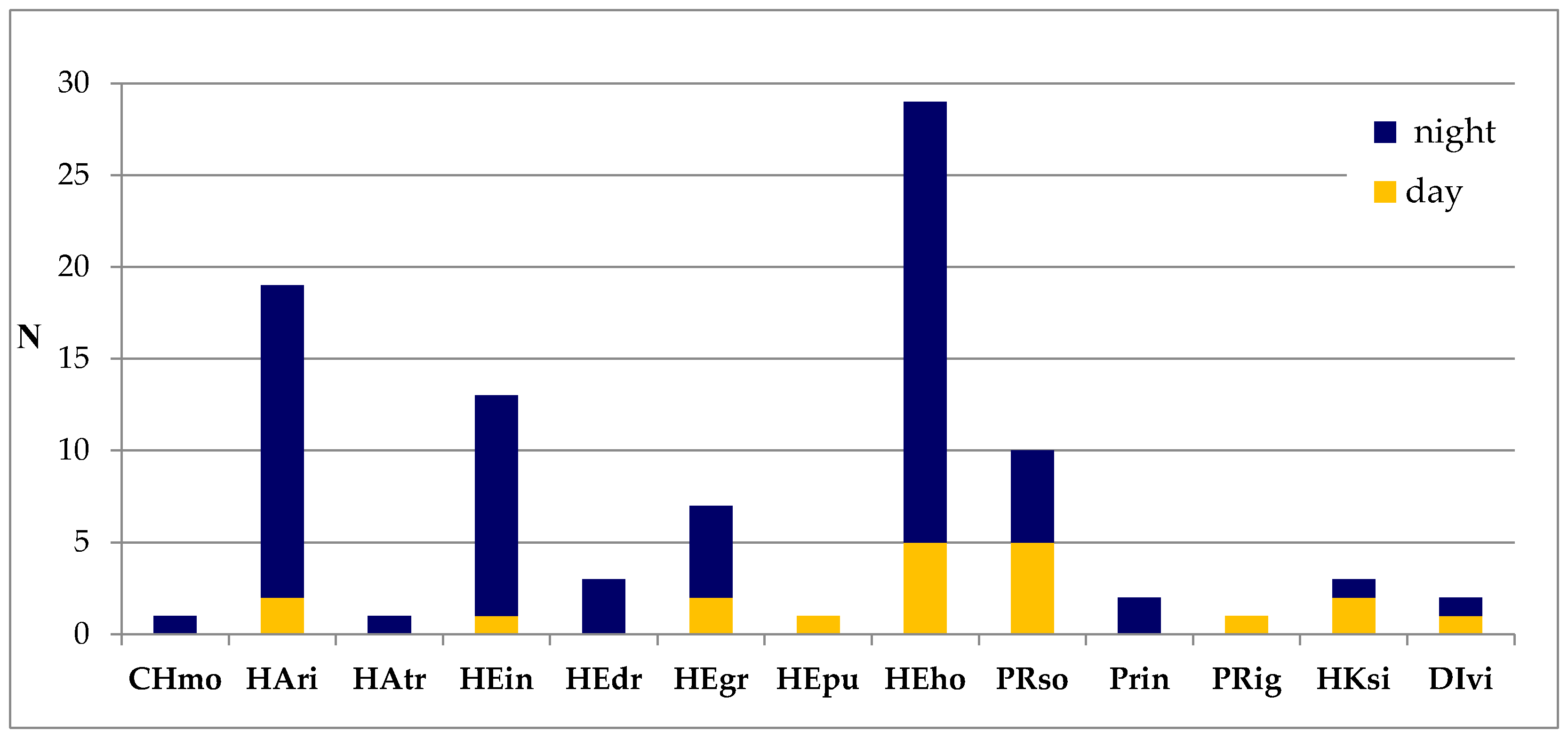
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Species | Codes | Family | IUCN | Distribution | Sites |
|---|---|---|---|---|---|
| Chamaeleo monachus | CHmo | Chamaeleonidae | NT | Socotra Island | 1 |
| Haemodracon riebeckii | HAri | Phyllodactylidae | LC | Socotra, Samha | 8 |
| Haemodracon trachyrhinus | HAtr | Phyllodactylidae | LC | Socotra Island | 1 |
| Hemidactylus dracaenacolus | HEdr | Gekkonidae | CR | Socotra Island | 2 |
| Hemidactylus granti | HEgr | Gekkonidae | NT | Socotra Island | 2 |
| Hemidactylus homoeolepis | HEho | Gekkonidae | LC | Socotra, Samha & Darsa | 9 |
| Hemidactylus inintellectus | HEin | Gekkonidae | LC | Socotra Island | 5 |
| Hemidactylus pumilio | HEpu | Gekkonidae | LC | Socotra Island | 1 |
| Pristurus insignoides | PRig | Sphaerodactylidae | LC | Socotra Island | 2 |
| Pristurus insignis | PRin | Sphaerodactylidae | LC | Socotra Island | 1 |
| Pristurus sokotranus | PRso | Sphaerodactylidae | LC | Socotra Island | 7 |
| Ditypophis vivax | DIvi | Pseudoxyrhophiidae | LC | Socotra Island | 2 |

References
- Lambeck, R.J. Focal species: A multi-species umbrella for nature conservation. Conserv. Biol. 1997, 11, 849–856. [Google Scholar] [CrossRef]
- Fleishman, E.; Murphy, D.D.; Brussard, P.F. A new method for selection of umbrella species for conservation planning. Ecol. Appl. 2000, 10, 569–579. [Google Scholar] [CrossRef]
- Roberge, J.; Angelstam, P. Usefulness of the umbrella species concept as a conservation tool. Conserv. Biol. 2004, 18, 76–85. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global biodiversity conservation?: The critical role of hotspots. In Biodiversity Hotspots Distribution and Protection of Conservation Priority Areas; Zachos, F.E., Habel, J.C., Eds.; Springer-Verlag: Basel, Switzerland, 2012; pp. 3–22. [Google Scholar]
