Abstract
Many bark and ambrosia beetle species (Coleoptera: Scolytinae and Platypodinae) are known to have spread worldwide in relation to international trade. Concerns have been expressed within the European and Mediterranean Plant Protection Organization (EPPO) about recent introductions of non-indigenous species of these groups. Regulation of the non-coniferous wood trade into many EPPO member countries is currently not sufficient to cover such risks. In 2018–2019, an EPPO study on the risk of bark and ambrosia beetles associated with imported non-coniferous wood was carried out, and the key characteristics contributing to the pest risk from introduced species were determined using expert consensus. This paper summarizes the key findings of the study, which are available in full detail on the EPPO website. The study identified biological and other risk factors and illustrated them with examples from 26 beetle species or groups of species known to be invasive or posing a threat to plant health. These representative species were classified into three categories based on known damage and level of uncertainty. In the present article, factorial discriminant analyses were used to identify features of bark and ambrosia beetle biology associated with damage caused and invasiveness. Based on the information assembled and consideration of the risk factors, it was recommended that in order to prevent the introduction of new bark and ambrosia beetles via non-coniferous wood commodities, horizontal phytosanitary measures should be adopted, irrespective of the host plant species and the origin (i.e., for all genera of non-coniferous woody plants and from all origins). Phytosanitary measures are presented here for various wood commodities.
Keywords:
Scolytinae; Platypodinae; quarantine; pest risk; biosecurity; invasive species; plant health 1. Introduction
Bark beetles and ambrosia beetles belong to two weevil subfamilies of Curculionidae: Scolytinae and Platypodinae. There are more than 6000 species of Scolytinae described to date, and the majority are tropical or subtropical species. About 2100 species of Scolytinae are ambrosia beetles [1], the remaining species are mostly bark beetles. The second subfamily Platypodinae includes around 1400 additional ambrosia beetle species [2]. Bark beetles feed on a variety of tree tissues, from bark to phloem, sapwood, and xylem. Ambrosia beetles live in the xylem where they cultivate symbiotic fungi on which they feed.
Bark and ambrosia beetles have attracted a lot of attention in the past decades following numerous introductions (the present article uses the terminology of the Glossary of Phytosanitary Terms of the International Plant Protection Convention [3]. In particular, ’introduction’ is defined as “the entry of a pest resulting in its establishment”) worldwide, and, in some cases, massive damage. Many Scolytinae and Platypodinae are known to have spread worldwide in relation to international trade. Recently, new non-indigenous species of bark and ambrosia beetles, not listed specifically as quarantine pests, have been introduced into the European and Mediterranean Plant Protection Organization (EPPO) region ([4,5,6]). EPPO is an intergovernmental organisation responsible for cooperation in plant health within the Euro-Mediterranean region. It has 52 member countries to date. Through the work of expert groups and decision-making bodies, recommendations are made to member countries in relation to plant protection products and phytosanitary regulations. One component of EPPO’s phytosanitary work is identifying pest risks and recommending pest risk management options. The identification of pest risks by EPPO and subsequent recommendations for regulation are typically based on a pest-specific approach, involving horizon-scanning, followed by species-specific pest risk analysis (PRA). This PRA approach identified several non-coniferous bark and ambrosia beetles presenting a potential risk, which were added to various EPPO lists (Table 1) or published in the EPPO Reporting Service.

Table 1.
Non-coniferous bark and ambrosia beetles on European and Mediterranean Plant Protection Organization (EPPO) lists (as of March 2020).
In 2017, the EPPO Panels on Phytosanitary Measures and on Quarantine Pests for Forestry expressed concerns about recent introductions of bark and ambrosia beetle species of non-coniferous wood. Indeed, the regulation of the non-coniferous wood trade into many EPPO member countries is currently not considered sufficient to cover such risks. All non-European Scolytinae attacking coniferous wood are regulated in the European Union and in some other non-EU EPPO countries, irrespective of their individual status on quarantine lists. However, such regulation (targeting the commodity—wood—and not the organisms themselves) does not exist for non-coniferous wood, and would be justified by high uncertainty about which species might be introduced into the region, and a lack of knowledge on the following:
- The ecology and impact of individual species such as host range and damage at origin, ability to attack new host species in new locations, associated fungi, and climatic adaptability;
- Unexpected performance of the beetle species in case of introduction into a new area. In particular, the novel host range and extent of damage in new areas can be predicted only with sufficient knowledge of each species’ native biology, which is not available for many species [7]. Moreover, the risk for different climatic zones in the EPPO region is difficult to assess. Several subtropical and tropical species have successfully established in the Mediterranean part of the EPPO region in recent years; and
- Pathways such as data on the trade of wood, which is mostly unavailable at the species and genus level in the EPPO region.
Due to this lack of knowledge, PRAs on individual species would in most cases contain major uncertainties, especially with regard to entry, establishment, and impact.
The EPPO panels determined that phytosanitary measures should be identified that are relevant for all species (from here onward ‘horizontal measures’) that may be appropriate to prevent further introductions of bark and ambrosia beetles associated with non-coniferous wood. The study presented here was therefore established based on representative bark and ambrosia beetle species of non-coniferous wood, which are not present in the EPPO region or are present but not widely distributed, for which such horizontal measures may be developed to cover the risk of all pests in these two groups.
