Abstract
Both exotic and native plant invasions can have profound impacts on ecosystems. While many studies have examined the effects of exotic plant invasions on soil properties, relatively few have tested the effects of native plant invasions on soil microbial communities. Furthermore, we know little about the effects of native plant invasions on microbial communities in litter. In subtropical forests in southern China, we sampled litter at three decomposition stages and top soil in three forest sands representing three stages of the invasion (not invaded, moderately and heavily invaded) by the Moso bamboo (Phyllostachys edulis (Carriere) J. Houzeau), a native species in China. We measured chemical properties (concentrations of C, N, P, Mg, Al, K, Ca, Mn, Cu, and Zn, and concentrations of cellulose and lignin) and microbial communities in litter and/or soil. The bamboo invasion, in general, decreased the element concentrations in litter and soil and also decreased total microbial abundance and diversity. Considering bacteria and fungi separately, the bamboo invasion decreased fungal diversity in litter and soil, but had little impact on bacterial diversity, suggesting that fungi are more sensitive and vulnerable to the bamboo invasion than bacteria. We conclude that native Moso bamboo invasions into subtropical forests may lead to a complex biogeochemical process in the litter–soil system, which may threaten local forest ecosystems by affecting microbial communities and, thus, litter decomposition and nutrient cycling.
1. Introduction
Invasions of exotic plant species, from their native ranges into new regions or countries where they did not occur historically, havebecome common worldwide and are receiving increasing attention [1,2,3,4,5]. It is well known thatexotic plant invasions can decrease native species diversity [6,7], changespecies composition [8,9], alterphysio-chemical properties in soil [10,11,12] and litter [13,14,15], and modify soil microbial communities [16,17,18]. Similarly, native plant invasions, i.e., invasions of native species into a new ecosystem located in the same region or country, may also cause significant ecological impacts on physio-chemical properties in soil [19,20,21] and litter [22,23] and soil microbial communities [24,25,26]. While the effects of exotic plant invasions have received much attention, relatively few studies have considered the impacts of native plant invasions [27,28,29], in particular on microbial communities in litter. Understanding the impacts on litter microbial communities is important as the quantity, diversity, and activity of soil microbes in litter play key roles in litter decomposition and, thus, nutrient cycling [13,14,15].
The invasion of native Moso bamboos (Phyllostachys edulis (Carriere) J. Houzeau) into subtropical evergreen forests has become a big problem in many areas in southern China [30]. In the past, the management of pure Moso bamboo forests for harvesting bamboo shoots and wood was profitable. Therefore, a large area of subtropical evergreen forests in southern China was turned into Moso bamboo forests by farmers [31]. However, with increasing labor costs and decreasing use of bamboo products, many Moso bamboo forests have been abandoned because they are no longer profitable [32]. After thiscease inmanagement, Moso bamboos started to invade adjacent evergreen forests, gradually replacing local species [33]. Previous studies have shown that Moso bamboo invasions could affect the diversity and composition of local plant species [34] and alter concentrationsof carbon (C), nitrogen (N), phosphorus (P), and organic matter in the soil [35,36,37,38]. However, little is known regarding the effect of Moso bamboo invasions on microbial communities in litter.
Plant invasions can greatly alter the structure and function of microbial communities [25,39,40]. For instance, both exotic and native plant invasions were found to increase bacterial and/or fungal diversity in soil and change soil microbial competition [17,18,24,41]. Exotic plant invasions could increase the abundance of nitrogen-fixing bacteria in the rhizosphere, resulting in increased nitrogen availability for plant growth [18]. Furthermore, exotic plant invasions canalter the diversity and composition of microbial communities in litter [42,43,44,45]. There is also evidence that exotic or native plant invasions haveno impact on litter microbial communitiesin the short term [46]. Thus, more studies are needed to test the generality of plant invasion impacts on microbial communities, especially those of native plant invasions, on the diversity and composition of litter microbial communities.
We sampled litter and soil in a secondary broadleaved forest (representing the forest stage not invaded by Moso bamboos), a mixed forest of broadleaved trees and bamboos (representing the forest stage moderately invaded by Moso bamboos), and a Moso bamboo forest (representing the forest stage heavily invaded by Moso bamboos). We measured their chemical properties and microbial communities. Specifically, we address the following questions: (1) Does the Moso bamboo invasion alter chemical properties in soil and litter? (2) Does the Moso bamboo invasion alter diversity and composition of microbial communities in soil and litter? We expected that the bamboo invasion would decrease nutrient element concentrations in soil and litter and thus lead to changes in microbial diversity and composition in soil and litter.
2. Materials and Methods
2.1. Study Site
The study was carried out at Miaoshanwu Nature Reserve (119°56′~120°02′ E; 30°03′~30°06′ N) in Fuyang, Zhejiang Province, China. The area has a subtropical monsoon climate, with a mean annual temperature of 16.1°C and an annual precipitation of 1000 to 1964 mm. The mean annual sunshine duration is 1995 h, and the frost-free period is approximately 232 days per year [47]. The zonal vegetation is subtropical evergreen broadleaved forests, and dominant tree species include Cyclobalanopsis glauca (Thunb.) Oerst., Lithocarpusglaber (Thunb.) Nakai., Quercus fabri Hance., Liquidambar formosana Hance., Schimasuperba Gardn. et Champ., Phoebe chekiangensis C. B. Shang., Cinnamomum camphora (L.) Sieb., and Machilus leptophylla Hand. -Mazz.
2.2. Sampling Design
In August 2018, we selected three transects spanning from a secondary broadleaved evergreen forest, via a transition zone, to a Moso bamboo forest, representing three typical stages of the Moso bamboo invasion into subtropical forests. The secondary broadleaved forest (SF, representing the forest stage not invaded by Moso bamboos) included mainly Phoebe chekiangensis C. B. Shang, Machilus leptophylla Hand.-Mazz., and a few herbs such as Dryopteris sparsa (Buch.-Ham. ex D. Don) O. Ktze. In the forest at the transition zone (TF, representing the forest stage moderately invaded by Moso bamboos), 60% of the area is covered by the bamboos and 40% by broadleaved evergreen trees. In the Moso bamboo forest (MB, representing the forest stage heavily invaded by Moso bamboos), the bamboos are the only species at both tree and shrub layers, and the common herbs include D.sparsa.