- Courchamp, F.; Hoffmann, B.D.; Russell, J.C.; Leclerc, C.; Bellard, C. Climate change, sea-level rise, and conservation: Keeping island biodiversity afloat. Trends Ecol. Evol. 2014, 29, 127–130. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton Univ. Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Losos, J.B.; Ricklefs, R.E. The Theory of Island Biogeography Revisited; Princeton Univ. Press: Princeton, NJ, USA, 2009. [Google Scholar]
- Autin, J.; Bellahsen, N.; Leroy, S.; Husson, L.; Beslier, M.-O.; d’Acremont, E. The role of structural inheritance in oblique rifting: Insights from analogue models and application to the Gulf of Aden. Tectonophysics 2013, 607, 51–64. [Google Scholar] [CrossRef]
- Miller, A.G.; Morris, M. Ethnoflora of the Soqotra Archipelago; The Royal Botanic Garden: Edinburgh, Scotland, 2004. [Google Scholar]
- Van Damme, K. Socotra Archipelago. In Encyclopedia of Islands; Gillespie, R., Clague, D., Eds.; Univ. California: Berkeley, CA, USA; Los Angeles, CA, USA, 2009; pp. 846–851. [Google Scholar]
- Razzetti, E.; Sindaco, R.; Griego, C.; Pella, F.; Ziliani, U.; Pupin, F.; Riservato, E.; Pelliteri-Rosa, D.; Butikofer, L.; Saeed Suleiman, A.; et al. Annotated checklist and distribution of the Socotran Archipelago Herpetofauna (Reptilia). Zootaxa 2011, 2826, 1–44. [Google Scholar] [CrossRef]
- Sindaco, R.; Metallinou, M.; Pupin, F.; Fasola, M.; Carranza, S. Forgotten in the ocean: Systematics, biogeography and evolution of the Trachylepis skinks of the Socotra Archipelago. Zool. Scr. 2012, 41, 346–362. [Google Scholar] [CrossRef]
- Vasconcelos, R.; Carranza, S. Systematics and biogeography of Hemidactylus homoeolepis Blanford, 1881, with the description of a new species from Arabia. Zootaxa 2014, 3835, 501–527. [Google Scholar] [CrossRef]
- Schätti, B.; Utiger, U. Hemerophis, a new genus for Zamenis socotrae Günther, and a contribution to the phylogeny of Old World racers, whip snakes, and related genera (Reptilia: Squamata: Colubrinae). Rev. Suisse Zool. 2001, 108, 919–948. [Google Scholar] [CrossRef]