The study was conducted between 2018 and 2019. The main outcomes are presented in this article, which provides a summary. All details can be found in the EPPO study on the risk of bark and ambrosia beetles associated with imported non-coniferous wood [8]. All statements in the study are supported by references to the literature. As this is a short summary, only very specific references are presented here, and all others are in the study. The objective of the article is to draw the attention of a wider audience than plant health regulators (who are the traditional EPPO partners) to this study, and to the challenges faced by plant health regulators when designing phytosanitary measures.
2. Methods and Definitions
The methodology was twofold. The EPPO study was carried out in 2018 by an EPPO Expert Working Group (EWG, authors of this article), with the participation of other experts on bark and ambrosia beetles and on pest risk analysis. The conclusions of the EWG were agreed upon by consensus. To corroborate the qualitative consensus building, and as a complement to the published EPPO study [8], factorial discriminant analyses were used to identify features of bark and ambrosia beetle biology associated with damage and invasiveness, hereafter called “risk factors”.
2.1. The Process of the European and Mediterranean Plant Protection Organization (EPPO) Expert Working Group
2.1.1. Selection of Representative Bark and Ambrosia Beetle Species
In order to identify key factors that influence the potential risks of successful entry, establishment, spread, and impact of bark and ambrosia beetles, an across-species approach was followed. Bark and ambrosia beetle species considered to present a potential risk to plant health in their native or introduced ranges were identified through expert knowledge or other sources (literature/databases). Species for which sufficient data were available to illustrate apparent risk factors were then selected. For each species, a pest information sheet was prepared based on the published literature including consideration of the potential risks for the EPPO region. Information on individual species was assembled mostly at the start of 2018, and no data were updated after September 2018.
2.1.2. Identification of Risk Factors and Categories
The main characteristics of bark and ambrosia beetles that contribute to their pest risk were analysed. The risk factors identified were detailed and illustrated with examples of the representative species.
All species were then summarised in case studies and classified into three categories:
- Known substantial damage;
- Indications of damage, with some uncertainties; and
- No documented damage, but several potential risk factors exist.
These categories were developed to take account of the known damage of each species and the uncertainties attached to pest risk. Expert judgement was used to allocate each species to a category.
2.1.3. Consideration of Possible Horizontal Measures
Possible horizontal measures were explored, and what coverage would be appropriate in terms of bark and ambrosia beetles, non-coniferous species, commodities, and commodity origins. Discussions were based on a review of the recommendations included in the International Standard for Phytosanitary Measures (ISPM) ISPM 39 International movement of wood [9] and risk management measures included in existing EPPO Standards and PRAs.
2.1.4. Definitions: Wood Commodities
For the purpose of studying entry pathways for representative species and identifying possible horizontal measures, the wood commodities were organized in three broad categories, which together comprise all the more detailed commodities defined in the EPPO study on wood commodities other than round wood, sawn wood, and manufactured items [10].
- Wood (all commodities of round wood and sawn wood, with or without bark)
- Wood chips, hogwood (“wood with or without bark in the form of pieces of varying particle size and shape, produced by crushing with blunt tools such as rollers, hammers, or flails” [10]), and processing wood residues (except sawdust and shavings);
- Bark (including for mulch, and other intended uses).
It should be noted that these commodities can be used as firewood.
The study did not consider measures for wood packaging material because these are already covered in ISPM 15 Regulation of wood packaging material in international trade [11]. When collecting information on representative bark and ambrosia beetle species, any known association with wood packaging material was nevertheless noted. Other wood commodities (e.g., sawdust and shavings, processed wood material) are considered to present a negligible risk based on EPPO [10] and previous EPPO PRAs on bark beetles.
Finally, other major pathways for individual representative species were recorded (e.g., plants for planting, cut branches, palm seeds, nuts, natural spread into the EPPO region), and their relevance noted, but measures were not proposed on pathways other than wood.
2.1.5. Definitions: Non-Coniferous Woody Plants and Their Trade
This study focuses on ‘non-coniferous’ wood, specifically woody angiosperms. An overview of non-coniferous woody plants in the EPPO region and available data on non-coniferous wood trade into the EPPO region were assembled. Although traded wood most commonly originates from ‘trees’, species of bark and ambrosia beetles were considered, irrespective of the type and form of their host plants, as long as those were non-coniferous woody plants. This is because the wood of some other types of non-coniferous plants is also traded (e.g., shrubs such as Buxus), and because woody plants other than trees also need to be considered in relation to the potential risks presented by non-native bark or ambrosia beetles.
Non-coniferous woody plants are key components of many habitat types in the EPPO region, with an overall very high economic, environmental, and social importance. Considering the scale of the EPPO region, it was not realistic to provide a full inventory of non-coniferous trees and bushes in the EPPO region, nor of all habitats concerned. However, major habitats were outlined: forests and other forested environments (including the Mediterranean maquis, riparian environments), plantations of fruit trees and bushes, and urban trees. As previous EPPO PRAs have shown, it was found during this study that most woody hosts are typically available in nurseries in the EPPO region for use as ornamentals, with the exception of exclusively tropical tree species [8]. Ornamental trees may be produced locally or imported, and there is a huge trade of plants for planting into the EPPO region [12].