We established a 10 × 10 m plot in each of the three invasion stages (i.e., SF, TF, and MB) along each of the three transects. There were three replicates per invasion stage and nine plots in total. Within each of the nine plots, we randomly selected five subplots of 1 × 1 m. In each subplot, we separately collected three litter samples at three different decomposition stages (coded as Stage I, II and III) and a soil sample at 0–10 cm depth (coded as IV for simplicity). In Stage I, the litter sample had not been decomposed; in Stage II, the litter sample had been partially decomposed; and in Stage III, the litter sample had been completely decomposed [48]. Five samples at the same stage and collected in the five subplots of the same plot were pooled into a composite sample.
After collection in the field, the soil samples were sieved to pass a 2-mm mesh, and the litter samples were cut into 0.25 cm2 pieces after removing small particles. One part of each sample was kept in an icebox, transported to the laboratory, and then stored at −80°C for microbial analyses. The other part of each soil sample was air-dried, sieved to pass a 0.15-mm mesh, and then finely ground for measuring concentrations of total organic carbon (TOC), total nitrogen (TN), and total phosphorus (TP). The remained part of each litter sample was dried at 65°C for 72 h, sieved to pass a 0.425-mm mesh, and then finely ground for measuring concentrations of cellulose, lignin, total carbon (TC), TN, TP, Al, Ca, Cu, Fe, K, Mg, Mn, and Zn.
2.3. Chemical Analyses
To compare the carbon content in soil and litter, we measured soil TOC, but not soil inorganic carbon. The concentrations of TOC in soil, TC in litter, and TN in both soil and litter were measured by a Multi N/C 3000 analyzer (Analytik Jena AG, Jena, Germany) [49]. The concentrations of TP in both soil and litter were measured by the Mo-Sb antispetrophotography method after the samples were digested with nitric acid.The concentrations of K in litter samples were measured by flamephotometry. The concentrations of Al, Ca, Cu, Fe, Mg, Mn, and Zn in litter samples were determined by inductively coupled plasma–optical emission spectroscopy (ICP–OES, iCAP 7400, Thermo Fisher Scientific, Waltham, MA, USA) after complete digestion.
To measure the concentrations of lignin and cellulose, litter samples were firstly hydrolyzed by the sulphuric acid. Next, the acid-soluble cellulose and lignin were measured by anthrone colorimetry and ultraviolet spectrophotometer, respectively. Then the acid-insoluble lignin after acid hydrolysis was determined as the mass of insoluble residue [50].
2.4. Microbial Analysis
Total genomic DNA was extracted from 500 mg of each soil and litter sample using the E.Z.N.A.® Stool DNA Kit (D4015, Omega, Inc., Norcross, GA, USA). The V3–V4 region of the prokaryotic (bacterial and archaeal) small-subunit (16S) rRNA gene was amplified with slightly modified versions of the primers of 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′). The ITS2 region of the eukaryotic (fungi) small-subunit rRNA gene was amplified with slightly modified versions of the primers of fITS7 (5′-GTGARTCAT CGAATCTTTG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′). The 5′ ends of the primers were sequencing universal primers tagged with specific barcodes.
To amplify the prokaryotic 16S and ITS fragments, the PCR conditions included an initial denaturation at 98 ℃for 30 s, 35 cycles of denaturation at 98 ℃ for 10 s, annealing at 54 ℃/52 ℃ for 30 s, and extension at 72 ℃ for 45 s, and then final extension at 72 ℃ for 10 min. PCR amplification was performed in a 25-µL reaction mixture consisting of 25 ng template DNA, 12.5 µL PCR premix, 2.5 µL of each primer, and PCR-grade water to adjust the volume. The PCR crude products were purified and quantified. The amplicon pools were prepared for sequencing. The libraries were sequenced on 300PE MiSeq runs, and one library was sequenced with both protocols using the standard Illumina sequencing primers to eliminate the need for a third (or fourth) index read.
2.5. Bioinformatics and Statistical Analysis
Microbial samples were sequenced on an Illumina MiSeq platform according to the manufacturer’s recommendations, provided by LC-Bio. Paired-end reads were assigned to samples based on their unique barcode and truncated by cutting off the barcode and primer sequence. They were merged using FLASH (Fast length adjustment of short reads). Quality filtering on the raw tags was performed under specific filtering conditions to obtain high-quality clean tags according to the fqtrim (v0.94). Chimeric sequences were filtered using Vsearch software (v2.3.4). Sequences with ≥97% similarity were assigned to the same operational taxonomic units (OTUs) by Vsearch (v2.3.4) [51,52,53].
For bacteria, a total of 1, 167, 540 and 396, 976 valid sequencing reads were obtained from the 27 litter samples and the ninesoil samples, respectively. The sequences were clustered into 1,020,078 and 347,410 reads for litter and soil, respectively. Based on the sequences, 7363 and 4498 OTUs were obtained for the litter and soil samples, respectively, and were used for downstream analysis after filtration. For fungi, a total of 812,759 and 168,759 valid sequencing reads were obtained from the 27 litter and 9 soil samples, respectively. The sequences were clustered into 787,458 and 164,611 reads for the litter and soil samples, respectively. A total of 2623 and 987 OTUs for the litter and soilsamples, respectively, were used for downstream analysis.
Representative sequences were chosen for each OTU, and taxonomic data were then assigned to each representative sequence using the Ribosomal Database Project classifier. The differences of the dominant species between different groups and multiple sequence alignments were conducted using the mafft software (v7.310) to study the phylogenetic relationship between different OTUs. OTU abundance information was normalized using the standard of sequence number corresponding to the sample with the least sequences. Alpha diversity was applied in analyzing the complexity of species diversity for a sample through four indices, including the Chao1, Shannon, Simpson, and the observed species indices, which were calculated with QIIME (v1.8.0).
We used two-way ANOVA to examine effects of forest types (no invasion (SF), intermediate invasion (TF) and heavy invasion (MB)) and litter decomposition stages (I, II, III, and/or VI) on the concentrations of macro-elements (C, N, P, and K) and meso- and micro-elements (Al, Ca, Cu, Fe, K, Mg, Mn, and Zn), the concentrations of cellulose and lignin, and microbial diversity (observed OTUs, Chao1 and Shannon indices). When a significant effect was detected, Duncan’s tests were used for multiple comparisons. Before ANOVA, data were checked for homogeneity of variance (by the Bartlett test). Data on concentrations of C, Mn, and Zn and Fe were logarithmically transformed, and data on the concentration of Mg were reciprocally transformed. SPSS (v19.0; IBM, Armonk, NY, USA) was used for ANOVA.