- Nagy, Z.T.; Joger, H.; Wink, M.; Glaw, F.; Vences, M. Multiple colonization of Madagascar and Socotra by colubrid snakes: Evidence from nuclear and mitochondrial gene phylogenies. Proc. R. Soc. B 2003, 270, 2613–2621. [Google Scholar] [CrossRef]
- Gómez-Díaz, E.; Sindaco, R.; Pupin, F.; Fasola, M.; Carranza, S. Origin and in situ diversification in Hemidactylus geckos of the Socotra Archipelago. Mol. Ecol. 2012, 21, 4074–4092. [Google Scholar] [CrossRef] [PubMed]
- Miller, A. Dracaena Cinnabari. The IUCN Red List of Threatened Species 2004: E.T30428A9548491. 2004. Available online: https://dx.doi.org/10.2305/IUCN.UK.2004.RLTS.T30428A9548491.en (accessed on 17 January 2020).
- Adolt, R.; Habrova, H.; Madera, P. Crown age estimation of a monocotyledonous tree species Dracaena cinnabari using logistic regression. Trees 2012, 26, 1287–1298. [Google Scholar] [CrossRef]
- Maděra, P.; Volařík, D.; Patočka, Z.; Kalivodová, H.; Divín, J.; Rejžek, M.; Vybíral, J.; Lvončík, S.; Jeník, D.; Hanáček, P.; et al. Sustainable Land Use Management Needed to Conserve the Dragon’s Blood Tree of Socotra Island, a Vulnerable Endemic Umbrella Species. Sustainability 2019, 11, 3557. [Google Scholar] [CrossRef]
- Stone, L.; Roberts, A. The checkerboard score and species distributions. Oecologia 1990, 85, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Gotelli, N.J.; Entsminger, G.L. EcoSim: Null models Software for Ecology; Version 7.0; Acquired Intelligence Inc.; Kesey-Bear, 2001; Available online: http://homepages.together.net/~gentsmin/ecosim.htm (accessed on 17 January 2018).
- Blondel, V.D.; Guillaume, J.-L.; Lambiotte, R.; Lefebvre, E. Fast unfolding of communities in large networks. J. Stat. Mech. Theory Exp. 2008, 10, P10008. [Google Scholar] [CrossRef]
- Batagelj, V.; Mrvar, A. Pajek 2.05. Program for Analysis and Visualization of Large Networks. 2001. Available online: http://mrvar.fdv.uni-lj.si/pajek/be2.htm (accessed on 17 January 2018).
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. 2009. Available online: https://gephi.org/publications/gephi-bastian-feb09.pdf (accessed on 17 January 2018).