2.2. Empirical Support: Factorial Discriminant Analyses
The information on risk factors (Table 2) was used to investigate whether they could help predict the amount of damage caused by beetles. For that, a factorial discriminant analysis (FDA, [13]) was used as a supervised classification method to discriminate between three categories of beetle species a priori classified by experts according to their level of damage (‘substantial’, ‘moderate’, and ‘none’; corresponding to the three categories of damage in Section 2.1.2), using risk factors as predictor variables. Results of the FDA were also used to identify which risk factors were the best at discriminating between the different categories of damage.
Table 2.
Rating of risk factors for the 26 representative species and groups of species. Legend: s.l. = sensu lato; Y = Yes; N = No; N? = doubt on the answer; Empty cell: no information available.
The dataset was composed of the 12 risk factors transformed into dummy variables [14] for the 26 beetle species or groups. No assumptions on prior probabilities were required [15]. As preliminary analyses with linear FDA showed that covariance matrices significantly differed among categories (p < 0.0001, Box test), a canonical FDA was performed.
The same FDA was applied for discriminating between two categories of beetle species classified as introduced or not introduced into new areas. The independent variables (predictors) were all risk factors except those related to the level of damage, because damage occurs post–introduction (i.e., inbreeding, polyphagy, ability to attack new hosts, fungus farming, association with pathogenic fungi, maturation feeding, aggregation pheromone, multivoltinism, and association to wood commodities).
All statistical analyses were made with XLSTAT [16].
3. Results
3.1. Representative Bark and Ambrosia Beetle Species
Out of 30 species or groups proposed, 24 species that were considered to present a potential risk were retained as well as one species complex (Euwallacea fornicatus sensu lato) and one group of species (Gnathotrupes spp. of Nothofagus spp.) (Table 2). This selection included 20 ambrosia beetles (12 Scolytinae and eight Platypodinae) and six bark beetles (Scolytinae). Many of the species retained are native or introduced to North America, or introduced into the EPPO region; detailed information is available and, if impact has occurred, extensive research has been conducted and the results are published in the literature.
Several of the selected species (e.g., Xyleborinus artestriatus or X. octiesdentatus) have never caused economic damage but are known to be frequent invaders. These were retained to illustrate pathways or risk factors.
3.2. Risk Factors
Biological and other factors that influence whether a bark or ambrosia beetle presents a potential risk of entry, establishment, spread, and impact for the EPPO region were identified through expert opinion and based on the selected species. Each factor was ranked by expert judgement as of major, medium, or minor importance. Risk factors considered to be of major or medium importance are summarised below, along with illustrative examples. The risk factors were developed in detail in the EPPO study, giving more examples of species. Minor factors were not considered further. For some factors, key elements that are especially important to the risk were identified for some factors and rated against each species. Table 2 presents the ratings of the elements most important to the risk factors for the representative species.
3.2.1. Biological Factors
- Mating strategy (major factor)
Many Scolytinae are inbreeders, which means that they mate with their siblings in the galleries where they developed (sib-mating). This is the case of species in the Xyleborini genera Ambrosiodmus, Cnestus, Euwallacea, Xyleborinus, Xyleborus, and Xylosandrus. Consequently, nearly all females leaving the tree have already mated. Inbreeding therefore facilitates entry (transfer to a host) and establishment.
- Host condition (major factor)
The ability to colonize live trees (whether stressed or apparently healthy) is key to the risk of entry (transfer to a host), establishment, and impact. All bark and ambrosia beetles considered in the EPPO study have some association with live trees. For example, Megaplatypus mutatus attacks only live standing trees, while Austroplatypus incompertus attacks apparently healthy trees and Cnestus mutilatus has been recorded on live stressed trees. Some species attacking live trees have also been recorded attacking timber, which is an advantage for entry (association with wood commodities). This is, for example, the case of Phloeotribus liminaris. For initial colonisation in a new area, some species may be able to utilize various material, from recently dead or weakened to living healthy trees.
- Host specificity (major factor)
Polyphagy and the ability to attack new hosts in new areas are advantages for entry, establishment (higher probability of finding a suitable host), and impact. Polyphagy was considered in terms of the number of host families recorded for the bark and ambrosia beetle species. The most polyphagous species are recorded on plants in over 50–60 families (e.g., Hypothenemus eruditus) (Table 2). Even bark and ambrosia species that are somewhat restricted in their host selection can typically utilize many species within host families or within higher ranks, and such species can colonize confamilial hosts in the newly occupied ranges. Most bark and ambrosia beetles considered in the study have been recorded on new hosts when introduced into new areas (e.g., Xyleborus glabratus).
- Associated fungi (major factor)
Ambrosia beetles are always associated with fungal symbionts [17] and may also carry other fungi. Bark beetles are also associated with fungi, with relationships ranging from casual associations or more consistent but non-obligate associations to coevolved nutritional mutualism [18]. Fungus-farming as carried out by ambrosia beetles is indirectly favourable to establishment as a mechanism for broadening host range. More importantly, an association of a bark or ambrosia beetle with a pathogenic fungus increases the potential impact. Unfortunately, this may not be recognised until the fungus is transported to a new invasion area or a new host, leading to unexpected impact such as took place for Geosmithia morbida (associated with Pityophthorus juglandis).
