Response of Vegetation to Changes in Temperature and Precipitation at a Semi-Arid Area of Northern China Based on Multi-Statistical Methods



Abstract
1. Introduction
2. Study Area and Data Sets
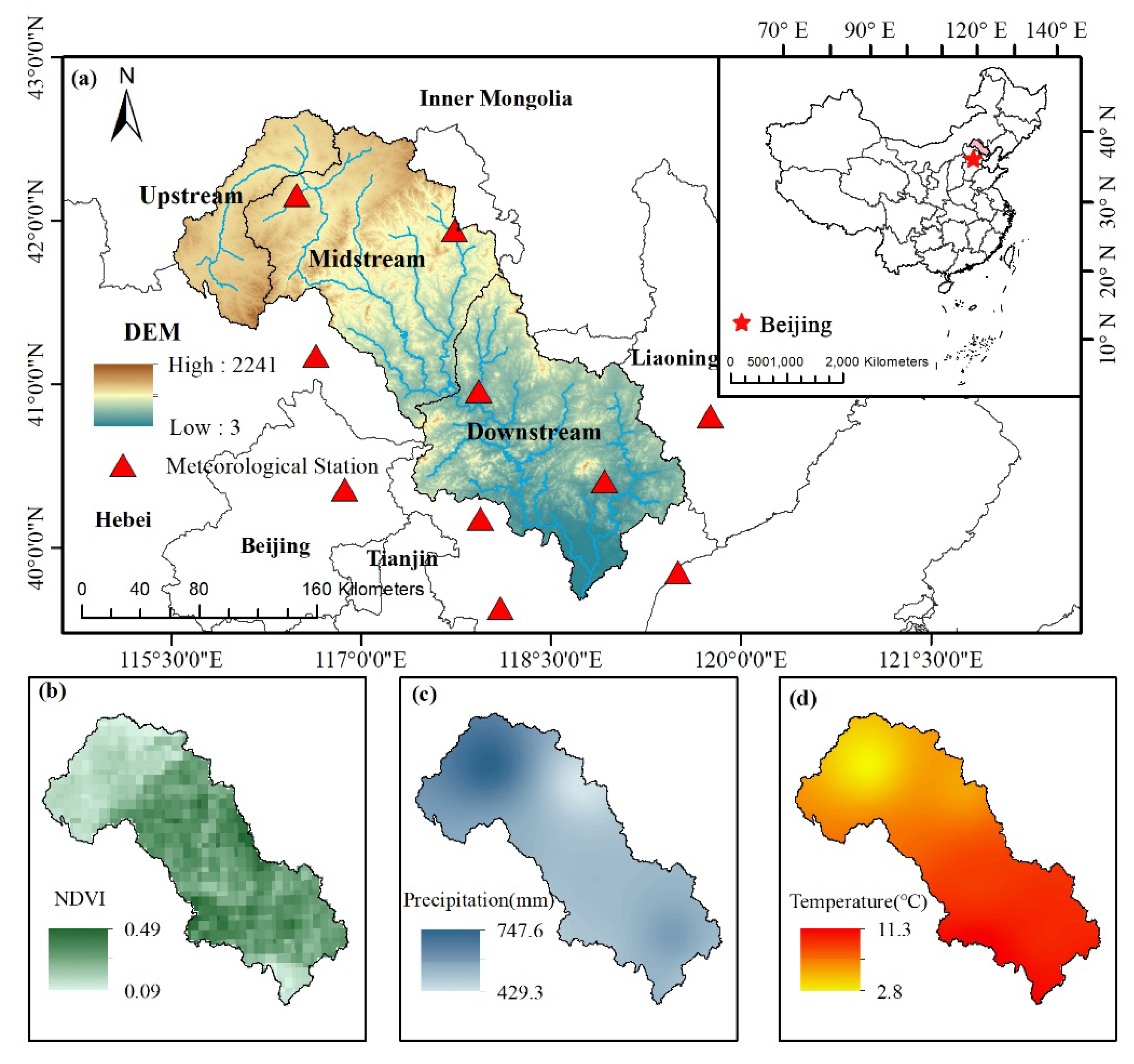
2.1. Study Area
2.2. Data Sets
2.2.1. Data Source
2.2.2. Data Pre-Treatment
3. Methods
3.1. Copula Based Bivariate Correlation Assessment
3.2. Composite Granger Causality Test for Legacy Effect and Casual Relationship
4. Results
4.1. Nonlinear Bivariate Correlation between NDVI and Precipitation/Temperature
4.2. Granger Cause and Legacy Effect
4.2.1. Time Series Sequence Identification
4.2.2. Optimal Lag Length and Granger Causality
5. Discussion
5.1. NDVI Response to Temperature
5.2. NDVI Response to Precipitation
5.3. Implication
5.4. Limitation and Uncertainty
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scheiter, S.; Higgins, S.I. Impacts of climate change on the vegetation of Africa: An adaptive dynamic vegetation modelling approach. Glob. Chang. Biol. 2009, 15, 2224–2246. [Google Scholar] [CrossRef]
- Hou, G.; Zhang, H.; Wang, Y. Vegetation dynamics and its relationship with climatic factors in the Changbai Mountain Natural Reserve. J. Mt. Sci. 2011, 8, 865–875. [Google Scholar] [CrossRef]
- Xu, H.-J.; Wang, X.-P.; Yang, T.-B. Trend shifts in satellite-derived vegetation growth in Central Eurasia, 1982–2013. Sci. Total Environ. 2017, 579, 1658–1674. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.; Zhou, X.; Yin, G.; Hao, F.; Zhang, X.; Hao, Z.; Singh, V.P.; Fu, Y.H. Extended growing season reduced river runoff in Luanhe River basin. J. Hydrol. 2020, 124538. [Google Scholar] [CrossRef]
- Fu, Y.H.; Piao, S.; Zhao, H.; Jeong, S.-J.; Wang, X.; Vitasse, Y.; Ciais, P.; Janssens, I.A. Unexpected role of winter precipitation in determining heat requirement for spring vegetation green-up at northern middle and high latitudes. Glob. Chang. Biol. 2014, 20, 3743–3755. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.; Mohammat, A.; Fang, J.; Cai, Q.; Feng, J. NDVI-based increase in growth of temperate grasslands and its responses to climate changes in China. Glob. Environ. Chang. 2006, 16, 340–348. [Google Scholar] [CrossRef]
- Piao, S.; Wang, X.; Ciais, P.; Zhu, B.; Wang, T.A.O.; Liu, J.I.E. Changes in satellite-derived vegetation growth trend in temperate and boreal Eurasia from 1982 to 2006. Glob. Chang. Biol. 2011, 17, 3228–3239. [Google Scholar] [CrossRef]
- Chu, H.; Venevsky, S.; Wu, C.; Wang, M. NDVI-based vegetation dynamics and its response to climate changes at Amur-Heilongjiang River Basin from 1982 to 2015. Sci. Total Environ. 2019, 650, 2051–2062. [Google Scholar] [CrossRef]
- Lamchin, M.; Lee, W.-K.; Jeon, S.W.; Wang, S.W.; Lim, C.H.; Song, C.; Sung, M. Long-term trend and correlation between vegetation greenness and climate variables in Asia based on satellite data. Sci. Total Environ. 2018, 618, 1089–1095. [Google Scholar] [CrossRef]