To examine whether microbial communities of the three forest types (SF, TF, and MB) and/or the four litter decomposition stages (I, II, III, and IV) could be distinguished, we analyzed microbial community data (OTUs with relative abundances >1% and occurringin >2 samples on the genus level) by principal coordinate analysis (PCoA). This analysis was done for the bacterial and fungus communities separately. The Bray–Curtis distance was used as a dissimilarity measure in PCoA. The analyses were conducted using CANOCOv5 (Biometrics Plant Research International, Wageningen, Netherlands) [54,55,56].
3. Results
3.1. Macro-Elements, Cellulose, and Lignin
The concentrations of C, N, P, K, and lignin varied significantly among the three forest types (Figure 1), showing an order of SF > TF > MB for the C, N, P and lignin concentrations, and an order of TF > SF > MB for the K concentration (Figure 1). The concentrations of C, N, P, K, cellulose, and lignin decreased with the decomposition stage from I to IV (all p < 0.01) in the three forest types, except for the P and K concentrations in MB (Figure 1). Significant interactions between forest types and decomposition stages were detected for the concentrations of C, N, P, and K, but not for those of cellulose and lignin, indicating that variations of the C, N, P, and K concentrations among the decomposition stages depended on forest types (Figure 1). Forest types had no effect on the cellulose concentration (p > 0.05; Figure 1g).
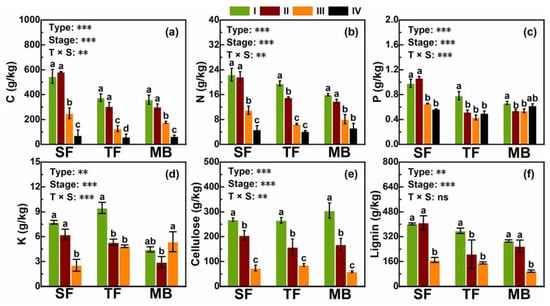
Figure 1.
Concentrations of macro-elements, cellulose, and lignin at the four stages of litter decomposition (I, II, III and IV) in the three forest types. (a–f) a–e, in turn, is the concentration of C, N, P, K, Celllose, and Lignin in the litter and soil. SF—secondary broadleaved forest, representing the stage not invaded by Moso bamboos; TF—forest at the transition zone, representing the stage moderately invaded by Moso bamboos; MB—pure Moso bamboo forest, representing the stage heavily invaded by Moso bamboos. Mean, SE (n = 9) and the significance levels (*** p < 0.001, ** p < 0.01, * p < 0.05, and ns p > 0.05) of two-way ANOVAs are also given. Different letters indicate significant differences between the four stages within a forest type. Concentrations of K, cellulose, and lignin were measured only for litter (I, II, and III), but not for soil (IV).
3.2. Meso- and Micro-Elements
Except for Fe (Figure 2g), the meso- and micro-elements measured significantly varied with both forest type and decomposition stage (Figure 2). However, only Ca and Al were influenced by the interaction between forest type and decomposition stage (Figure 2a–c). Fe in all the three forest types (Figure 2g) and Al in MB and TF (Figure 2b) did not change with the decomposition stage; all other elements in other cases showed a decreasing trend with increasingdecomposition stage from I to IV (Figure 2). Except for Fe (MB = TF = SF), SF had the highest levels of those meso- and micro-elements (Figure 2).
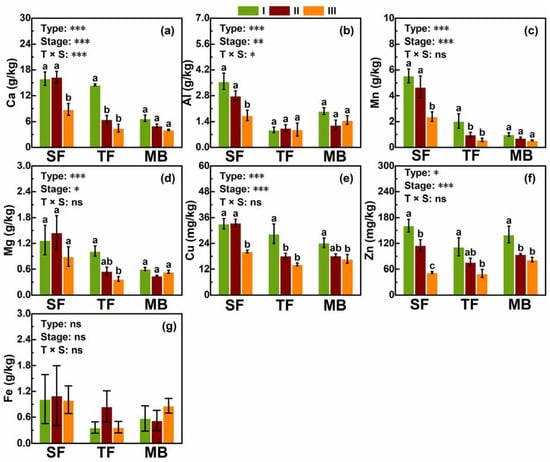
Figure 2.
Concentrations of meso- and micro-elements at the three stages of litter decomposition (I, II, and III) in the three forest types. (a–g) a–g, in turn, is the concentration of Ca, Al, Mn, Mg, Cu, Zn, and Fe in the litter. The codes for forest types are described as in Figure 1. Mean, SE (n = 9), and the significance levels (*** p < 0.001, ** p < 0.01, * p < 0.05, and ns p > 0.05) of two-way ANOVAs are also given. Different letters indicate significant differences between the four stages within a forest type.
3.3. Bacteria
The total number of bacterial OTUs was 6489 in SF, 6652 in MB, and 6727 in TF. Averaging across the four stages, the number of OTUs, Chao1 and Shannon indices of bacteria were not affected by forest types (Figure 3a–c; all p > 0.05). Decomposition stages had significant effects on bacteria, and bacteria were, in general, more abundant and diverse in Stages II and III than in Stages I and IV (Figure 3a–c; all p < 0.001).
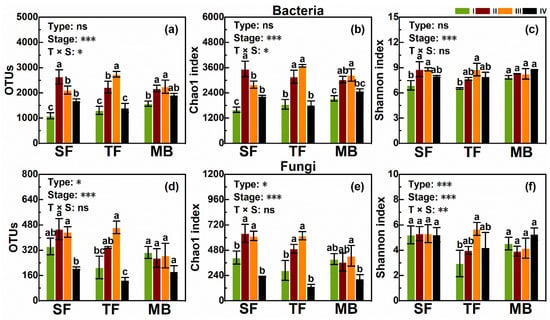
Figure 3.
Community diversity of bacteria (a–c) and fungi (e,f) at the four stages of litter decomposition (I, II, III and IV) in the three forest types. The codes for forest types are described as in Figure 1. Mean, SE (n = 9) and the significance levels (*** p < 0.001, ** p < 0.01, * p < 0.05, and ns p > 0.05) of two-way ANOVAs are also given. Different letters indicate significant differences between the four stages within a forest type.