- Climatic requirements (major factor)
The climatic conditions to which a bark or ambrosia beetle is adapted is important for the potential risk (especially establishment and spread). Species known to occur in areas climatically similar to those occurring in the EPPO region are very likely to establish. Many bark and ambrosia beetles originate from tropical or subtropical areas, and establishment is likely to fail because of an unsuitable climate. Nevertheless, the Mediterranean Basin has already proven to be sufficiently warm for many such species (e.g., Xyleborus bispinatus). In addition, the trend of increasing temperatures in the EPPO region may increase the likelihood of establishment of cold-intolerant species.
- Dispersal capacity (major factor)
The capacity to disperse, either naturally or human-assisted, increases the risk of entry, establishment (finding a host far from the entry point), and spread for all bark and ambrosia beetles. The size of the pioneer population will influence dispersal capacity (because there will be more individuals dispersing long distances among the members of a larger population). All bark and ambrosia beetles have some natural dispersal capacity through flight in at least one of the sexes, but the flight biology and capacity remain essentially unknown for most species. Among the representative species, Platypus quercivorus is recorded as having a good flight capacity (in experiments in a flight mill, some individuals flew over 25 km [19]). Human-assisted dispersal through the movement of wood is, in any case, the main factor for establishing new infestations of bark and ambrosia beetles.
- Mass attacks (medium factor)
Most, if not all, bark and ambrosia beetles infesting non-coniferous wood can establish at low population densities (just a single female in sib-mating species) on recently dead, weakened, or apparently healthy hosts. However, mass-attack, based on aggregation pheromones, can facilitate the establishment of some species (e.g., Acanthotomicus suncei [20]) by allowing them to overcome the defences of living trees. Mass attack as a necessary requirement (which is mostly known for species on coniferous wood) can be unfavourable to establishment, while solitary colonizers have displayed high success in entry and establishment. However, mass attacks once a species is established and the epidemic threshold is reached (e.g., Pityophthorus juglandis or Xylosandrus crassiusculus) can result in higher impact.
- Voltinism and sister broods (medium factor)
The ability to develop several separate or overlapping generations (e.g., Xylosandrus compactus) increases both the risk of spread and impact through higher population density. This also occurs in the case of sister broods (subsequent oviposition by the same female in different galleries).
3.2.2. Additional Factors
- Association with wood commodities (major factor)
All of the representative bark and ambrosia beetles have some association with non-coniferous wood commodities, thus increasing the risk of entry. Bark and ambrosia beetles are commonly associated with wood commodities in trade and are frequently intercepted. This is the case for example for Euplatypus parallelus, Xyleborinus artestriatus, and X. octiesdentatus. The association of bark and ambrosia beetles with different wood commodities is considered in Table 3.
Table 3.
Association of bark and ambrosia beetles with wood commodities.
There are uncertainties about the association of bark and ambrosia beetles with some wood commodities, especially wood chips, hogwood and processing wood residues. However, given the small size of bark and ambrosia beetles (adult body size ranges from <1 mm to 9 mm for the representative species), such an association remains a potentially important risk.
There are also uncertainties about the association with wood commodities for species that preferably attack small diameter material such as branches or twigs, because they would mostly not be present in tree trunks. However, such species may also attack larger diameter material, and some wood commodities may also contain small diameter material or whole trees.
Wood packaging material not treated according to ISPM 15 [11] was considered a potential pathway for all species considered, given the large number of historical interceptions and introductions apparently linked to this pathway. Finally, a potential association with plants for planting (and possibly cut branches) was identified, with variable levels of uncertainty, for all bark and ambrosia beetle species in the EPPO study including those that attack small-diameter material such as twigs.
- Known introductions (major factor)
The past history of introductions of a bark or ambrosia beetle documents a risk of entry and establishment. Most species used as representative examples are known to have moved to new areas including the three Euwallacea spp., Monarthrum mali, and Scolytus schevyrewi. In a review of the literature, a total of 138 exotic Scolytinae species have been recorded to have established in more than one land mass [21].
- Trade of wood commodities (major factor)
The existence of a trade is an essential factor for the risk of entry of bark and ambrosia beetles. At the scale of the EPPO region, there is no complete picture of all wood commodities or genera and species of wood imported into the region. Nevertheless, a huge diversity of non-coniferous species is known to be used for their wood. Data available in FAOSTAT [22] for EPPO countries show a broad trade of non-coniferous wood commodities including tropical and non-tropical round wood, sawn wood, and wood chips and particles. The wood commodities imported from outside the EPPO region come from a large diversity of origins in the northern and southern hemispheres (Table 4).
Table 4.
Summary of analysis of FAOSTAT [22] data. Imports from non-EPPO countries into EPPO countries in 2015.
- Detection and identification in commodities and in trees (major factor)
Bark and ambrosia beetles are difficult to detect and identify, thus increasing the risk of entry and establishment. They are generally small and cryptic, and may be transported in small numbers, making them difficult to detect. Detection may be facilitated where there are known specific attractants (e.g., pheromones or kairomones). Commonly used attractants (e.g., ethanol) allow for a large range of bark and ambrosia beetles to be detected. However, the identification of bark and ambrosia beetles is difficult and requires either specialized taxonomic expertise or access to gene sequencing, which is not always available throughout the EPPO region.
- Difficulty of eradication and containment (major factor)
Eradication and containment of bark and ambrosia beetles is very difficult in nearly all circumstances, increasing the risk of spread and impact. Most non-native species of bark and ambrosia beetles are detected long after establishment, when the population density has increased, and the species may already have started spreading naturally or through human-assisted pathways. In addition, there are few control measures that are effective due to the beetles’ cryptic life cycle.