- De la Cruz-Francisco, V. Rhizophora mangle Linnaeus, 1753 as umbrella species and biological reason for the protection and restoration of Tampamachoco lagoon, Veracruz, Mexico. BIOCYT 2012, 5, 341–352. [Google Scholar]
- Vasconcelos, R.; Montero-Mendieta, S.; Simó-Riudalbas, M.; Sindaco, R.; Santos, X.; Fasola, M.; Llorente, G.; Razzetti, E.; Carranza, S. Unexpectedly high levels of cryptic diversity uncovered by a complete DNA barcoding of reptiles of the Socotra Archipelago. PLoS ONE 2016, 11, e0149985. [Google Scholar] [CrossRef]
- Vasconcelos, R.; Razgour, O.; Tarroso, P.; Fasola, M.; Carranza, S.; Alves, P.C. Combining molecular and landscape tools for targeting evolutionary processes in reserve design: An approach for islands. PLoS ONE 2018, 13, e0200830. [Google Scholar] [CrossRef]
- García, C.; Vasconcelos, R. The beauty and the beast: Endemic mutualistic interactions promote community-based conservation on Socotra Island (Yemen). J. Nat. Conserv. 2017, 35, 20–23. [Google Scholar] [CrossRef]
- Dormann, C.F.; Bobrowski, M.; Dehling, D.M.; Harris, D.J.; Hartig, F.; Lischke, H.; Moretti, M.D.; Pagel, J.; Pinkert, S.; Schleuning, M.; et al. Biotic interactions in species distribution modelling: 10 questions to guide interpretation and avoid false conclusions. Glob. Ecol. Biogeogr. 2018, 27, 1004–1016. [Google Scholar] [CrossRef]
- Adolt, R.; Pavlis, J. Age structure and growth of Dracaena cinnabari populations on Socotra. Trees 2004, 18, 43–53. [Google Scholar] [CrossRef]
- Attorre, F.; Francesconi, F.; Taleb, N.; Scholte, P.; Saeed, A.; Alfo, M.; Bruno, F. Will dragonblood survive the next period of climate change? Current and future potential distribution of Dracaena cinnabari (Socotra, Yemen). Biol. Conserv. 2007, 138, 430–439. [Google Scholar] [CrossRef]


| S | Lat | Long | Location | N | MD1 | XD3 | G | SH | BE |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 12.572 | 54.048 | Adho Di Meleh, Haggeher | 6 | 1500 | 1500 | 117 | 272 | 4.6 |
| 2 | 12.572 | 54.049 | Ba’a, 2 km NE of Diksam | 7 | 125 | 1360 | 158 | 281 | 4.8 |
| 3 | 12.467 | 54.002 | Qafshifo, 3 km SE Firmihin | 18 | 410 | 1186 | 246 | 288 | 7.2 |
| 4 | 12.476 | 54.017 | Firmihin Protected area | 12 | 1500 | 1500 | 226 | 336 | 5.4 |
| 5 | 12.485 | 54.008 | Haif, Firmihin | 4 | 345 | 1112 | 160 | 306 | 4.8 |
| 6 | 12.484 | 53.913 | Shibehon plateau | 4 | 262 | 1414 | 175 | 246 | 5.4 |
| 7 | 12.589 | 54.305 | Homhil, Hallah | 9 | 140 | 1342 | 225 | 290 | 5.6 |
| 8 | 12.475 | 54.149 | Di Gisfo, Wadi Di-Fa’rhroh | 8 | 192 | 1156 | 173 | 275 | 5.9 |
| 9 | 12.533 | 54.315 | Killisan, 3km NW Qademinoh | 17 | 150 | 1286 | 207 | 268 | 5.1 |
| 10 | 12.500 | 53.987 | Diksam, Qafshifo | 4 | 65 | 1269 | 160 | 333 | 5.6 |
| 11 | 12.565 | 54.013 | Skand, Haggeher | 2 | 233 | 1191 | 114 | 260 | 4.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasconcelos, R.; Pujol-Buxó, E.; Llorente, G.A.; Saeed, A.; Carranza, S. Micro-Hotspots for Conservation: An Umbrella Tree Species for the Unique Socotran Reptile Fauna. Forests 2020, 11, 353. https://doi.org/10.3390/f11030353
Vasconcelos R, Pujol-Buxó E, Llorente GA, Saeed A, Carranza S. Micro-Hotspots for Conservation: An Umbrella Tree Species for the Unique Socotran Reptile Fauna. Forests. 2020; 11(3):353. https://doi.org/10.3390/f11030353
Chicago/Turabian StyleVasconcelos, Raquel, Eudald Pujol-Buxó, Gustavo A. Llorente, Ahmed Saeed, and Salvador Carranza. 2020. "Micro-Hotspots for Conservation: An Umbrella Tree Species for the Unique Socotran Reptile Fauna" Forests 11, no. 3: 353. https://doi.org/10.3390/f11030353
APA StyleVasconcelos, R., Pujol-Buxó, E., Llorente, G. A., Saeed, A., & Carranza, S. (2020). Micro-Hotspots for Conservation: An Umbrella Tree Species for the Unique Socotran Reptile Fauna. Forests, 11(3), 353. https://doi.org/10.3390/f11030353