In the bacterial communities, for the 20 genera with the highest abundance, there was a shift in the relative abundance of, e.g., Burkholderia, Sphingomonas, Enterobacteriaceae, Luteibacter, and Acidisphaera from SF to TF (Figure4). At Stage I, from SF to TF, the abundance of Burkholderia, Sphingomonas, and Enterobacteriaceae decreased by 8.92, 3.66, and 4.22%, respectively, but that of Luteibacter and Acidisphaera increased (Figure 4a). At Stage II, the relative abundance of Burkholderia increased by 10.41% from SF to TF, but that of Enterobacteriaceae decreased by 7.89% (Figure 4b). At Stage III, the relative abundance of WPS-2_genera decreased by 10.4% from SF to TF (Figure 4c). At Stage IV, no change was found between forest types in the first 20 genera (Figure 4c). The remaining taxa (labeled as “others”) increased from SF to TF at all four stages (Figure 4).
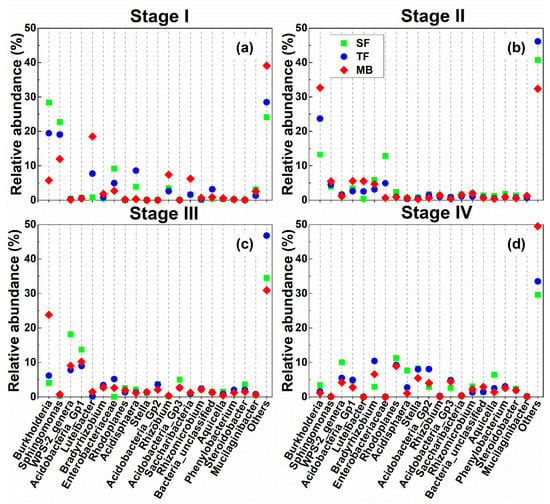
Figure 4.
Relative abundances of bacterial OTU clusters at the genus level from the four litter decomposition stages (I, II, III, and IV) in the three forest types. (a–d) a–d, in turn, is the litter decomposition stages I, II, III, IV. The codes for forest types are described as in Figure 1. The data shown are the 20 taxa with the highest abundances, as well as the summed abundance of the remaining taxa (labeled as “others”).
The first two axes of PCoA explained 72.36% and 8.96% of the variance, respectively. The bacterial communities of the four decomposition stages were well separated along the first axis of PCoA, but those of the three forests types (SF, TF, and MB) could not be distinguished in the ordination space (Figure S1A).
3.4. Fungi
The total number of fungal OTUs was 1853 in TF, 1650 in SF, and 1495 in MB. The forest type significantly affectedthe number of OTUs, Chao1 and Shannon indices of fungi (Figure 3d–f; all p< 0.05). The number of OTUs, Chao1 index, and Shannon index were higher in SF than in MB and TF, but did not differ between MB and SF (Figure 3d–e). Decomposition stages highly and significantly affected the abundance and diversity of fungi (all p< 0.001), and fungi were, in general, more abundant and diverse in Stages II and III as compared to Stages I and IV (Figure 3e–f).
The relative abundance of the first 20 genera varied with both forest types and decomposition stages (Figure 5). At Stage I, from SF to TF, the relative abundance of an unclassified genus of Ascomycota, Sordariomycetes, and Pleosporales decreased by 11.95%, 7.27%, and 7.21%, respectively, but that of Dothideomycetes, Capnodiales, and Amphisphaeriaceae increased by 11.99%, 29.6%, and 20.09%, respectively (Figure 5a). At Stage II, from SF to TF, the relative abundance of the unclassified genus of Ascomycota and Basidiomycota decreased by 24.51% and 12.74%, respectively, but that of Trechispora, Dothideomycetes, Hypocrea, and Geastrum increased by 15.08%, 11.42%, 13.05%, and 8.26%, respectively (Figure 5b). At Stage III, the relative abundance of Russula decreased by 16.03%, but that of Mycena increased by 14.77% (Figure 5c). At Stage IV, the relative abundance of Trechispora decreased by 17.01%, but the unclassified genus of Ascomycota and Tomentella increased by 8.09% and 26.47%, respectively (Figure 5d). The remaining unclassified fungal taxa (labeled as “others”) decreased from SF to TF at all four stages (Figure 5).
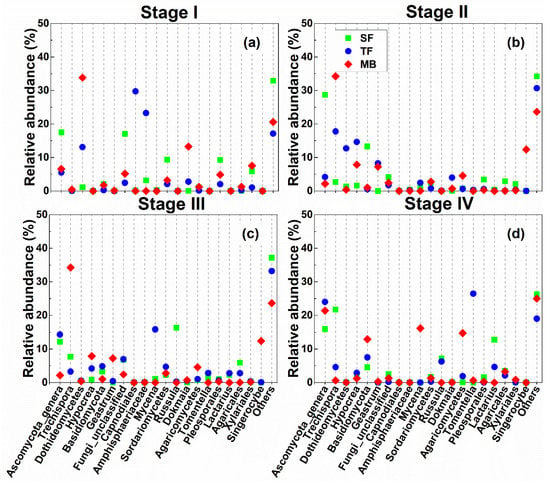
Figure 5.
Relative abundances of fungal OTU clusters at the genus level from the four litter decomposition stages (I, II, III, and IV) in the three forest types. (a–d) a–d, in turn, is the litter decomposition stages I, II, III, IV. The codes for forest types are described as in Figure 1. The data shown are the 20 taxa with the highest abundances, as well as the summed abundance of the remaining taxa (labeled as “others”).
The first two axes of PCoA explained 39.9% and 22.7% of the variance, respectively. The fungal communities of the four decomposition stages were well separated along the first axis of PCoA, but those of the three forest types (SF, TF, and MB) could not be distinguished in the ordination space (Figure S1B).