- Suitable habitats (colonisation of new habitats and natural range of plant species) (major factor)
The existence of suitable habitats increases the risk of entry, establishment, spread, and impact. Areas where the diversity of trees is high are more favourable for entry and establishment because non-indigenous beetles are more likely to find a suitable host. Species may also shift habitats, for example, between forest and orchards (e.g., Euwallacea fornicatus sensu lato, and Ambrosiodmus rubricollis). Finally, urban environments provide high host diversity and trees that are often in stressed condition, enabling colonisation by opportunistic wood borers.
- Known damage (major factor)
Bark and ambrosia beetles may cause economic damage including the degradation of wood value, negative impacts on exports of wood commodities, tree decline or tree death (e.g., Austroplatypus and Platypus species); environmental damage such as ecosystem changes through the death of trees (e.g., Xyleborus glabratus) or social damage including job losses (e.g., due to processing units closing because of losses of wood production) and effects on the recreational value of an area (e.g., Pityophthorus juglandis). Previous high impact is an indicator of potential risk. However, there are often uncertainties in terms of the hosts that will be attacked in a new invaded area, and their susceptibility and the potential damage cannot be accurately predicted. This is especially the case for beetle species for which data is lacking such as Acanthotomicus suncei and Gnathotrupes spp. of Nothofagus.
- Management practices (medium factor)
Good management practices can reduce susceptibility to infestation by bark and ambrosia beetles and would protect plants from attacks by secondary pests. Conversely, management leading to increased tree stress reduces resistance to attack, and/or resilience following an attack.
3.3. Categories of Pests
Based on the information gathered for each species and on expert judgement, the bark and ambrosia beetles were classified into the three categories outlined in Section 2.1.2: (1) Known substantial damage; (2) Indications of damage, with some uncertainties; and (3) No documented damage, but several potential risk factors exist. The category ratings are indicated in Table 2.
3.4. Identification of Horizontal Measures
Based on the information assembled and risk factors identified, the EWG recommended that in order to prevent the introduction of new bark and ambrosia beetles on non-coniferous wood commodities into the EPPO region, horizontal measures should be adopted irrespective of the host plant species and the origin (i.e., for all genera of non-coniferous woody plants and from all origins). In particular, because bark and ambrosia beetles from tropical origins have entered and established in temperate regions, tropical wood would also be covered. Furthermore, treatments that target both bark and ambrosia beetles were recommended.
The study recognized that there are few measures in place for non-coniferous wood commodities in EPPO countries, and requiring such measures is very likely to have an impact on trade, particularly for round wood.
The minimum requirement suggested is that non-coniferous wood commodities should be accompanied by a phytosanitary certificate. Exporting countries would consequently need to perform inspections prior to export.
In addition, pest risk management options were identified and proposed. We considered measures that are already recommended and in use for such commodities, rather than identifying novel management options. Measures were similar for the three commodity groups considered: wood (round wood and sawn wood, with or without bark); wood chips, hogwood, and processing wood residues (except sawdust and shavings); and bark. The horizontal measures proposed were heat treatment, ionizing radiation (only for round wood and sawn wood), and appropriate fumigation. These measures were then approved by EPPO governing bodies, and subsequently recommended to the National Plant Protection Organizations (NPPOs) of EPPO countries. The horizontal measures proposed are already recommended within EPPO for coniferous wood commodities, are effective against bark and ambrosia beetles, and are included in ISPM 39 [9].
Other possible measures were discussed but not proposed, including debarking on its own (because it does not address ambrosia beetles present in the sapwood), chipping to a certain size (because it does not guarantee freedom from the pest), and a generic requirement for a pest free area (PFA) for bark and ambrosia beetles. The inclusion of a PFA in horizontal measures was not considered appropriate, however, as by definition a PFA is related to specific pests, and its suitability and reliability as a measure has to be considered on a case by case basis depending on the biology of the species, the situation in the area(s) where it is present, and the possibility of early detection. Horizontal measures proposed in this study aim to provide protection against a wide range of non-coniferous bark and ambrosia beetles, rather than a series of individual species.
3.5. Factorial Discriminant Analyses for the Categorisation of Beetles According to the Level of Damage and Ability to Colonise New Areas
The first factorial discriminant analysis showed significant differences between the means of the three damage categories of beetles on the combination of risk factors (Wilks’ lambda test, p < 0.0001). It yielded two significant and marginally significant (alpha level 0.05) canonical functions (F1, p < 0.0001 and F2, p = 0.067, respectively; represented by the axes in Figure 1). The first canonical function was mainly driven by three risk factors (i.e., the ability to kill trees, association with pathogenic fungi, and known ability to attack new host tree species (positive values of axis F1)). The second canonical function mainly accounted for the release of an aggregation pheromone (negative values of axis F2). The most damaging beetle species were those that carried pathogenic fungi, produced aggregation pheromones, and were able to colonise and eventually kill new host tree species (Figure 1). The confusion matrix (Table 5) showed 15.4% misclassification, which was only due to four species classified a priori as moderately damaging (category ‘indications of damage, some uncertainties’) and a posteriori classified as substantially damaging, namely Euwallacea interjectus, E. validus, Xyleborus bispinatus, and Xylosandrus crassiusculus.