- Igbawua, T.; Zhang, J.; Chang, Q.; Yao, F. Vegetation dynamics in relation with climate over Nigeria from 1982 to 2011. Environ. Earth Sci 2016, 75, 596. [Google Scholar] [CrossRef]
- Al-Harthy, M.; Begg, S.; Bratvold, R.B. Copulas: A new technique to model dependence in petroleum decision making. J. Pet. Sci. Eng. 2007, 57, 195–208. [Google Scholar] [CrossRef]
- Kendall, M.G. A new measure of rank correlation. Biometrika 1938, 30, 81–93. [Google Scholar] [CrossRef]
- Sklar, M. Fonctions de repartition a n dimensions et leurs marges. Publ. Inst. Stat. Univ. Paris 1959, 8, 229–231. [Google Scholar]
- Kong, D.; Miao, C.; Duan, Q.; Lei, X.; Li, H. Vegetation-Climate Interactions on the Loess Plateau: A Nonlinear Granger Causality Analysis. J. Geophys. Res. 2018, 123, 11068–11079. [Google Scholar] [CrossRef]
- Granger, C.W.J. Investigating Causal Relations by Econometric Models and Cross-spectral Methods. Econometrica 1969, 37, 424. [Google Scholar] [CrossRef]
- Granger, C.W.J.; Siklos, P.L. Systematic sampling, temporal aggregation, seasonal adjustment, and cointegration theory and evidence. J. Econom. 1995, 66, 357–369. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, Y.; Hao, F.; Li, C.; Wang, X. Hydrological Components Variability under the Impact of Climate Change in a Semi-Arid River Basin. Water 2019, 11, 1122. [Google Scholar] [CrossRef]
- Weiser, C.J. Cold Resistance and Injury in Woody Plants: Knowledge of hardy plant adaptations to freezing stress may help us to reduce winter damage. Science 1970, 169, 1269–1278. [Google Scholar] [CrossRef]
- Nelsen, R.B. An introduction to Copulas, 2nd. ed.; Corr. 2. pr. Softcover version of original hardcover edition 2006; Springer: New York, NY, USA, 2010; ISBN 9780387286785. [Google Scholar]
- Ribeiro, A.F.S.; Russo, A.; Gouveia, C.M.; Páscoa, P. Copula-based agricultural drought risk of rainfed cropping systems. Agric. Water Manag. 2019, 223, 105689. [Google Scholar] [CrossRef]
- Tu, C. Cointegration-based financial networks study in Chinese stock market. Phys. A Stat. Mech. Its Appl. 2014, 402, 245–254. [Google Scholar] [CrossRef]
- Sims, C.A. Macroeconomics and Reality. Econometrica 1980, 48, 1. [Google Scholar] [CrossRef]
- Maziarz, M. A review of the Granger-causality fallacy. J. Philosoph. Econ. Reflect. Econ. Soc. Issues 2015, VIII, 86–105. [Google Scholar]
- Nerlove, M.; Wallis, K.F. Use of the Durbin-Watson Statistic in Inappropriate Situations. Econometrica 1966, 34, 235. [Google Scholar] [CrossRef]