4. Discussion
4.1. Effects of Moso Bamboo Invasions on Nutrients in Litter and Soil
Plant invasions may affect ecosystems through multiple processes, including the alteration ofenvironmental factors (light and soil nutrients), soil physio-chemical properties, and soil microbial abundance, diversity, and composition [57,58]. Previous studies have shown that bamboo invasions can change C, N, and P concentrations in the soil layer [35,36,37,38,59]. In this study, C, N, P, and lignin concentrations decreased after the bamboo invasion, which is consistent with previous findings [35,36,37,38,59]. In addition, some other studies have shown that the concentrations of soil-exchangeable K, Ca, and Mg changed after the invasion of bamboos, which were close to those in the pure forest of bamboos [60]. In this study, Mg, Al, Ca, Mn, Cu, and Zn concentrations were lower in TF than in SF. The mechanisms of litter decomposition and soil nutrient dynamics are still not clear, but recent studies have shown that it may be closely related to the composition of bacterial and fungal communities [61].
4.2. Effects of Bamboo Invasion on Bacteria and Fungi in Litter and Soil
A previous study showed that the invasion of Moso bamboos increased the diversity of soil bacterial communities [41]. Our results showed that the invasion of Moso bamboo increased microbial diversity in litter and soil, but some bacteria or fungi exhibited different responses to the bamboo invasion. The proportion of bacteria (Burkholderia, Luteibacter, and Acidisphaera) and fungi (Trechispora, Capnodales, Amphisphaeriaceae, Dothideomycetes, and Hypocrea) in litter in TF increased after the bamboo invasion. The increase of these microorganisms may promote litter decomposition [62,63,64]. Conversely, the bamboo invasion led to a decrease in the proportion of some bacteria, such as Sphingomonas and Enterobacteriaceae, which may be due to the inhibition of the growth of some microorganisms in bamboo forests. Such a decrease in microbial diversity also occurred after invasions by plants other than Moso bamboos [55,65].
Given that there were different microbial communities in different litter and soil layers, more definitive results were obtained by stratifying litter into different stages under the three forest types. Litter at Stage I is easily exposed to tissue leaching, initial colonization, and decay; litter at stage II is composed of rich “soil fungi”; and litter at Stage III possesses low mass-loss rates [48]. Litter in the soil layer (at Stage IV) has the largest microbial abundance, which directly affects the composition of microbial communities [56,66]. In this study, microbes were more abundant in Stages II and III than in Stages I and IV. An increase in fungal abundance and diversity mainly occurred during Stage III. This indicates that the bamboo invasion has different effects at different litter decomposition stages.
Compared with bacteria, fungi showed different responses to bamboo invasions because of their low diversity and abundance. It has been proposed that fungal communities in litter are mainly influenced by dominant plant species, while fungal communities in soil are influenced by both dominant plant species and soil physio-chemical properties [67]. In this study, fungi were more susceptible to the bamboo invasion than bacteria, which may be caused by different physiological metabolisms of bacteria and fungi themselves. It is also found that fungal communities are more vulnerable to changes in dominant vegetation than bacterial communities [42]. These findings may explain why fungal communities were more affected by the Moso bamboo invasion in our study.
4.3. Litter Microbial Communities in Relation to Litter Characteristics
Litter nutrients have an important effect on the decomposition of litter and the growth of microorganisms, and the invasive species can influence the growth of the microorganisms and the decomposition of litter by changing the nutrient composition of mixed litter [67,68,69]. Although many studies have reported that changes in tree species affect the composition of microbial communities, it is not clear how microbial communities respond to tree species changes [59,70,71]. Litter with a high concentration of N and a low concentration of lignin is generally considered to be susceptible to microbial decomposition. The heterogeneity and interaction of litter can promote the mobility of microbes and improve the utilization of C and N by microbes [72]. For example, compared to coniferous forests, broadleaved forests produce a smaller amount of litter with high N, thereby releasing a larger amount of cations such as Ca2+ and Mg2+, promoting the soil to become acidic, and affecting the growth and activity of the microbes [70]. Lignin is mainly degraded by a specific fungal community [72]. In our study, bacterial diversity in litter was significantly negatively correlated with the concentration of cellulose, lignin, N, C, and P in litter, as well as with the concentration of metal elements such as Zn, Cu, Ca, K, and Al in litter (Figures S2 and S3). The fungal diversity in litter was significantly negatively correlated with the cellulose concentration in litter (Figure S3). We show that changes in nutrients in litter can affect the structure of microbial communities in the litter and soil [21,63].
5. Conclusions
We found that native Moso bamboo invasions could decrease soil nutrient concentrations and modify fungal abundance, diversity, and composition. Also, fungi were more sensitive to native Moso bamboo invasions than bacteria. These findings suggest that native Moso bamboo invasions into subtropical forests may lead to a complex biogeochemical process in the litter–soil system, which can further threaten the local forest ecosystems by affecting microbial communities and, thus, litter decomposition and nutrient cycling. Further studies are needed to better understand how these changes will affect the litter decomposition processes and, thus, soil quality and plant–soil feedback processes.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/11/3/314/s1, Figure S1: Principal coordinate analysis of (A) bacterial and (B) fungal communities at the four litter decomposition stages, Figure S2: Correlation matrix between bacterial diversity and content of C, N, P, cellulose and lignin in litter layer, Figure S3: Correlation matrix between bacterial diversity and metal elements in litter layer, Figure S4: Fungal diversity and content of C, N, P, cellulose and lignin in litter layer, Figure S5: Correlation matrix between fungal diversity and content of C, N, P, cellulose and lignin in litter layer.
Author Contributions
Conceptualization, X.-K.T. and X.-G.G.; methodology, X.-K.T. and M.-Y.W.; software, X.-K.T., M.-Y.W., and P.M.; formal analysis, X.-K.T. and X.-G.G.; investigation, X.-K.T., X.-G.G. and B.-Z.Z.; resources, P.M.; data curation, P.M., J.-S.Z. and B.-Z.Z.; writing—original draft preparation, X.-K.T. and M.-Y.W.; writing—review and editing, F.-H.Y., J.-S.Z. and M.-H.L.; supervision, F.-H.Y., X.-G.G. and J.-S.Z.; project administration, B.-Z.Z. and P.M.; funding acquisition, X.-K.T., X.-G.G. and F.-H.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Fundamental Research Funds of CAF (CAFYBB2018SY015 and CAFYBB2018GB001) and NSFC (31870610).