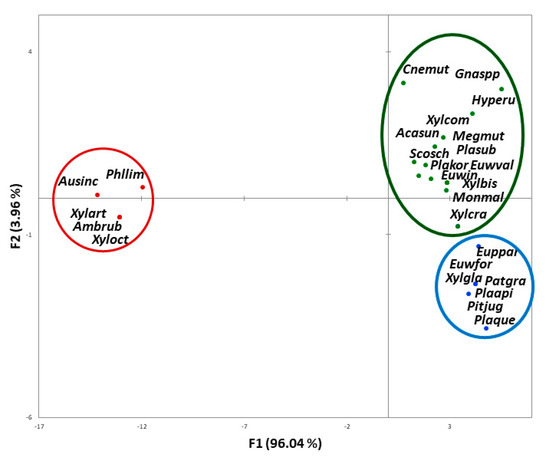
Figure 1.
Scatter plot of the two discriminant functions (with percentage of variance explained in parentheses) showing the position of the 26 species and groups of species, and associated categories of damage [red circle far left (no damage), green oval top right (moderate damage), blue circle bottom right (substantial damage)] in the risk factors space. Acasun: Acanthotomicus suncei; Ambrub: Ambrosiodmus rubricollis; Ausinc: Austroplatypus incompertus; Cnemut: Cnestus mutilatus; Euppar: Euplatypus parallelus; Euwfor: Euwallacea fornicatus; Euwin: Euwallacea. interjectus; Euwval: Euwallacea validus; Gnaspp: Gnathotrupes spp. of Nothofagus; Hyperu: Hypothenemus eruditus; Megmut: Megaplatypus mutatus; Monmal: Monarthrum mali; Patgra: Platypus gracilis; Phllim: Phloeotribus liminaris; Pitjug: Pityophthorus juglandis; Plaapi: Platypus apicalis; Plakor: Platypus koryoensis; Plaque: Platypus quercivorus; Plasub: Platypus subgranosus; Scosch: Scolytus schevyrewi; Xylart: Xyleborinus artestriatus; Xylbis: Xyleborus bispinatus; Xylcom: Xylosandrus compactus; Xylcra: Xylosandrus crassiusculus; Xylgla: Xyleborus glabratus; Xyloct: Xyleborinus octiesdentatus.
Table 5.
Confusion matrix for factorial discriminant analysis of three categories of damage by the bark and ambrosia beetles studied.
The second factorial discriminant analysis showed significant differences between the means of risk factors for introduced vs. not yet introduced beetles (Wilks’ lambda test, p < 0.001), identifying one significant canonical function (F1, p < 0.001). The area under the ROC curve (receiver operating characteristic curve) was 0.99, indicating a very good fit. Species that have already been introduced into new areas were mainly categorised by inbreeding reproduction, polyphagy, and the lack of aggregation pheromone. The confusion matrix (Table 6) showed 3.8% misclassification, which was due to only one species classified a priori as already introduced and a posteriori classified as not introduced, namely Euplatypus parallelus.
Table 6.
Confusion matrix for factorial discriminant analysis of two categories of bark and ambrosia beetles according to their past introduction into new areas.
4. Discussion
Our study identified and detailed factors known to influence the risk of entry, establishment, spread, and impact for bark and ambrosia beetles. The potential pest risk associated with bark and ambrosia beetles relates to a large number of factors that have been considered in the context of pest risk assessment (i.e., risks of entry, establishment, spread, and impact). The species considered in this study exhibited several of these factors, which can individually or in combination increase the pest risk (Table 2). However, the main factors that drive successful establishment and impact vary from species to species and are not always fully identified.
Although only a small proportion of all Scolytinae and Platypodinae probably have the characteristics for successful establishment and impact in the EPPO region, lack of knowledge regarding individual species and unexpected performance in case of introduction to a new area challenge the current approach of pest-specific risk analysis. Bark and ambrosia beetle species respond differently to new conditions, especially host availability and climate. Unexpected damage may occur only when the species colonise a new host, a new area, or become associated with a new pathogenic fungus. The pathogenicity of the associated fungi may also vary with the hosts they encounter.
This study relied mostly on species for which detailed information was available. However, assessing risk is significantly more complicated and subject to uncertainties when limited information is available, or when introduction has not occurred or has not led (yet) to damage (for example, in the case of Gnathotrupes spp. of Nothofagus, or of Acanthotomicus suncei). There are probably hundreds if not thousands of Scolytinae and Platypodinae species of non-coniferous woody plants worldwide that are not yet present in the EPPO region and belong to this category.
Finally, throughout the EPPO region, there is a huge diversity of non-coniferous woody plants that are important in their native habitats or in cultivation and need to be protected from potentially destructive pests. Climate change may have yet unknown consequences in terms of climatic suitability for the pests, but host tree stress is likely to increase in the region in the future, and stressed hosts are typically more susceptible to bark and ambrosia beetle attack. In particular, woody plants in urban environments are often more stressed and may be more susceptible to attack than in other areas.
We concluded that the overall risk cannot be addressed by targeting only particular hosts that are major species for the EPPO region (e.g., key forestry species) because many bark or ambrosia beetles are polyphagous and thus may be introduced to other novel hosts. Consequently, the current approach of relying upon pest-specific PRAs for regulation is too limited to cover the huge diversity of plant species being imported. We also noted that the eradication and containment of bark and ambrosia beetles following establishment and spread are extremely difficult to achieve.