- Lucht, W.; Prentice, I.C.; Myneni, R.B.; Sitch, S.; Friedlingstein, P.; Cramer, W.; Bousquet, P.; Buermann, W.; Smith, B. Climatic control of the high-latitude vegetation greening trend and Pinatubo effect. Science 2002, 296, 1687–1689. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y. Preliminary Estimation of the Realistic Optimum Temperature for Vegetation Growth in China. Environ. Manag. 2013, 52, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Kimball, J.S.; Nemani, R.R.; Running, S.W.; Hong, Y.; Gourley, J.J.; Yu, Z. Vegetation Greening and Climate Change Promote Multidecadal Rises of Global Land Evapotranspiration. Sci. Rep. 2015, 5, 15956. [Google Scholar] [CrossRef]
- Saxe, H.; Cannell, M.G.R.; Johnsen, Ø.; Ryan, M.G.; Vourlitis, G. Tree and forest functioning in response to global warming. New Phytol. 2001, 149, 369–399. [Google Scholar] [CrossRef]
- Qian, Y.Q.; He, F.P.; Wang, W. Seasonality, rather than Nutrient Addition or Vegetation Types, Influenced Short-Term Temperature Sensitivity of Soil Organic Carbon Decomposition. PLoS ONE 2016, 11, e0153415. [Google Scholar] [CrossRef]
- Monteith, J.L. The Photosynthesis and Transpiration of Crops. Exp. Agric. 1966, 2, 1–14. [Google Scholar] [CrossRef]
- Criddle, R.S.; Hopkin, M.S.; Mcarthur, E.D.; Hansen, L.D. Plant distribution and the temperature coefficient of metabolism. Plant. Cell Environ. 1994, 17, 233–243. [Google Scholar] [CrossRef]
- Ong, C.K.; Wilson, J.; Deans, J.D.; Mulayta, J.; Raussen, T.; Wajja-Musukwe, N. Tree–crop interactions: Manipulation of water use and root function. Agric. Water Manag. 2002, 53, 171–186. [Google Scholar] [CrossRef]
- Mitchell, C.P.; Ford-Robertson, J.B.; Hinckley, T.; Sennerby-Forsse, L. Ecophysiology of Short Rotation Forest Crops; Springer: Amsterdam, The Netherlands, 1992; p. 8489. [Google Scholar]
- Ichii, K.; Kawabata, A.; Yamaguchi, Y. Global correlation analysis for NDVI and climatic variables and NDVI trends: 1982–1990. Int. J. Remote Sens. 2002, 23, 3873–3878. [Google Scholar] [CrossRef]
- Ukkola, A.M.; Prentice, I.C.; Keenan, T.F.; Van Dijk, A.I.; Viney, N.R.; Myneni, R.B.; Bi, J. Reduced streamflow in water-stressed climates consistent with CO2 effects on vegetation. nclimate 2016, 6, 75–78. [Google Scholar] [CrossRef]
- Massoud, E.; Turmon, M.; Reager, J.; Hobbs, J.; Liu, Z.; David, C.H. Cascading Dynamics of the Hydrologic Cycle in California Explored through Observations and Model Simulations. Geosciences 2020, 10, 71. [Google Scholar] [CrossRef]
- Bai, Y.; Ochuodho, T.O.; Yang, J. Impact of land use and climate change on water-related ecosystem services in Kentucky, USA. Ecol. Indic. 2019, 102, 51–64. [Google Scholar] [CrossRef]
- Yang, X.J. China’s rapid urbanization. Science 2013, 342, 310. [Google Scholar] [CrossRef] [PubMed]
- Massoud, E.C.; Purdy, A.J.; Miro, M.E.; Famiglietti, J.S. Projecting groundwater storage changes in California’s Central Valley. Sci. Rep. 2018, 8, 12917. [Google Scholar] [CrossRef] [PubMed]
- Gourdji, S.M.; Sibley, A.M.; Lobell, D.B. Global crop exposure to critical high temperatures in the reproductive period: Historical trends and future projections. Environ. Res. Lett. 2013, 8, 24041. [Google Scholar] [CrossRef]