Acknowledgments
We thank anonymous reviewers for helpful comments that improved the original manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Marbuah, G.; Gren, I.-M.; McKie, B. Economics of Harmful Invasive Species: A Review. Diversity 2014, 6, 500–523. [Google Scholar] [CrossRef]
- Engelkes, T.; Meisner, A.; Morrien, E.; Kostenko, O.; Van Der Putten, W.H.; Macel, M. Herbivory and dominance shifts among exotic and congeneric native plant species during plant community establishment. Oecologia 2016, 180, 507–517. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Müller-Schärer, H.; Van Kleunen, M.; Cai, A.-M.; Zhang, P.; Yan, R.; Dong, B.-C.; Yu, F.-H. Invasive alien plants benefit more from clonal integration in heterogeneous environments than natives. New Phytol. 2017, 216, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-J.; Chen, D.; Yan, R.; Yu, F.-H.; Van Kleunen, M. Invasive alien clonal plants are competitively superior over co-occurring native clonal plants. Perspect. Plant Ecol. Evol. Syst. 2019, 40, 125484. [Google Scholar] [CrossRef]
- Zenni, R.D.; Ziller, S.R.; Pauchard, A.; Rodriguez-Cabal, M.; Nuñez, M.A. Invasion science in the developing world: a response to ricciardi et al. Trends Ecol. Evol. 2017, 32, 807–808. [Google Scholar] [CrossRef] [PubMed]
- Prevéy, J.S.; Germino, M.J.; Huntly, N.; Inouye, R. Exotic plants increase and native plants decrease with loss of foundation species in sagebrush steppe. Plant Ecology 2009, 207, 39–51. [Google Scholar] [CrossRef]
- Davies, K.W. Plant community diversity and native plant abundance decline with increasing abundance of an exotic annual grass. Oecologia 2011, 167, 481–491. [Google Scholar] [CrossRef]
- A Driscoll, D. Disturbance maintains native and exotic plant species richness in invaded grassy woodlands. J. Veg. Sci. 2017, 28, 573–584. [Google Scholar] [CrossRef]
- Cleland, E.E.; Larios, L.; Suding, K. Strengthening invasion filters to reassemble native plant communities: soil resources and phenological overlap. Restor. Ecol. 2012, 21, 390–398. [Google Scholar] [CrossRef]
- Jackson, R.B.; Banner, J.L.; Jobbágy, E.G.; Pockman, W.; Wall, D.H. Ecosystem carbon loss with woody plant invasion of grasslands. Nature 2002, 418, 623–626. [Google Scholar] [CrossRef]
- Sardans, J.; Bartrons, M.; Margalef, O.; Gargallo-Garriga, A.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Sigurdsson, B.D.; Chen, H.Y.H.; Penuelas, J. Plant invasion is associated with higher plant-soil nutrient concentrations in nutrient poor-environments. Glob. Chang. Boil. 2016, 23, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Sardans, J.; Wang, C.; Zeng, C.; Tong, C.; Chen, G.; Huang, J.; Pan, H.; Peguero, G.; Vallicrosa, H.; et al. The response of stocks of C, N, and P to plant invasion in the coastal wetlands of China. Glob. Chang. Boil. 2018, 25, 733–743. [Google Scholar] [CrossRef]
- Aragón, R.; Montti, L.; Ayup, M.M.; Fernandez, R.D. Exotic species as modifiers of ecosystem processes: Litter decomposition in native and invaded secondary forests of NW Argentina. Acta Oecologica 2014, 54, 21–28. [Google Scholar] [CrossRef]
- Braun, K.; Collantes, M.B.; Yahdjian, L.; Escartin, C.; Anchorena, J.A. Increased litter decomposition rates of exotic invasive species Hieracium pilosella (Asteraceae) in Southern Patagonia, Argentina. Plant Ecology 2019, 220, 393–403. [Google Scholar] [CrossRef]
- Castro-Diez, P.; Álvaro, A.; Romero-Blanco, A. Effects of litter mixing on litter decomposition and soil properties along simulated invasion gradients of non-native trees. Plant Soil 2019, 442, 79–96. [Google Scholar] [CrossRef]
- Inderjit; Van Der Putten, W.H. Impacts of soil microbial communities on exotic plant invasions. Trends Ecol. Evol. 2010, 25, 512–519. [Google Scholar] [CrossRef]
- Hynson, N.A.; Merckx, V.S.; Perry, B.A.; Treseder, K. Identities and distributions of the co-invading ectomycorrhizal fungal symbionts of exotic pines in the Hawaiian Islands. Boil. Invasions 2013, 15, 2373–2385. [Google Scholar] [CrossRef]
- McLeod, M.L.; Cleveland, C.C.; Lekberg, Y.; Maron, J.L.; Philippot, L.; Bru, D.; Callaway, R.M. Exotic invasive plants increase productivity, abundance of ammonia-oxidizing bacteria and nitrogen availability in intermountain grasslands. J. Ecol. 2016, 104, 994–1002. [Google Scholar] [CrossRef]
- Perkins, L.; Johnson, D.W.; Nowak, R.S. Plant-induced changes in soil nutrient dynamics by native and invasive grass species. Plant Soil 2011, 345, 365–374. [Google Scholar] [CrossRef]
- Wickramathilake, B.A.K.; Weerasinghe, T.K.; Ranwala, S.M.W. Impacts of woody invader Dillenia suffruticosa (Griff.) Martelli on physio-chemical properties of soil and, below and above ground flora. J. Trop. Forest. Environ. 2013, 3, 66–75. [Google Scholar] [CrossRef]
- Feng, J.; Zhou, J.; Wang, L.; Cui, X.; Ning, C.; Wu, H.; Zhu, X.; Lin, G. Effects of short-term invasion of Spartina alterniflora and the subsequent restoration of native mangroves on the soil organic carbon, nitrogen and phosphorus stock. Chemosphere 2017, 184, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Tong, C.; Zhang, L.; Wang, W.; Gauci, V.; Marrs, R.; Liu, B.; Jia, R.; Zeng, C. Contrasting nutrient stocks and litter decomposition in stands of native and invasive species in a sub-tropical estuarine marsh. Environ. Res. 2011, 111, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Mincheva, T.; Barni, E.; Varese, G.C.; Brusa, G.; Cerabolini, B.E.L.; Siniscalco, C. Litter quality, decomposition rates and saprotrophic mycoflora in Fallopia japonica (Houtt.) RonseDecraene and in adjacent native grassland vegetation. Acta Oecologica 2014, 54, 29–35. [Google Scholar] [CrossRef]