In this study, we considered that tropical wood also presents a risk, as polyphagous beetle species prone to attacking new hosts may be introduced on tropical wood and then establish on novel host species in the EPPO region. There is a known trade of wood from tropical areas to the EPPO region. Tropical and subtropical bark and ambrosia beetle species are known to have established in other climatic conditions, and establishment of such species in the EPPO region is more likely to occur in the future under a warming climate.
The factorial discriminant analyses identified important predictive factors for the impact of invasive bark and ambrosia beetles. The potential level of damage is predicted to be greater for beetle species that combine three characteristics i.e., carry pathogenic fungi, produce aggregation pheromones, and are able to colonise new host tree species. The capacity to colonize new host trees increases with the degree of polyphagy in forest insects [23], which may result from feeding on fungi rather than tree tissues as in ambrosia beetles. Following successful colonisation by pioneer insects, the ability to emit aggregation pheromones results in the attraction of many congeners. The mass attack eventually overcomes the host’s defences, causing tree death and thus more damage [24].
The second factorial discriminant analysis suggested that the ability to colonize new areas is greater in bark and ambrosia beetle species that are polyphagous, reproduce through inbreeding, and do not use aggregation pheromones. Species with a large diet breadth are more likely to find suitable host trees in the area of introduction and to avoid losing fitness whilst switching to new hosts [23]. Inbreeding reproduction is another feature facilitating the successful establishment of a new population as the pioneer beetles do not need to find congeners to mate and reproduce (low Allee threshold). This effect has been recently confirmed by Lantschner et al. [25], showing that scolytine inbreeders have been able to colonize more regions worldwide than outbreeders. Their typical absence of aggregation pheromones supports this, since it suggests that a small number of individuals can successfully colonize new host trees.
It is interesting to note that aggregation pheromones could play opposite roles for any given species. It appears to be a hindrance for introduction (a need for aggregation would correspond to a higher Allee threshold), but an asset for impact (a mass attack is more likely to overwhelm the defences of a potential host). As more information about the biology of individual exotic beetle species becomes available, it is likely that such an approach using factorial discriminant analysis can be used for a better prediction of risk for individual species based on their life history traits. However, counter examples offered by other studies suggest that one should be wary of generalising. Mech et al. [26] reviewed 58 non-native conifer-specialist insects in North America, belonging to five orders, 20 families, and five feeding guilds, and concluded that insect life history traits comparable to those examined in the present study (reproductive strategy, polyphagy, pest status, dispersal, etc.) had little predictive value regarding the impact of these species in new territories, whilst factors related to the newly colonised areas (shared evolutionary history of the native and novel hosts, shade, drought tolerance of the novel hosts, absence of a coevolved congener on a shared host) were better predictors.
5. Conclusions
Based on the analysis made in the EPPO study, the EWG recommended that EPPO member countries adopt phytosanitary measures for bark and ambrosia beetles for all non-coniferous wood from all origins, and a range of phytosanitary measures was identified. It was recognized that applying these measures to all non-coniferous wood from all origins would have significant impact on trade. However, such measures would reduce the risk of further introductions of bark and ambrosia beetles. The NPPOs of EPPO member countries will now individually decide whether and how to implement the measures recommended by EPPO governing bodies. As is normal in the EPPO system, the suitability of individual phytosanitary measures can be reconsidered when new knowledge becomes available.
Although this study focused on wood commodities, the need for the regulation of plants for planting in relation to Scolytinae and Platypodinae should also be emphasised, since many potential pest species may be able to enter via this pathway.
Author Contributions
Conceptualisation, investigation, methodology: all authors; writing—original draft, F.G., J.-C.G. and H.J.; Writing—review and editing, A.B., A.B.B., A.O., B.H., D.I., F.P. and J.H. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
We are grateful to all experts who have participated in the EPPO study. This includes experts who reviewed pest information sheets, PRA core members who reviewed potential risks for individual species, other experts who answered specific queries and provided information as well as the members of the EPPO Panel on Phytosanitary Measures, EPPO Panel on Quarantine Pests for Forestry, and EPPO Working Party on Phytosanitary Regulations for their contribution through a critical analysis of the outcomes. JH was partly funded by a cooperative agreement with USDA APHIS.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hulcr, J.; Atkinson, T.H.; Cognato, A.I.; Jordal, B.H.; McKenna, D.D. Morphology, taxonomy, and phylogenetics of bark beetles. In Bark Beetles, Biology and Ecology of Native and Invasive Species, 1st ed.; Vega, F.E., Hofstetter, R.W., Eds.; Elsevier; Academic Press: London, UK, 2015; pp. 41–84. [Google Scholar]
- Jordal, B. Molecular phylogeny and biogeography of the weevil subfamily Platypodinae reveals evolutionarily conserved range patterns. Mol. Phylogenet. Evol. 2015, 92, 294–307. [Google Scholar] [CrossRef] [PubMed]
- FAO. ISPM 5 Glossary of Phytosanitary Terms; International Plant Protection Convention (IPPC), Food and Agriculture Organization (FAO): Rome, Italy, 2018. [Google Scholar]
- Kirkendall, L.; Faccoli, M. Bark beetles and pinhole borers (Curculionidae, Scolytinae, Platypodinae) alien to Europe. ZooKeys 2010, 56, 227–251. [Google Scholar] [CrossRef] [PubMed]
- Rassati, D.; Lieutier, F.; Faccoli, M. Alien wood-boring beetles in Mediterranean regions. In Insects and Diseases of Mediterranean Forest Systems; Paine, T.D., Lieutier, F., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 293–327. [Google Scholar]
- Inward, D.J.G. Three new species of ambrosia beetles established in the UK illustrate unresolved risks of timber importation. J. Pest. Sci. 2020, 93, 117–126. [Google Scholar] [CrossRef]
- Hulcr, J.; Black, A.; Prior, K.; Chen, C.Y.; Li, H.F. Studies of ambrosia beetles (Coleoptera: Curculionidae) in their native ranges help predict invasion impact. Fla. Entomol. 2017, 100, 257–262. [Google Scholar] [CrossRef]
- EPPO. EPPO Study on the Risk of Bark and Ambrosia Beetles Associated with Imported Non-Coniferous Wood; EPPO Technical Document No. 1081; EPPO: Paris, France, 2020. Available online: https://www.eppo.int/RESOURCES/eppo_publications (accessed on 12 March 2020).