- Victor, D.G.; Ausubel, J.H. Restoring the Forests. Foreign Aff. 2000, 79, 127. [Google Scholar] [CrossRef]
- Zhang, P.; Cai, Y.; Yang, W.; Yi, Y.; Yang, Z.; Fu, Q. Contributions of climatic and anthropogenic drivers to vegetation dynamics indicated by NDVI in a large dam-reservoir-river system. J. Clean. Prod. 2020, 256, 120477. [Google Scholar] [CrossRef]
- Qu, S.; Wang, L.; Lin, A.; Yu, D.; Yuan, M. Distinguishing the impacts of climate change and anthropogenic factors on vegetation dynamics in the Yangtze River Basin, China. Ecol. Indic. 2020, 108, 105724. [Google Scholar] [CrossRef]
- Bakhshi, J.; Javadi, S.A.; Tavili, A.; Arzani, H. Study on the effects of different levels of grazing and exclosure on vegetation and soil properties in semi-arid rangelands of Iran. Acta Ecol. Sin. 2019. [Google Scholar] [CrossRef]
- Zhao, P.; Fan, W.; Liu, Y.; Xu, X. Calculation of FAPAR over ragged terrains: A case study at Saihanba. In Proceedings of the 2016 IEEE International Geoscience & Remote Sensing Symposium, Beijing, China, 10–15 July 2016; pp. 3656–3659. [Google Scholar]
- Loehle, C. Height growth rate tradeoffs determine northern and southern range limits for trees. J. Biogeogr. 1998, 25, 735–742. [Google Scholar] [CrossRef]
- Steiner, K.; Herweg, K.; Dumanski, J. Practical and cost-effective indicators and procedures for monitoring the impacts of rural development projects on land quality and sustainable land management. Agric. Ecosyst. Environ. 2000, 81, 147–154. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | C (u, v) | Parameter |
|---|---|---|
| Clayton | ||
| Frank | ||
| Gumbel |
| Season | Sub-Watersheds | Series | Frank | Clayton | Gumbel | |||
|---|---|---|---|---|---|---|---|---|
| RMSE | AIC | RMSE | AIC | RMSE | AIC | |||
| Late Spring | Upstream | NDVI-P | 0.067 | −4.403 | 0.068 | −4.365 | 0.074 | −4.219 |
| NDVI-T | 0.024 | −6.460 | 0.027 | −6.253 | 0.056 | −4.767 | ||
| Midstream | NDVI-P | 0.108 | −3.457 | 0.105 | −3.498 | 0.119 | −3.258 | |
| NDVI-T | 0.046 | −5.168 | 0.045 | −5.213 | 0.095 | −3.705 | ||
| Downstream | NDVI-P | 0.138 | −2.966 | 0.133 | −3.032 | 0.153 | −2.751 | |
| NDVI-T | 0.059 | −4.778 | 0.061 | −4.604 | 0.118 | −3.268 | ||
| Summer | Upstream | NDVI-P | 0.060 | −4.633 | 0.063 | −4.530 | 0.060 | −4.629 |
| NDVI-T | 0.055 | −4.787 | 0.057 | −4.733 | 0.060 | −4.636 | ||
| Midstream | NDVI-P | 0.039 | −5.493 | 0.040 | −5.449 | 0.040 | −5.424 | |
| NDVI-T | 0.039 | −5.502 | 0.043 | −5.279 | 0.044 | −5.246 | ||
| Downstream | NDVI-P | 0.047 | −5.108 | 0.048 | −5.072 | 0.048 | −5.086 | |
| NDVI-T | 0.049 | −5.027 | 0.051 | −4.952 | 0.052 | −4.901 | ||
| Early Autumn | Upstream | NDVI-P | 0.087 | −3.797 | 0.086 | −3.885 | 0.091 | −3.792 |
| NDVI-T | 0.039 | −5.465 | 0.039 | −5.477 | 0.079 | −4.068 | ||