- Niu, H.-B.; Liu, W.-X.; Wan, F.-H.; Liu, B. An invasive aster (Ageratina adenophora) invades and dominates forest understories in China: altered soil microbial communities facilitate the invader and inhibit natives. Plant Soil 2007, 294, 73–85. [Google Scholar] [CrossRef]
- Perkins, L.; Nowak, R.S. Native and non-native grasses generate common types of plant-soil feedbacks by altering soil nutrients and microbial communities. Oikos 2012, 122, 199–208. [Google Scholar] [CrossRef]
- Mamet, S.D.; Lamb, E.G.; Piper, C.L.; Winsley, T.; Siciliano, S.D. Archaea and bacteria mediate the effects of native species root loss on fungi during plant invasion. ISME J. 2017, 11, 1261–1275. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Chauhan, B.S.; Farooq, M.; Shabbir, A.; Adkins, S.W. What do we really know about alien plant invasion? A review of the invasion mechanism of one of the world’s worst weeds. Planta 2016, 244, 39–57. [Google Scholar] [CrossRef]
- Warren, R.J.; King, J.R.; Tarsa, C.; Haas, B.; Henderson, J. A systematic review of context bias in invasion biology. PLoS One 2017, 12, e0182502. [Google Scholar] [CrossRef]
- Nackley, L.; West, A.; Skowno, A.L.; Bond, W.J. The nebulous ecology of native invasions. Trends Ecol. Evol. 2017, 32, 814–824. [Google Scholar] [CrossRef]
- Ying, W.; Jin, J.; Jiang, H.; Zhang, X.; Lu, X.; Chen, X.; Zhang, J. Satellite-based detection of bamboo expansion over the past 30 years in Mount Tianmushan, China. Int. J. Remote. Sens. 2016, 37, 2908–2922. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Yao, X.M.; Tian, X.M. The regeress analysis between fertilization and the output, the economic benefit in Phyllostachys pubescens for culm-producing. Chin. Agr. Sci. Bull. 2009, 25, 166–169. [Google Scholar]
- Li, X.X.; Liu, P.; Rong, J.D.; Li, S.K.; Chen, L.Y.; He, T.Y.; Zheng, Y.S. Research state and development trend of moso bamboo forest management. World Bamboo Ratt. 2018, 16, 58–61. [Google Scholar]
- Xu, Q.-F.; Liang, C.-F.; Chen, J.-H.; Li, Y.-C.; Qin, H.; Fuhrmann, J. Rapid bamboo invasion (expansion) and its effects on biodiversity and soil processes +. Glob. Ecol. Conserv. 2020, 21, e00787. [Google Scholar] [CrossRef]
- Bai, S.B.; Zhou, G.M.; Wang, Y.X.; Liang, Q.Q.; Chen, J.; Cheng, Y.Y.; Shen, R. Plant species diversity and dynamics in forests invaded by moso bamboo (Phyllostachys edulis) in tianmu mountain nature reserve. Biodiversity Sci. 2013, 21, 288–295. [Google Scholar]
- Fukushima, K.; Usui, N.; Ogawa, R.; Tokuchi, N. Impacts of moso bamboo (Phyllostachys pubescens) invasion on dry matter and carbon and nitrogen stocks in a broad-leaved secondary forest located in K yoto, western Japan. Plant. Spec. Biol. 2015, 30, 81–95. [Google Scholar] [CrossRef]
- Wang, H.-C.; Tian, G.; Chiu, C.-Y. Invasion of moso bamboo into a Japanese cedar plantation affects the chemical composition and humification of soil organic matter. Sci. Rep. 2016, 6, 32211. [Google Scholar] [CrossRef] [PubMed]
- Shiau, Y.-J.; Chiu, C.-Y. Changes in Soil Biochemical Properties in a Cedar Plantation Invaded by Moso Bamboo. Forests 2017, 8, 222. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, L.; Deng, B.; Liu, Y.; Kong, F.; Huang, G.; Zou, Q.; Liu, Q.; Guo, X.; Fu, Y.; et al. Effects of moso bamboo (Phyllostachys edulis) invasions on soil nitrogen cycles depend on invasion stage and warming. Environ. Sci. Pollut. Res. 2017, 24, 24989–24999. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Stanek, M.; Nobis, M.; Zubek, S. Species-specific effects of plant invasions on activity, biomass, and composition of soil microbial communities. Boil. Fertil. Soils 2016, 52, 841–852. [Google Scholar] [CrossRef]
- Dawson, W.; Schrama, M. Identifying the role of soil microbes in plant invasions. J. Ecol. 2016, 104, 1211–1218. [Google Scholar] [CrossRef]
- Xu, Q.; Jiang, P.-K.; Wu, J.-S.; Zhou, G.-M.; Shen, R.-F.; Fuhrmann, J.J. Bamboo invasion of native broadleaf forest modified soil microbial communities and diversity. Boil. Invasions 2014, 17, 433–444. [Google Scholar] [CrossRef]
- Bani, A.; Pioli, S.; Ventura, M.; Panzacchi, P.; Borruso, L.; Tognetti, R.; Tonon, G.; Brusetti, L. The role of microbial community in the decomposition of leaf litter and deadwood. Appl. Soil Ecol. 2018, 126, 75–84. [Google Scholar] [CrossRef]
- Purahong, W.; Wubet, T.; Lentendu, G.; Schloter, M.; Pecyna, M.J.; Kapturska, D.; Hofrichter, M.; Krüger, D.; Buscot, F. Life in leaf litter: novel insights into community dynamics of bacteria and fungi during litter decomposition. Mol. Ecol. 2016, 25, 4059–4074. [Google Scholar] [CrossRef] [PubMed]
- Hobara, S.; Osono, T.; Hirose, D.; Noro, K.; Hirota, M.; Benner, R. The roles of microorganisms in litter decomposition and soil formation. Biogeochem. 2013, 118, 471–486. [Google Scholar] [CrossRef]
- Keiser, A.D.; Knoepp, J.D.; Bradford, M.A. Microbial communities may modify how litter quality affects potential decomposition rates as tree species migrate. Plant Soil 2013, 372, 167–176. [Google Scholar] [CrossRef]
- Elgersma, K.J.; Ehrenfeld, J.G.; Yu, S.; Vor, T. Legacy effects overwhelm the short-term effects of exotic plant invasion and restoration on soil microbial community structure, enzyme activities, and nitrogen cycling. Oecologia 2011, 167, 733–745. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.L.; Zhou, B.; Ge, X.G.; Wang, X.-M.; Li, Q. Leaf-litter decomposition dynamic, carbon loss and nutrient return for Moso Bamboo forest with different litter mass accumulation. Adv. Mater. Res. 2013, 726, 4222–4225. [Google Scholar] [CrossRef]