- FAO. ISPM 39 International Movement of Wood; IPPC, FAO: Rome, Italy, 2017. [Google Scholar]
- EPPO. EPPO Study on Wood Commodities Other than Round Wood, Sawn Wood and Manufactured Items; EPPO Technical Document No. 1071; EPPO: Paris, France, 2015.
- FAO. ISPM 15 Regulation of Wood Packaging Material in International Trade; IPPC, FAO: Rome, Italy, 2019. [Google Scholar]
- EPPO. EPPO Study on the Risk of Imports of Plants for Planting; EPPO Technical Document No. 1061; EPPO: Paris, France, 2012.
- Zuur, A.; Ieno, E.N.; Smith, G.M. Analyzing Ecological Data; Springer: New York, NY, USA, 2007; 672p. [Google Scholar]
- Huberty, C.J.; Wisenbaker, J.M.; Smith, J.D.; Smith, J.C. 1986 Using categorical variables in discriminant analysis. Multivar. Behav. Res. 1986, 21, 479–496. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.A. The use of multiple measurements in taxonomic problems. Ann. Eugen. 1936, 7, 179–188. [Google Scholar] [CrossRef]
- Addinsoft. XLSTAT Statistical and Data Analysis Solution; Addinsoft: Long Island, NY, USA, 2019. [Google Scholar]
- Hulcr, J.; Stelinski, L.L. The ambrosia symbiosis: From evolutionary ecology to practical management. Annu. Rev. Entomol. 2017, 62, 285–303. [Google Scholar] [CrossRef] [PubMed]
- Ploetz, R.C.; Hulcr, J.; Wingfield, M.J.; de Beer, Z.W. Destructive tree diseases associated with ambrosia and bark beetles: Black swan events in tree pathology? Plant Dis. 2013, 97, 856–872. [Google Scholar] [CrossRef] [PubMed]
- Okada, R.; Pham, D.L.; Ito, Y.; Yamasaki, M.; Ikeno, H. Measuring the flight ability of the ambrosia beetle, Platypus quercivorus (Murayama), using a low-cost, small, and easily constructed flight mill. J. Vis. Exp. 2018, 138, e57468. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Cognato, A.I. Acanthotomicus suncei, a new sweetgum tree pest in China (Coleoptera: Curculionidae: Scolytinae: Ipini). Zootaxa 2018, 4471, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Grégoire, J.C.; Battisti, A.; Benko Beloglavec, A.; Grousset, F.; Hrašovec, B.; Hulcr, J.; Inward, D.; Jactel, H.; Orlinski, A.; Petter, F. Common Features Among Cosmopolitan Bark- and Ambrosia Beetles: Strong Trends but Too Many Idiosyncrasies for Safe Risk Assessments. Unpublished work, 2020. [Google Scholar]
- FAOSTAT. Statistical data of the Food and Agriculture Organization; FAO: Rome, Italy, 2018; Available online: http://www.fao.org/faostat/en/#home (accessed on 12 March 2020).
- Bertheau, C.; Brockerhoff, E.G.; Roux-Morabito, G.; Lieutier, F.; Jactel, H. Novel insect-tree associations resulting from accidental and intentional biological ‘invasions’: A meta-analysis of effects on insect fitness. Ecol. Lett. 2010, 13, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Raffa, K.F.; Grégoire, J.C.; Lindgren, B.S. Natural history and ecology of bark beetles. In Bark Beetles, Biology and Ecology of Native and Invasive Species, 1st ed.; Vega, F.E., Hofstetter, R.W., Eds.; Elsevier Academic Press: London, UK, 2015; pp. 1–40. [Google Scholar]
- Lantschner, M.V.; Corley, J.C.; Liebhold, A.M. Drivers of global Scolytinae invasion patterns. Ecol. Appl. 2020. [Google Scholar] [CrossRef] [PubMed]
- Mech, A.M.; Thomas, K.A.; Marsico, T.D.; Herms, D.A.; Allen, C.R.; Ayres, M.P.; Gandhi, K.J.K.; Gurevitch, J.; Havill, N.P.; Hufbauer, R.A.; et al. Evolutionary history predicts high-impact invasions by herbivorous insects. Ecol. Evol. 2019, 9, 12216–12230. [Google Scholar] [CrossRef] [PubMed]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).