| Midstream | NDVI-P | 0.108 | −3.454 | 0.111 | −3.393 | 0.115 | −3.329 | |
| NDVI-T | 0.051 | −4.966 | 0.057 | −4.712 | 0.106 | −3.496 | ||
| Downstream | NDVI-P | 0.095 | −3.698 | 0.101 | −3.575 | 0.106 | −3.488 | |
| NDVI-T | 0.059 | −4.667 | 0.064 | −4.496 | 0.097 | −3.662 | ||
| Sub-Watersheds | Factors | ADF Coefficient | t-Statistic | p | Conclusion |
|---|---|---|---|---|---|
| Upstream | NDVI | −3.914 | −3.421 | 0.012 | Stationary |
| T | −3.701 | −3.422 | 0.023 | Stationary | |
| P | −4.31 | −3.421 | 0.003 | Stationary | |
| Midstream | NDVI | −2.727 | −3.421 | 0.066 | Non-stationary |
| T | −3.791 | −3.422 | 0.018 | Stationary | |
| P | −4.434 | −3.421 | 0.002 | Stationary | |
| Downstream | NDVI | −3.309 | −3.421 | 0.046 | Stationary |
| T | −3.774 | −3.421 | 0.018 | Stationary | |
| P | −4.302 | −3.421 | 0.003 | Stationary |
| Sub-Watershed | Optimal Lag Length | H0 (Null Hypothesis) | F-Statistics | p-Value | Conclusion |
|---|---|---|---|---|---|
| Upstream | 2 | NDVI is not the Granger cause of Precipitation | 3.078 | 0.1011 | Accept |
| Precipitation is not the Granger cause of NDVI | 12.6634 | 0.0031 | Reject | ||
| Downstream | 3 | NDVI is not the Granger cause of Precipitation | 1.5953 | 0.2464 | Accept |
| Precipitation is not the Granger cause of NDVI | 5.0371 | 0.028 | Reject | ||
| Upstream | 2 | NDVI is not the Granger cause of Temperature | 0.132 | 0.9383 | Accept |
| Temperature is not the Granger cause of NDVI | 13.3876 | 0.0001 | Reject | ||
| Midstream | 4 | NDVI is not the Granger cause of Temperature | 0.8006 | 0.6574 | Accept |
| Temperature is not the Granger cause of NDVI | 22.8862 | 0.0012 | Reject | ||
| Downstream | 4 | NDVI is not the Granger cause of Temperature | 0.9745 | 0.5788 | Accept |
| Temperature is not the Granger cause of NDVI | 35.8145 | 0.0007 | Reject |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Zhang, X.; Fu, Y.; Hao, F.; Yin, G. Response of Vegetation to Changes in Temperature and Precipitation at a Semi-Arid Area of Northern China Based on Multi-Statistical Methods. Forests 2020, 11, 340. https://doi.org/10.3390/f11030340
Wu Y, Zhang X, Fu Y, Hao F, Yin G. Response of Vegetation to Changes in Temperature and Precipitation at a Semi-Arid Area of Northern China Based on Multi-Statistical Methods. Forests. 2020; 11(3):340. https://doi.org/10.3390/f11030340
Chicago/Turabian StyleWu, Yifan, Xuan Zhang, Yongshuo Fu, Fanghua Hao, and Guodong Yin. 2020. "Response of Vegetation to Changes in Temperature and Precipitation at a Semi-Arid Area of Northern China Based on Multi-Statistical Methods" Forests 11, no. 3: 340. https://doi.org/10.3390/f11030340
APA StyleWu, Y., Zhang, X., Fu, Y., Hao, F., & Yin, G. (2020). Response of Vegetation to Changes in Temperature and Precipitation at a Semi-Arid Area of Northern China Based on Multi-Statistical Methods. Forests, 11(3), 340. https://doi.org/10.3390/f11030340