- Kanerva, S.M.; Smolander, A. Microbial activities in forest floor layers under silver birch, Norway spruce and Scots pine. Soil Boil. Biochem. 2007, 39, 1459–1467. [Google Scholar] [CrossRef]
- Bao, S.D. Agro-Chemical Analysis of Soil; Agricultural Publish House of China: Beijing, China, 2000; pp. 146–336. [Google Scholar]
- Rowland, A.P.; Roberts, J.D. Lignin and cellulose fractionation in decomposition studies using acid?detergentfibre methods. Commun. Soil Sci. Plant Anal. 1994, 25, 269–277. [Google Scholar] [CrossRef]
- Blaxter, M.; Mann, J.; Chapman, T.; Thomas, F.; Whitton, C.; Floyd, R.; Abebe, E. Defining operational taxonomic units using DNA barcode data. Philos. Trans. R. Soc. B Boil. Sci. 2005, 360, 1935–1943. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Drenovsky, R.E.; Grewell, B.J.; D’Antonio, C.M.; Funk, J.L.; James, J.J.; Molinari, N.; Parker, I.M.; Richards, C. A functional trait perspective on plant invasion. Ann. Bot. 2012, 110, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Tlaskal, V.; Baldrian, P.; Voriskova, J.; Boer, W. Bacterial succession on decomposing leaf litter exhibits a specific occurrence pattern of cellulolytic taxa and potential decomposers of fungal mycelia. FEMS Microbiol. Ecol. 2016, 92, 177. [Google Scholar] [CrossRef] [PubMed]
- Grayston, S.J.; Prescott, C.E. Microbial communities in forest floors under four tree species in coastal British Columbia. Soil Boil. Biochem. 2005, 37, 1157–1167. [Google Scholar] [CrossRef]
- Ushio, M.; Kitayama, K.; Balser, T.C. Tree species-mediated spatial patchiness of the composition of microbial community and physicochemical properties in the topsoils of a tropical montane forest. Soil Boil. Biochem. 2010, 42, 1588–1595. [Google Scholar] [CrossRef]
- Sodhi, D.; Livingstone, S.W.; Carboni, M.; Cadotte, M.W. Plant invasion alters trait composition and diversity across habitats. Ecol. Evol. 2019, 9, 6199–6210. [Google Scholar] [CrossRef]
- Wu, C.; Mo, Q.; Wang, H.; Zhang, Z.; Huang, G.; Ye, Q.; Zou, Q.; Kong, F.; Liu, Y.; Wang, G.G. Moso bamboo (Phyllostachys edulis (Carriere) J. Houzeau) invasion affects soil phosphorus dynamics in adjacent coniferous forests in subtropical China. Ann. For. Sci. 2018, 75, 24. [Google Scholar] [CrossRef]
- Umemura, M.; Takenaka, C. Changes in chemical characteristics of surface soils in hinoki cypress (Chamaecyparis obtusa) forests induced by the invasion of exotic Moso bamboo (Phyllostachys pubescens) in central Japan. Plant Species Boil. 2014, 30, 72–79. [Google Scholar] [CrossRef]
- Prescott, C.E.; Grayston, S.J. Tree species influence on microbial communities in litter and soil: Current knowledge and research needs. For. Ecol. Manag. 2013, 309, 19–27. [Google Scholar] [CrossRef]
- Ke, P.; Miki, T.; Ding, T.-S. The soil microbial community predicts the importance of plant traits in plant-soil feedback. New Phytol. 2014, 206, 329–341. [Google Scholar] [CrossRef]
- Day, N.; Dunfield, K.E.; Antunes, P.M. Fungi from a non-native invasive plant increase its growth but have different growth effects on native plants. Boil. Invasions 2015, 18, 231–243. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, J.; Liu, J.; Jiang, K.; Xiao, H.; Du, D. Responses of the soil fungal communities to the co-invasion of two invasive species with different cover classes. Plant Boil. 2017, 20, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Hodge, A.; Fitter, A. Microbial mediation of plant competition and community structure. Funct. Ecol. 2012, 27, 865–875. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, J.-T.; Ma, K. Soil biota reduce allelopathic effects of the invasive Eupatorium adenophorum. PLoS ONE 2011, 6, e25393. [Google Scholar] [CrossRef]
- Urbanová, M.; Šnajdr, J.; Baldrian, P. Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees. Soil Boil. Biochem. 2015, 84, 53–64. [Google Scholar] [CrossRef]
- Sun, H.; Wang, Q.; Liu, N.; Li, L.; Zhang, C.-G.; Liu, Z.B.; Zhang, Y.-Y. Effects of different leaf litters on the physicochemical properties and bacterial communities in Panax ginseng -growing soil. Appl. Soil Ecol. 2017, 111, 17–24. [Google Scholar] [CrossRef]
- He, Z.; Yu, Z.; Huang, Z.; Davis, M.; Yang, Y. Litter decomposition, residue chemistry and microbial community structure under two subtropical forest plantations: A reciprocal litter transplant study. Appl. Soil Ecol. 2016, 101, 84–92. [Google Scholar] [CrossRef]
- Dukunde, A.; Schneider, D.; Schmidt, M.; Veldkamp, E.; Daniel, R. Tree species shape soil bacterial community structure and function in temperate deciduous forests. Front. Microbiol. 2019, 10, 1519. [Google Scholar] [CrossRef]
- Bissett, A.; Brown, M.V.; Siciliano, S.D.; Thrall, P.H. Microbial community responses to anthropogenically induced environmental change: towards a systems approach. Ecol. Lett. 2013, 16, 128–139. [Google Scholar] [CrossRef]
- Boberg, J.; Finlay, R.; Stenlid, J.; Ekblad, A.; Lindahl, B.D. Nitrogen and carbon reallocation in fungal Mycelia during decomposition of boreal forest litter. PLoS ONE 2014, 9, e92897. [Google Scholar] [CrossRef]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest soil bacteria: diversity, involvement in ecosystem processes, and response to global change. Microbiol. Mol. Boil. Rev. 2017, 81, 00063-16. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).